

Abstract
Aim: We investigated differences of metastatic spread of normal proteinase-activated receptor-2 (Par2+/+) melanoma B16 in Par2−/− (knock-out) animals compared to C57Bl6 mice. Materials and Methods: Nine knock-out mice B6.Cg-F2rl1tm1Mslb/J (Par2−/−) and nine C57Bl6/J controls were subcutaneously inoculated with B16 melanoma tissue cells. Twelve days after inoculation, all primary tumors were removed. Survival and metastatic spread was followed for up to 100 days after primary tumor extirpation. Results: Excised primary tumors were on average larger in Par2−/− mice (360 mm3 vs. 221 mm3 in C57Bl6/J). Distant spontaneous metastases developed in only 3 of 9 of Par2−/− mice in comparison to 6 of 9 controls. The average survival time was 84 days in Par2−/− animals compared to 63 days in C57Bl6/J mice. Conclusion: Host Par2 melanoma model contributes to the limitation of local cancer progression in one area, while on the other hand is important for enhancing distant metastatic spread.
- PAR2
- melanoma
- metastasis
- murine model
Proteinase-activated receptor-2 (PAR2) belongs to a family of G-protein-coupled receptors activated by tethered ligand sequences within the amino terminal part of the molecule that is made accessible by site-specific proteolysis by trypsin and trypsin-like proteinases. PAR2 is widely expressed on many different cells within the body and is thought to contribute to a broad range of normal and disease-related processes including embryogenesis, pain and nociception, acute and chronic inflammation, arthritis and cancer (1). Its role has been investigated in different types of cancer, including melanoma (2), where it was found to contribute to tumor cell motility and metastatic progression (3). However, unlike PAR1 (activated by thrombin and thrombin-like proteinases), there is no difference in expression of PAR2 between melanocytic nevi and malignant melanoma (4). Recently, it was shown that PAR2 regulates invasive behavior associated with human melanoma cells via activation of protein kinase D1 (5). Even so, the contribution of PAR2 activation to malignant development and metastasis formation by stimulation of trypsin and trypsin-like proteinases is generally accepted (6). Additionally there is the possibility of non-specific enhancement of different intracellular processes secondary to PAR2 activation in primarily transformed malignant cells that must be considered (7). However, only a few articles have discussed the role of the host (whole body environment) PAR2 in tumor growth and metastasis. Camerer et al. showed that in Par2-knockout mice in the metastatic model of B16 melanoma injected into the tail vein, no difference in metastatic spread was observed (8). On the other hand, Schaffner et al. reported the importance of PAR2 signaling in promoting angiogenesis in breast cancer (9). To evaluate the role of host PAR2 in a spontaneous metastatic model, which encompasses more physiological conditions, we utilized an animal experiment to further define the receptor role during local progression of malignant disease and its metastatic spread. Our previous pilot study showed the metastatic potential of normal (Par2+/+) B16 melanoma (10) in Par2-deficient mice, however, due to the limited number of animals, statistically evaluable results related to metastatic spread and survival could not be obtained at all. Moreover, we were also unable to precisely evaluate local growth of tumors. Therefore the experiment was repeated with larger cohorts of transgenic animals and wild-type controls.
Materials and Methods
Animals. The study was performed in accordance to the Declaration of Helsinki and guidelines on animal experimentation of the First Medical School, Charles University and all procedures were approved by the animal Experimentation Review Ethics Committee under reference number 255/08 l.LFUK.
In the presented study, nine male inbred Par2−/− B6.Cg-F2rl1tm1Mslb/J mice (Jackson Laboratories, Bar Harbor, ME, USA) and nine wild-type (WT) inbred C57Bl6/J (AnLab s.r.o.-Charles River, Prague, Czech Republic) having equal genetic backgrounds were subcutaneously injected with 4.0×106 B16 melanoma cells (B16F1, European Collection of Cell Cultures, England) in 0.2 ml into the right flank. The mice were kept in a barrier facility for animals, provided with radiation-sterilized bedding (SAWI Research Bedding, JELU-WERK, Germany), fed with radiation-sterilized ST-1 diet (Bergmann, Prague, Czech Republic); the mice received autoclaved water ad libitum.
RT PCR. Total RNA was used for evaluation of Par2 status in both groups of animals (TRIzol®Reagent; Invitrogen, Carlsbad, California, USA). Primers for Par2 were 5’-TGGCCATTGGAGTCTTCCTGTT-3’, 5’-TAGCCCTCTGCCTTTTCTTCTC-3’, for Par1 were 5’-TCCTTTCTCACACTTCCACC-3’and 5-GTTCAGGGCTAAACTCTACC-3’. DNA polymerase was used for reverse transcription and PCR (SuperScript® III One-Step RT-PCR System with Platinum®Taq; Invitrogen, Carlsbad, California, USA). Amplification cycles consisted of 45 s at 93°C, 45 s at 55°C and 1 min at 72°C for 30 cycles. The PCR reaction was performed on a MJ Research PTC-200 PCR thermo cycler (Bio-Rad Laboratories, Hercule, California, USA). Products were analyzed electrophoretically on a 1% agarose gel with ethidium bromide.
Tumor growth. Twelve days after subcutaneous transplantation of B16 melanoma cells, the primary tumors were removed and measured. The volume was calculated from the formula V=½A×B2 where A was the largest and B the smallest dimension of the tumor.
Observation, autopsy and histology. Primary tumors were removed on day 12, survival and metastatic spread was followed for up to 100 days from primary tumor extirpation. Removed tumors were characterized macroscopically and histopathologically to evaluate the level of regressive changes and other differences between tumors from Par2−/− animals and WT mice. The deterioration of animals was followed in both groups every day for a total of 100 days. All surviving animals were sacrificed after this time period.
Dissection and histological verification was carried-out in all animals, those that died of disease and those that were sacrificed after 100 days. The lung, liver and local recurrence of primary tumors and metastases into axillary lymph nodes, as far as they were macroscopically distinct, were subjected to standard histopathological examinations. Briefly, they were fixed in 10% buffered formalin and embedded in paraffin blocks. Serial slices of 5 μm were subsequently prepared and stained with hematoxylin and eosin (HE). The lungs were embedded in toto using the above procedure and transversally cut at the level of the heart ventricles and embedded in paraffin blocks, which were serially sectioned. After H&E staining, metastatic processes were either verified or excluded.

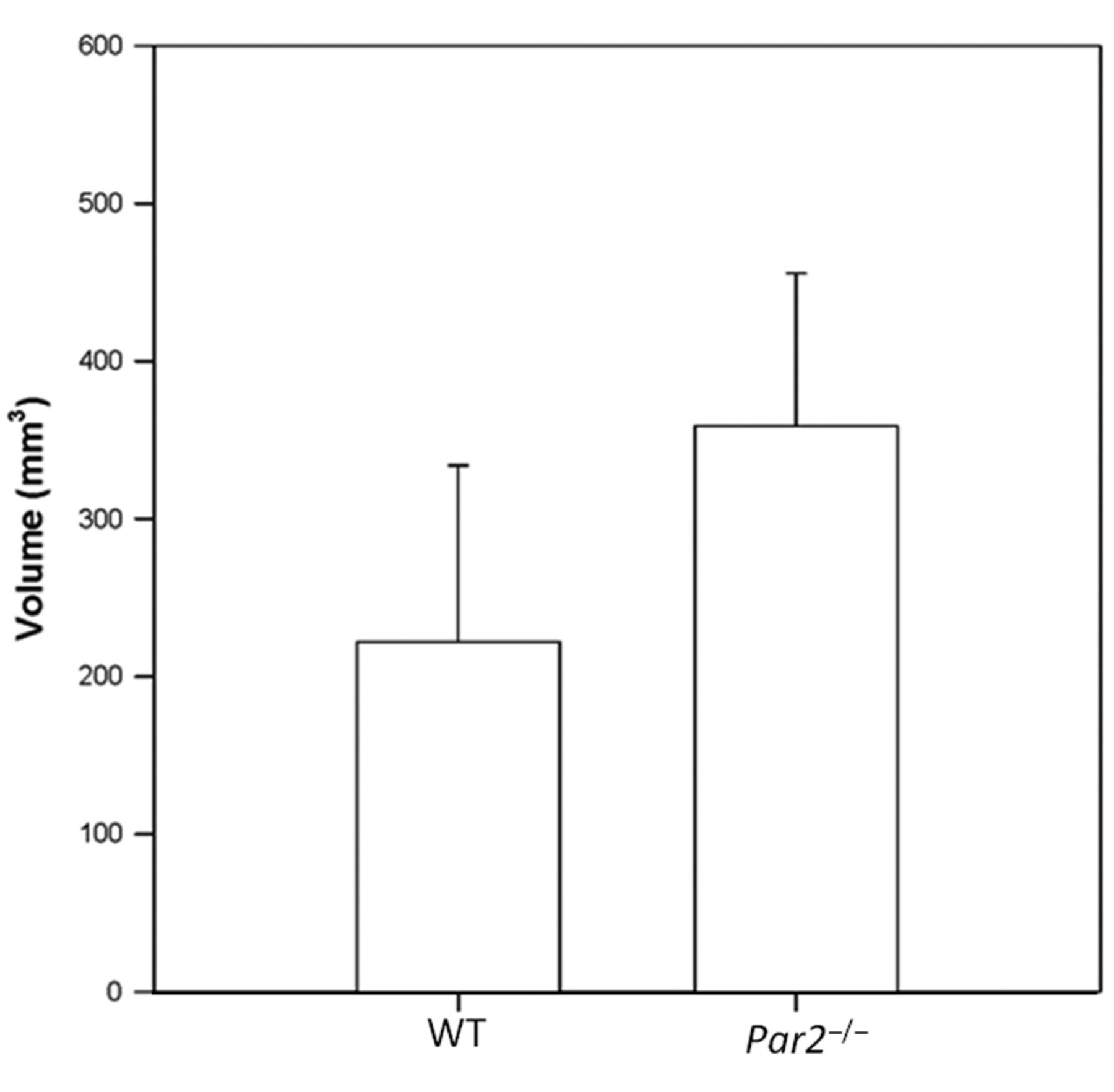
Volume of primary tumors excised from Par2−/− and WT mice (av. 360 mm3, median 325 mm3, SD=+/−96,6 in Par2−/− (n=9) vs. 221 mm3, 205 mm3, SD=+/−111,9, respectively in WT (n=9) mice). The difference between groups is statistically significant (p=0.013).
Immunohistochemistry. Microscopic tissue slices, 5 μm-thick, were de-paraffinized, rehydrated and pretreated in a citrate buffer (pH 7.6) for 3×5 min in a microwave oven. Endogenous peroxidase was blocked with a water solution containing 0.01% sodium azide and 1% hydrogen peroxide. Non-specific positivity was blocked by rabbit serum in tris buffered saline (TBS) (150 μl/10 ml) for 30 min. Primary polyclonal rabbit antibody against S100 protein was used at a dilution of 1:500 (Dako, Glostrup, Denmark) and slices were incubated overnight at 4°C. The detection of immunostaining was performed using the Envision® kit (Dako, Glostrup, Denmark) and diaminobenzidine was used as a chromogen. Slides incubated with secondary antibody only and with nonspecific isotype-matched primary antibodies were used as control for specificity. Mayer's hematoxylin was used as a nuclear counterstain. As a negative control, slides of bronchial endoscopic samples were incubated with nonspecific isotype-matched primary antibodies and with secondary antibody only.
Statistics. Standard two-sample t-test was used for testing tumor volume for both WT and Par2−/− groups. The same test was used for testing tumor weight equity for WT and Par2−/− groups. Survival curves of WT and Par2−/− groups were tested using the Mantel-Cox test of hazard equity. All the statistical calculations were performed using the MATLAB Statistical Toolbox MATLAB 7.8.0 Statistical Toolbox (Mathworks Inc., Natick, Massachusetts, USA) with a critical probability of 0.05.

Histology (A and B) and immunohistochemistry (C and D) of tumors from WT (A and C) and Par2−/− (B and D) mice. A and B: No significant regressive changes in primary tumors in either group were observed. HE, original magnification ×400. C and D: The expression of S100 as a marker for malignant melanoma was mostly the same intensity in both cohorts of investigated tumors. Original magnification ×400.
Results
Using RT PCR, we confirmed that the knock-out animals were lacking Par2 RNA (data not shown). Excised tumors differed in volume, being greater in Par2−/− mice than in WT (av. 360 mm3, median 325 mm3, SD=+/−96,6 in Par2−/− (n=9) vs. 221 mm3 and 205 mm3, SD=+/−111.9, respectively in WT (n=9) mice) (Figure 1). Tumors also differed in average weight (av. 0.288 g, median 0.290 g, SD=+/−0.078 in Par2−/− vs. 0.215 g and 0.217 g, SD=+/−0,088, respectively, in WT). The tumor weight equality hypothesis for WT vs. Par2−/− was not rejected (p=0.0821), however, tumor volume differences between groups were highly statistically significant (p=0.013). Microscopically, we observed regressive changes in primary tumors in both groups without significant differences (Figure 2A and B). The expression of S100 showed mostly the same intensity in both cohorts of investigated tumors (Figure 3C and D).
Oddly, distant spontaneous metastases developed in only 3 out of 9 Par2−/− animals in comparison to 6 out of 9 controls. No recurrence was recorded in either group. In WT animals, metastatic dissemination was recorded in the lungs (5 out of 9), in the axillary lymph nodes of the right forelimb (4 out of 9) and in the brain (2 out of 9). In PAR-2 knock-out, disseminated melanoma was observed in the lungs (3 out of 9), in the axillary lymph node of the right forelimb (3 of 9) and in the right kidney (1 out of 9).
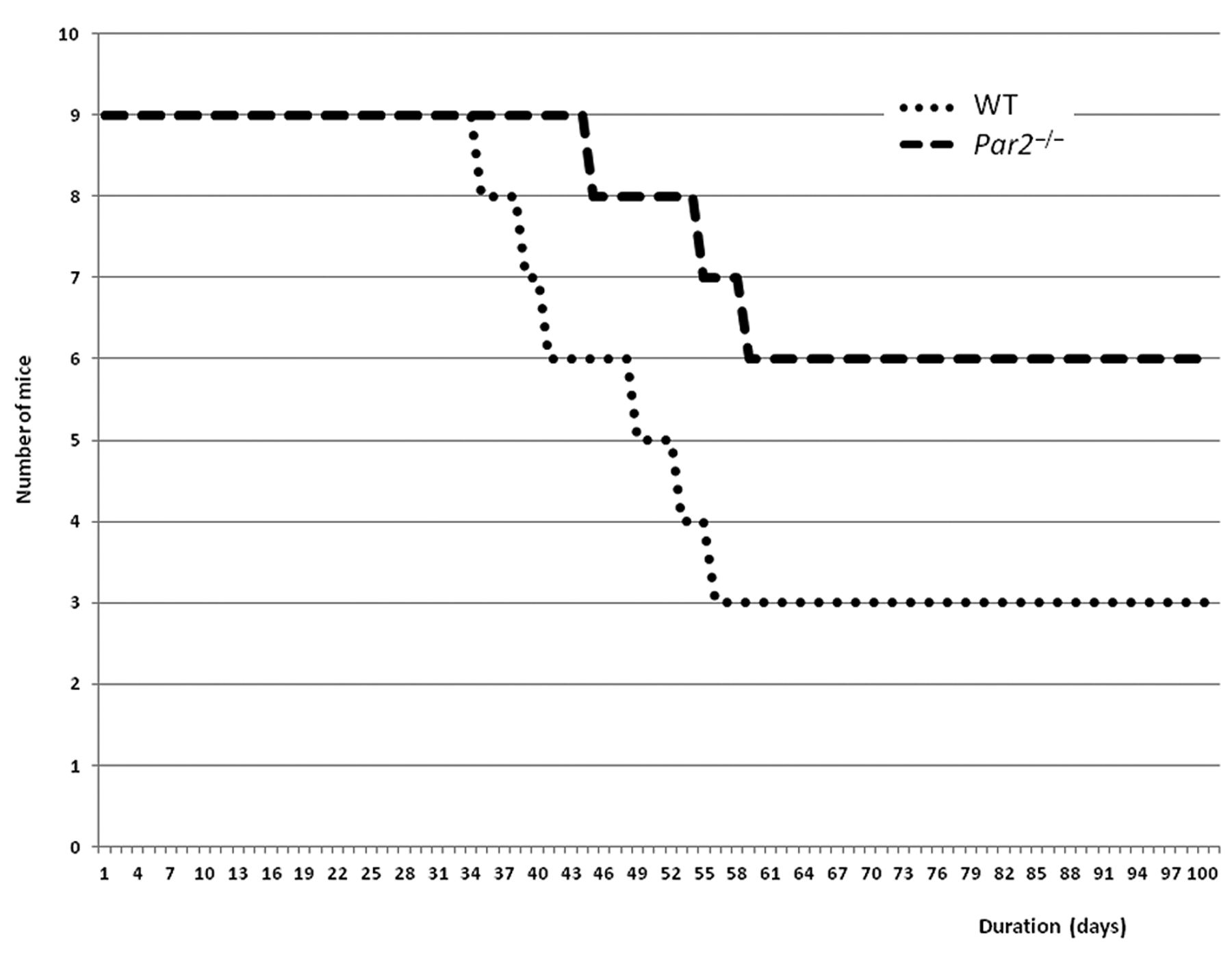
The average survival time was 84 days in Par2−/− animals compared to 63 days in WT mice (Figure 3). This difference was highly statistically significant (p-value for Mantel-Cox test was 0.031).

{kind=link}
{kind=link}
{kind=link}
Survival analysis of Par2−/− (n=9) and WT (n=9) mice. The average survival time was 84 days in the Par2−/− group compared to 63 days in the WT group (p=0.0031, Mantel-Cox test).
Discussion
In line with the published role of PAR2 in dendritic cell maturation (11, 12) and antigen presentation (13), and our previous results with low statistical significance, we observed increased primary tumor volume in Par2 knock-out mice compared to controls (WT). This result was statistically, although not clinically significant (i.e. the average tumor volume of the target was not reduced by 50% compared to controls) and cannot explain the natural behavior of B16 melanoma at the primary site. Thus, the situation is not physiological because implanted tumor cells act as a genetically unique allograft which stimulates, to a greater or lesser degree, the host's cytotoxic immune defense, which probably dominates at the local level. However, repeated results confirm that the absence of Par2 could have a positive influence on local tumor progression, which could have potential therapeutic benefits via Par2 regulation. At the same time, we cannot overlook the fact that prolonged survival and reduction of metastatic dissemination was observed in Par2−/− mice, which are totally opposite results compared to previously published observations by Camerer et al. (8). These disparities need to be explained using a different method of metastasis induction. Our previous pilot results also cannot be taken into account due to their low statistical significance, lack of a comparable control group and different times of primary tumor growth (14 vs. 12 days). Based on our previous unpublished experience, we know that distant metastatic spreading markedly increases with one to two days of prolongation of primary tumor growth. The primary objective of our pilot study was to set up the experimental procedure and to evaluate the capability of metastatic spread in the model of Par2 knock-out mice. Apparently long-lasting, spontaneously spreading tumor cells from primary B16 melanomas utilize the body's intrinsic Par2 to facilitate dissemination. Moreover, links between cancer spread, inflammation, angiogenesis and vascular growth, and coagulation/fibrinolysis are obvious and the direct action of PAR2 has already been partly elucidated (14).
This suggests that not only are cancer cells involved in local progression and angiogenesis (15,16), but host tissue PAR2 expression is also involved, and may directly contribute to cancer dissemination through increased release of tumorous cells from the primary tumor environment, or by promoting endothelial trapping and angiogenesis (3) in distant organs utilizing similar mechanisms. In the past, we entertained another explanation for the anti-metastatic effect and survival prolongation, which involved activation of a mixture of trypsin, chymotrypsin and papain by Par2 (17). Nevertheless, the absence of functional Par2 could still be an important factor influencing melanoma growth and in vivo spread, probably through tumor cell migration, invasiveness and metastasis formation. Moreover, just recently, the role of tissue factor/factor VIIa (TF/FVIIa) complex was found to be associated with PAR2-mediated tumor proliferation and progression (18). TF is expressed on many cancer cell types and its host- and tumor-originated contribution to disease progression probably depends on the cancer type itself. However, in murine models, TF induces cell proliferation rather than cell adhesion (19). Our results suggest that having a shared common activation pathway (i.e. TF/FVIIa/Par2), it is the host's Par2 expressed on endothelial cells that probably prevails in distant cell adhesion and metastasis. Involvement of other factors or enzymes in the coagulation cascade [like FXa in Par2 activation and the contribution of Par1 activation of endothelial cells (Par1 is not expressed on murine platelets)] must be considered in connection to host-related tumor progression in addition to TF/FVIIa activation.
Our results agree with previously published articles suggesting an important dual role for PAR2 in melanoma: This receptor contributes to the limitation of local cancer progression in one area, while on the other hand, is important for enhancing distant metastatic spread. Further studies using larger sample sizes are needed to comprehensively elucidate the role of this receptor as a potential therapeutic target in malignant melanoma.
Acknowledgements
The Authors would like to thank Tom Secrest for revisions of the English version of this article. This study was partly supported by grant no. P303/12/1791 to RM and no. 302/10/0346 to PJ by the Grant Agency of the Czech Republic, by the research project PRVOUK - Oncology P27, awarded by Charles University in Prague and the research project RVO67985823.
- Received March 21, 2014.
- Revision received May 23, 2014.
- Accepted May 26, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved