Article Text

Abstract
Aims Denosumab, a fully human monoclonal antibody directed against RANKL, has recently been introduced in the treatment strategy of giant cell tumour of bone (GCTB). Aim of this study was to investigate the phenotypical modifications induced by denosumab treatment in a series of 15 GCTB.
Methods The tumours were characterised for histone 3.3 mutations, and studied immunohistochemically for the modifications of RANKL, RANK, SATB2 and RUNX2 expression, as well as of tumour proliferative activity and angiogenesis.
Results Nine of 11 tumours investigated presented a histone 3.3 mutation in H3F3A, and 2 of these for which the analysis was carried out in pretreatment and post-treatment specimens showed the same mutation in both. Denosumab induced the disappearance of osteoclast-like giant cells, leaving residual spindle neoplastic cells arranged in a storiform pattern, with deposition of trabecular collagen matrix and osteoid, which tended to maturation in the peripheral portions of the lesion. RANK and RANKL expression was variable, with no significant variation after treatment. Moreover, we did not observe any significant modification of the expression of the osteoblastic markers SATB2 and RUNX2. Denosumab treatment determined a significant reduction of the proliferative index and of tumour angiogenesis (p=0.001, Wilcoxon rank-sum test).
Conclusions These results indicate that denosumab induces a partial maturation towards the osteoblastic phenotype of the neoplastic cells of GCTB, with production of fibrous and osteoid matrix, but with minor immunophenotypical changes. Finally, we first report an antiangiogenic activity of denosumab in GCTB, possibly mediated by a RANKL-dependent pathway.
- BONE TUMOURS
- IMMUNOCYTOCHEMISTRY
- CANCER GENETICS
Statistics from Altmetric.com
Introduction
Giant cell tumour of bone (GCTB) is a rare locally aggressive tumour, which most often affects young adults with predilection for the epiphyses of long bones.1 Histologically, the tumour consists of a proliferation of mononuclear cells, accompanied by a population of non-neoplastic osteoclast-like giant cells and mononuclear osteoclast precursors. Currently, it is thought that proliferating neoplastic cells produce a number of cytokines and mediators, including the receptor activator of nuclear factor κ-B-ligand (RANK-RANKL) system, that recruit osteoclast precursors and induce their maturation into multinucleated osteoclast. These cells ultimately determine bone destruction and allow the growth of the tumour. Recently, a putative driver mutation in the H3F3A gene of histone 3.3 has been identified as a specific genetic change of GCTB.2
The standard management of GCTB is based on surgery with several local adjuvant treatments like methacrylate cement, phenol or cryotherapy to reduce the risk of recurrence, while bisphosphonates are used in some cases to decrease bone resorption and for pain relief in inoperable tumours or metastatic disease.3 In the last 5 years the use of denosumab, a fully human monoclonal antibody already licensed for postmenopausal osteoporosis and prevention of skeletal related events in bone metastases from solid tumours, has been introduced in the treatment strategy of GCTB.4 This antibody specifically binds to RANKL, preventing its interaction with RANK and thus the formation of osteoclastic giant cells and the consequent excessive bone resorption. Denosumab has shown to be a turning point in the management of GCTB, allowing treatment of patients that cannot undergo surgery due to excessive morbidity for the anatomical location of the lesion and allowing a less morbid surgery after reduction of the osteolytic area, with general beneficial effects on patients’ symptoms.3 ,4
In this study we examined the histological and immunophenotypical features of a series of GCTB, characterised for histone 3.3 mutations, before and after denosumab administration, comparing baseline and resection specimens. In particular, we investigated the modifications in the expression of RANKL and RANK, and of the osteoblastic markers SATB2 and RUNX2. Moreover, we examined the effects of the treatment on tumour proliferative activity and on the angiogenesis through the determination of microvascular density (MVD).
Patients and methods
Patients
Clinical data of the 15 patients who received denosumab treatment preoperatively are summarised in table 1. All, but one, patients were affected by primary skeletal lesions. The only exception was a patient who presented a local soft tissue recurrence of a GCTB of the distal femur which had been treated 16 months before with curettage.
Summary of the clinical features of the series and mutation analysis
After screening for contraindications and informed consent acquisition, patients were treated preoperatively with denosumab 120 mg subcutaneously (days 1, 8, 15, 28 and every 4 weeks thereafter) for at least 3 months and then postoperatively every 4 weeks for 6 months. Timing of surgery was based on clinical and radiological findings; the median time on preoperative denosumab was 5.7 months (range 3–6). All patients underwent surgery, which consisted of curettage with local adjuvant treatments in nine cases and tumour resection in six cases. At a median follow-up of 13 months, three patients developed a local recurrence and are currently on treatment with denosumab at the same schedule. We did not report any serious adverse reaction to denosumab, mostly for the short length of the treatment over the time in our series.
Histone 3.3 mutation analysis
The thick tissue sections (7–10 μm) of formalin-fixed paraffin-embedded (FFPE) blocks were submitted to an overnight digestion with proteinase K at 56°C. DNA was extracted from FFPE tissues with the Qiamp DNA FFPE tissue kit (Qiagen, Hilden, Germany) accordingly to the manufacturer's protocol in a final volume of 60 µL. Nucleic acid samples were checked for concentration and quality by using the NanoDrop ND-1000 Spectrophotometer (Thermo Scientific, NYSE:TMO).
The primer sets were selected to cover the hot spot sites of genes and were as follows: H3F3A (NC_000001.11) forward primer 5′-tgtttggtagttgcatatggtga-3′ and reverse primer 5′-acaagagagactttgtcccatt-3′; H3F3B (NC_000017.11) forward primer 5′-ttatcttcggggcgtctttc-3′ and reverse primer 5′-gagcaggggaggagtgag-3′. The targeted sequences were tested to exclude non-specific amplification of homologous regions by University of California Santa Cruz (UCSC) Genome Browser—BLAT BLAST-like alignment tool Search (http://www.genome.ucsc.edu/cgi-bin/hgBlat?command=start) and Primer-BLAST (basic local alignment search tool) (http://www.ncbi.nlm.nih.gov/tools/primer-blast/).
Amplification of both genes was performed in an ABI 2720 thermal cycler (Applied Biosystems-Life Technologies, Foster City, California, USA) by starting from two different quantity of DNA (20 ng and 100 ng) in a final volume of 20 μl and using HotStarTaq DNA Polymerase (Qiagen, Milan, Italy) with 500 nM of each primer. The thermal protocol entailed an initial denaturation at 95°C for 5 min, followed by 40 cycles at 95°C for 20 s, 58°C for 30 s and 72°C for 30 s, with a final elongation step at 72°C for 10 min.
PCR products were purified by using FastGene Gel/PCR Extraction kit (Nippon Genetics Europe GmbH, Düren, Germany). DNA fragments were submitted to the cycle sequencing reaction with BigDye Terminator V.1.1 Cycle Sequencing Kit (Applied Biosystems-Life Technologies). The purified sequences, obtained by ZR DNA Sequencing Clean-up Kit (Zymo Research Corporation, Irvine, California, USA), were run on an ABI PRISM 310 Genetic Analyzer instrument (Applied Biosystems, Darmstadt, Germany) and manually analysed.
Sequence variants were reported according to the nomenclature edited by the Human Genome Variation Society (http://www.hgvs.org/mutnomen/recs-DNA.html) considering the nucleotide 1 as the A of the ATG-translation initiation codon.
Immunohistochemistry
For immunohistochemical staining, FFPE tissue sections (5 μm) were deparaffined, hydrated and after endogenous peroxidase inactivation immunostained with BenchMark Ultra stainer (Ventana, Tucson, Arizona, USA), than revealed with iVIEW DAB detection kit, providing a brown reaction product. Table 2 shows antibody source, dilution and antigen retrieval protocol. After completing the staining process, the slides were removed from the autostainer, counterstained with haematoxylin, dehydrated and mounted with a permanent medium. As negative control, we substituted primary antibody with a Ventana dispenser filled with non-immune serum at the same concentration for each immunohistochemical reaction. As positive control, we used reactive bone and bone fracture specimens.
List of the antibodies used in the present study
The results of the immunohistochemical staining were evaluated according to a semiquantitative scale as follows: −=0; +<10%; ++=11–49%; +++>50%, on baseline and on-study specimens. The Ki67 proliferation index was assessed in areas of greater expression determining the percentage of positive cells in at least 200 cells. Determination of tumoral neoangiogenesis in specimens stained with CD31 was performed according to the method of Weidner et al.5 MVD was assessed in areas containing the greatest number of vascular profiles (hot spots). The analysis was performed in three high power fields for each section of each tumour and the results expressed by average value.
Statistical analysis
All statistical tests were performed using SPSS software (release V.12.0). Wilcoxon signed-rank test was used to compare the proliferation index and MVD between baseline and on-study specimens. p Values <0.05 were considered statistically significant.
Results
Histone 3.3 mutation analysis
We performed direct sequencing for the presence of H3F3A (NM_002107.4) and H3F3B (NM_005324.4) variants in coding region between codons 1 and 42, including the hot spot codons (28, 35 and 37), on DNA extracted from FFPE tissue. Mutation analysis was carried out in 11 tumours for which non-decalcified specimens were available, and it was successfully performed in 9 tumours, while in 2 instances the quality of extracted DNA was not sufficient to complete the test. Eight tumours presented a p.Gly35Trp (p.G35W; NP_002098.1) mutation in the H3F3A gene, while a p.Gly35Val (p.G35V) was detected in one case (table 1). No mutation was identified in the H3F3B gene. In two cases it was possible to perform the analysis in pretreatment and post-treatment tumour samples, and the same p.Gly35Trp (p.G35W) mutation was detected in both specimens.
Histopathology
Post-treatment tumour samples showed pronounced changes in comparison with pretreatment samples, which mainly consisted of (1) disappearance of osteoclast-like giant cells, (2) presence of cellular areas formed by sheets of round to ovoid tumour cells, (3) cellular areas formed by spindle cells arranged in a storiform pattern with little or no extracellular matrix, (4) areas characterised by the production of abundant fibrillary extracellular matrix organised in trabecular structures or with a honeycomb pattern (figure 1). In resection specimens, these histological patterns were not haphazardly distributed, but showed the tendency towards a ‘zonal’ distribution, with more cellular areas in the tumour central portion, and matrix-rich areas in the periphery (figure 2). Moreover, at the periphery of the tumour, the tumour osteoid-like matrix seemed to merge with the host bone (figure 2).

Histological appearance of denosumab treated giant cell tumour of bone. The residual tumour is composed of bland-appearing spindle cells organised in short fascicles with a storiform pattern (A), associated with collagen matrix production. This matrix appears either as thin bands (B), or as thicker connected trabecular structures with a honeycomb appearance (C–D). Densely collagenised areas, almost devoid of tumour cells, are also seen (E). One tumour was completely necrotic (F).

Giant cell tumour of the proximal fibula after denosumab treatment. Whole tumour section (left) with corresponding enlarged details (right). The tumour centre is almost devoid of cells (A), while moving towards the periphery cellular areas with benign fibrous histiocytoma-like appearance are recognised (B), together with increasing amounts of matrix formation, which progressively assumes the appearance of osteoid matrix (C) and merges with the host cortical bone (D).
Further histological patterns observed in post-treatment samples included the presence of (1) groups of foamy histiocytes interspersed among tumour spindle cells, (2) focal areas of coagulative necrosis, (3) areas of round to oval acellular hyaline matrix, sometimes with a rosette-like appearance. In one case, treatment resulted in complete necrosis of the tumour (figure 1). One denosumab-treated soft tissue recurrence of GCTB consisted of a neoplastic population of spindle cells arranged in elongated fascicles, with scant extracellular matrix and presence of few osteoclast-like giant cells.
Immunohistochemistry
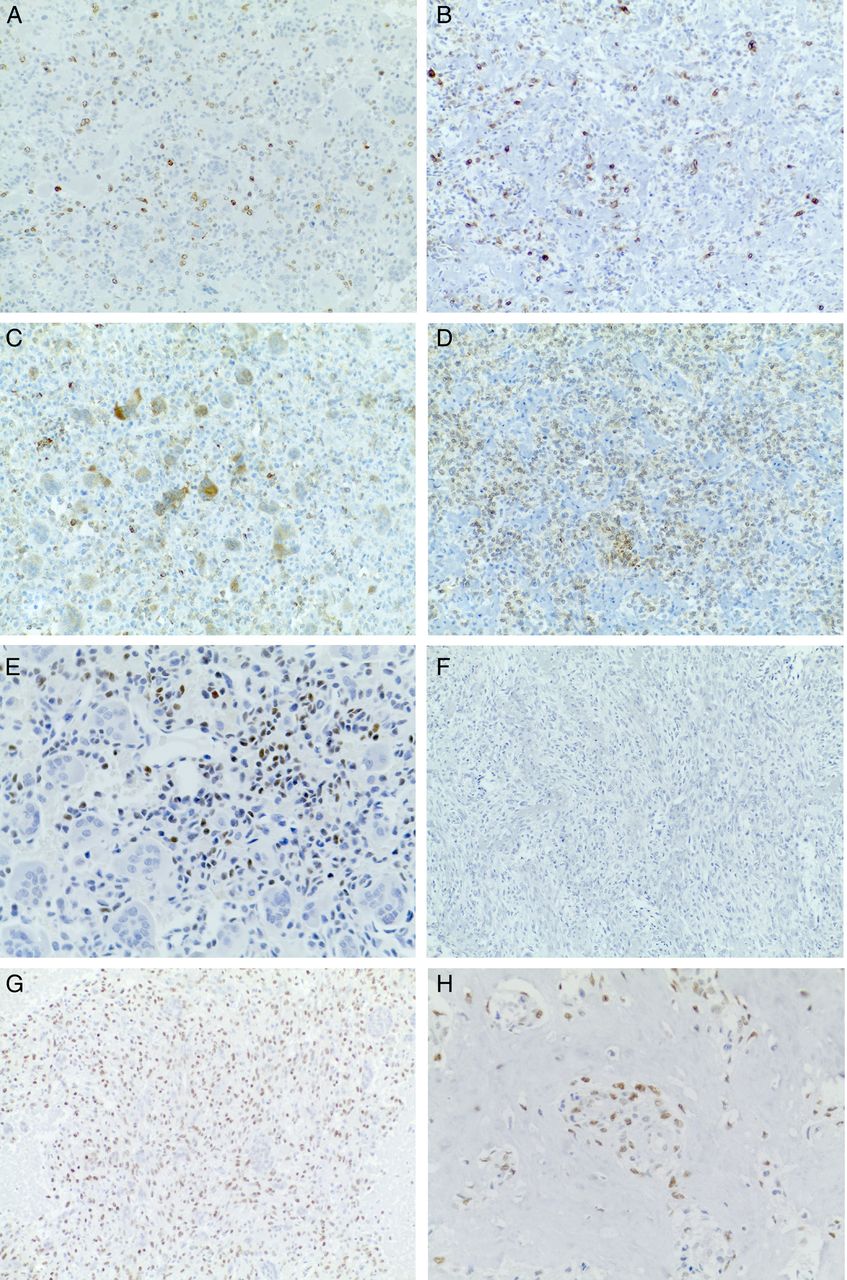
The results of the immunohistochemical studies are summarised in table 3 and illustrated in figure 3.
Summary of the results of the immunohistochemical studies conducted on 15 giant cell tumours of bone treated with denosumab

Representative images of the immunohistochemical studies. Images on the left column are from baseline specimens, while those on the right column are from on-study specimens. RANK is expressed by a subpopulation of mononuclear cells (A and B), while RANKL is expressed by multinucleated giant cells and mononuclear cells (C and D). Mononucleated cells show nuclear immunostaining for the osteoblastic marker SATB2 (E), while this marker is not expressed by residual cells after treatment in areas with no matrix formation or with collagen matric production (F). RUNX2 was extensively positive in pretreatment and post-treatment lesions (G and H).
Cytoplasmic positivity for RANKL was observed in mononuclear tumour cells in all baseline tumour samples, with a percentage ranging between 5% and 10% in most cases, while in four cases the staining was more diffuse. Interestingly positive cells were not uniformly distributed within the tumour population, but tended to group together. RANKL expression did not vary significantly in post-treatment samples, with the exception of three cases that showed reduced expression (from ++ to +), and one case that showed increased expression (from + to ++).
In baseline samples, RANK immunostaining was observed in the cytoplasm and/or membrane of osteoclast-like giant cells and in a fraction of mononuclear cells, ranging between 5% and 10%. In on-study samples RANK expression was variable, being absent in two cases, present in few cells in seven cases or present in most tumour cells in five cases.
Nuclear immunoreactivity for the osteoblastic marker SATB2 was observed in 12 of 15 pretreatment tumour samples. Positive cells were mononucleated and tended to group within the neoplastic population. Moreover, SATB2 positivity was associated with focal areas of osteoid matrix production in six tumours. In post-treatment samples SATB2, expression was detected in five tumours, in association with production of osteoid-like matrix, while cellular areas were always negative. All pretreatment tumour samples showed diffuse nuclear positivity for RUNX2, whereas in post-treatment specimens the expression was unchanged in 10 cases, or reduced to less than 10% of cells in 4 cases.
Evaluation of tumour angiogenesis and proliferative activity
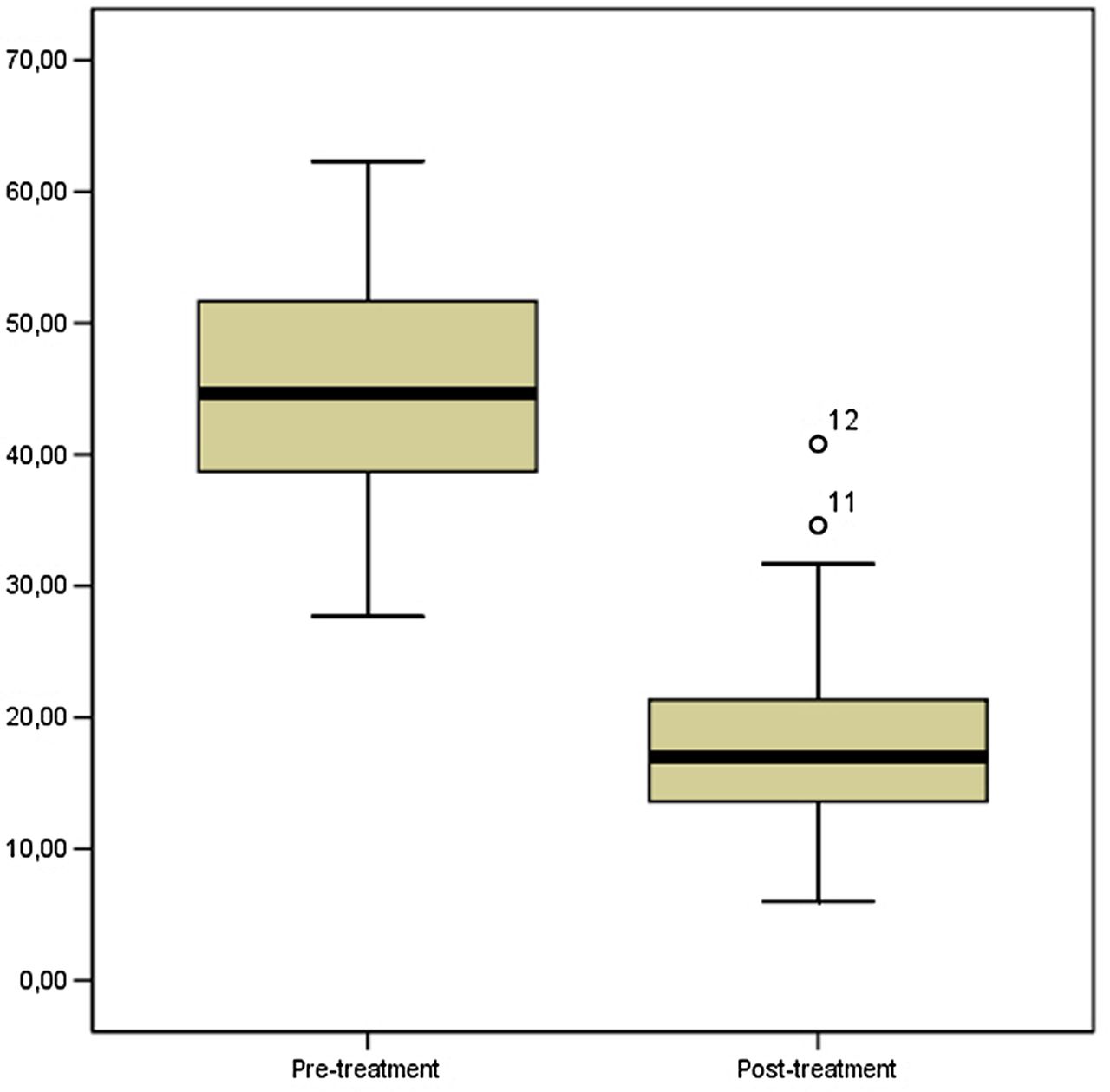
In the present series of GCTB, tumour angiogenesis evaluated as MVD decreased significantly after denosumab treatment. In baseline tumour samples mean MVD was 45.68±9.88 SD (range 27.67–62.33), while in on-study tumour samples it was 19.69±9.68 SD (range 6.00–40.80) (p=0.001 Wilcoxon signed-rank test). These results are shown in figure 4.

Denosumab treatment induced a significant reduction of microvessel density in the present series of giant cell tumour of bone (p=0.001, Wilcoxon signed-rank test).
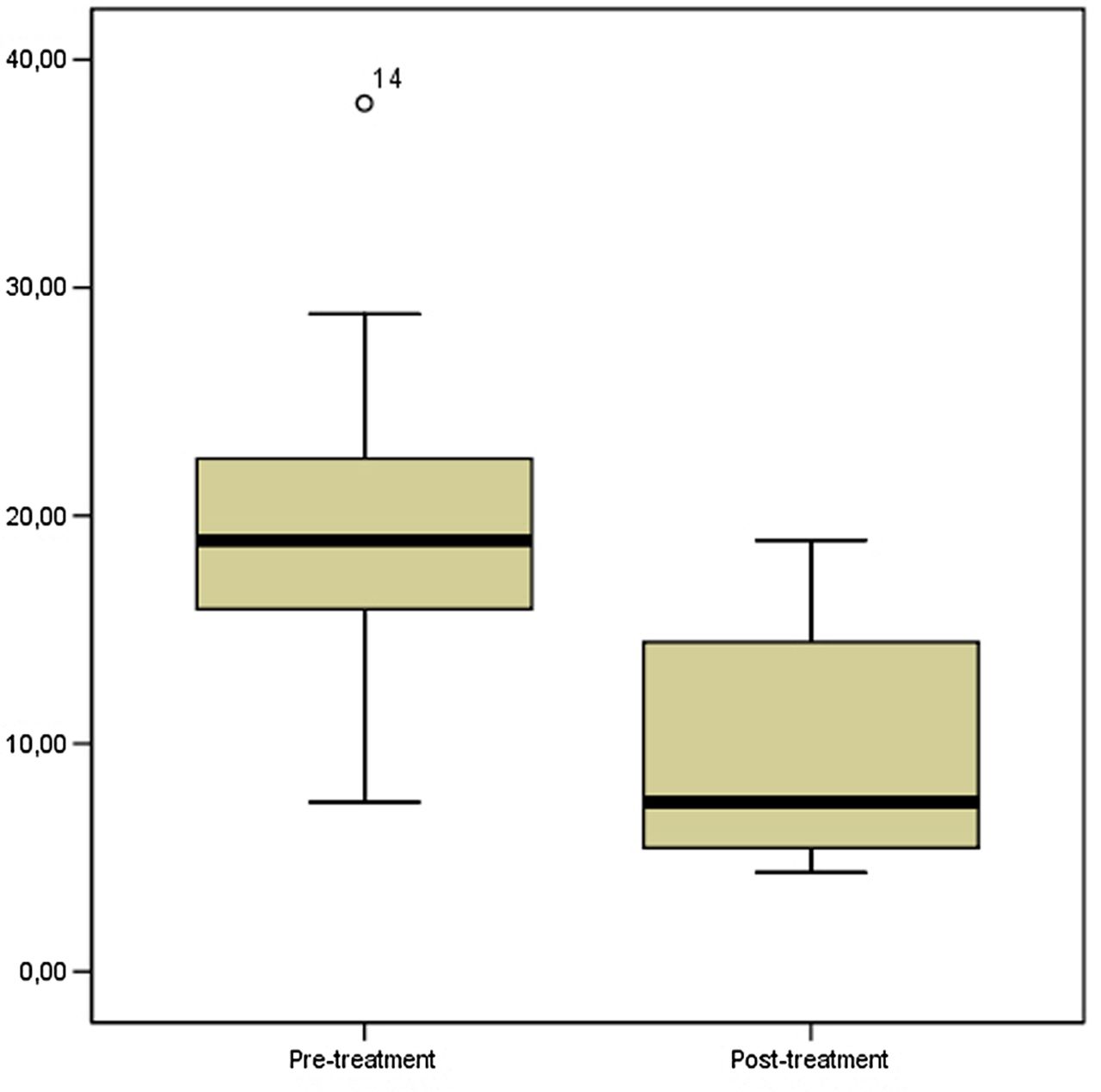
The Ki-67 labelling index ranged between 7.43% and 38.09% in baseline GCTB samples (mean 20.23±7.19 SD), while it decreased significantly in post-treatment samples, ranging between 4.34% and 18.92% (9.65±5.19 SD) (p=0.001 Wilcoxon signed-rank test). These results are illustrated in figure 5.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Proliferative activity determined as Ki67 labelling index was significantly lower in post-treatment specimens (p=0.001, Wilcoxon signed-rank test).
Discussion
The present study confirms that denosumab treatment induces deep changes in the morphology of GCTB.6–9 As in previous reports, we noted total or subtotal disappearance of osteoclast-like giant cells, and a residual tumour population formed mainly of spindle cells without atypia, arranged in fascicles, often with a storiform pattern, resulting in a fibrous histiocytoma-like appearance. Areas of collagen matrix formation, either with a diffuse or a honeycomb/trabecular appearance, merging with osteoid formation, were also visible. However, the analysis of whole tumour section revealed these areas were not haphazardly distributed, but they were rather organised according to a ‘zoning’ pattern, with more cellular areas in the tumour centre and matrix forming areas at the periphery.
Our series of GCTB was characterised at the molecular level for the presence of histone 3.3 mutations. These molecular alterations have been specifically associated with different bone tumour types, since GCTB presents usually G34W mutation (here reported in accordance with adopted nomenclature as p.Gly35Trp) in H3F3A, whereas H3 histone 3.3 M3F family 3B mutations have been detected only in a subset of cartilage tumours, including chondroblastoma and chondrosarcoma.2 The highly specific association of H3F3A mutation with GCTB supports its use as a possible diagnostic marker in histologically ambiguous cases, and to screen giant cell rich lesions of bone before denosumab treatment. In addition, it has been suggested that this histone 3.3 mutation may be relevant for the development of GCTB by inducing osteoclast recruitment, through the alteration of expression of essential osteoclast signalling pathways, such as RANK ligand or colony stimulating factor 1.2 In the present series, we confirm that GCTB presents mainly H3F3A mutations in glycine 34, with no involvement of the H3F3B gene. Moreover, we detected the same mutation in pre denosumab treatment and post denosumab treatment samples, supporting the hypothesis that this drug does not eliminate tumour cells, but it mainly modifies the microenvironment of the tumour by inhibiting the recruitment of osteoclast-like giant cells. Accordingly, we observed only minor modifications in the immunophenotype of tumour cells, at least regarding the expression of RANK, RANKL and osteoblastic markers.
Previous in vivo and in vitro studies have shown that GCTB stromal cells are, at least in part, of osteoblast lineage and express osteoblast and preosteoblast molecular markers.10–14 In order to assess whether denosumab treatment induces differentiation of the neoplastic cells of GCTB along the osteoblastic lineage, we examined the expression of RUNX2 and SATB2 in pretreatment and post-treatment tumour specimens. In agreement with previous studies,15 ,16 we observed that GCTB stains for both markers, although the staining for SATB2 tended to be less diffuse and presented in small groups of neoplastic cells, often associated with osteoid matrix production, whereas RUNX2 was expressed in most neoplastic cells. While RUNX2 expression remained unchanged after treatment, SATB2 was detected only in a subset of cells in areas of mature bone formation, and was not detected in cellular areas or in association with collagen matrix production. Considering that RUNX2 is highly expressed in the early phases of osteoblast differentiation and it is progressively reduced in the mature osteoblasts,17 while SATB2 is important for bone matrix production and mineralisation by mature osteoblasts,18–21 our results indicate that the neoplastic cells of GCTB may indeed have a intrinsically defective osteoblastic phenotype which is not or only partially associated with mature bone matrix production after denosumab treatment. In agreement with this hypothesis, high levels of RUNX2 expression have been associated with reduced osteocalcin production and the maintenance of an immature osteoblastic phenotype.17 ,22 ,23 However, the low expression of SATB2 in the postoperative specimens might also be explained considering that the median time on preoperative treatment in our series is 5.7 months which could be a short time for the acquisition of a mature osteoblastic phenotype with new woven bone formation.6
In the present study, we observed a significant reduction of proliferative activity after treatment, as indicated by the evaluation of the Ki67 labelling index. Similarly, Branstetter et al,6 reported a reduction of Ki67 positive tumour stromal cells, although they did not quantify the results. This effect is likely to be due to a modification of the microenvironment, rather than a direct effect, since in vitro studies have not demonstrated either a significant inhibitory effect on proliferative activity in GCTB cultures,24 or a permanent modification of their phenotype.25
Here, we first report that denosumab treatment has an antiangiogenic effect in GCTB in vivo, as shown by significantly reduced tumour microvessel density. However, the mechanisms responsible for this antiangiogenic effect need to be elucidated. Previous studies have shown that vascular endothelial growth factor (VEGF) induces RANK expression by endothelial cells, making them more responsive to RANKL effects.26 Thus, RANKL proangiogenic effect could be modulated by VEGF itself through the induction of RANK overexpression. Moreover, RANKL increases vascular permeability and angiogenesis through an interaction with endothelial nitric oxide synthase,27 independently from VEGF activity.28
In conclusion, our study confirms that denosumab treatment induces deep changes in the morphology and biology of GCTB, but does not eliminate tumour cells, as indicated by the persistence of a population with the same H3 histone H3F3A family 3A subunit mutation. The clinical benefit of this treatment appears to be related, at least in part, to a reduction of the proliferative activity and tumour angiogenesis.
Take home messages
Giant cell tumour of bone presents mainly H3F3A mutations in glycine 34, with no involvement of the H3F3B gene.
Denosumab treated giant cell tumours of bone show no significant modification of RANK and RANKL expression, and of the osteoblastic markers SATB2 and RUNX2.
Denosumab induces significant reduction of the proliferative index and of tumour angiogenesis.
References
Supplementary materials
Abstract in Italian
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Abstract in Italian - Online abstract
Footnotes
IG and IM contributed equally.
Handling editor Cheok Soon Lee
Contributors IG and AF: reviewed the histological slides, including immunohistochemistry. IM: wrote the manuscript. LS: performed the molecular studies and reviewed the manuscript. AS: performed the immunohistochemical studies and reviewed the manuscript. GGB, DC, GB, GS, RC and AD: provided clinical data and reviewed the manuscript.
Competing interests None declared.
Ethics approval Institutional Ethics Committee.
Provenance and peer review Not commissioned; externally peer reviewed.