


Abstract
Determinants of differentiation and apoptosis induction by the novel histone deacetylase inhibitor (HDACI) LAQ824 were examined in human leukemia cells (U937 and Jurkat). Exposure of U937 cells to a low concentration of LAQ824 (30 nM) resulted in a delayed (2 h) increase in reactive oxygen species (ROS), induction of p21WAF1/CIP1, pRb dephosphorylation, growth arrest of cells in G0/G1 phase, and differentiation. On the other hand, exposure of cells to a higher concentration of LAQ824 (75 nM) resulted in the early (30 min) generation of ROS, arrest of cells in G2/M phase, down-regulation of XIAP (at the transcriptional level) and Mcl-1 (through a caspase-mediated process), the acid sphingomyelinase-dependent generation of ceramide, and profound mitochondrial injury, caspase activation, and apoptosis. LAQ824-induced lethality in U937 cells did not involve the extrinsic apoptotic pathway, nor was it associated with death receptor up-regulation; instead, it was markedly inhibited by ectopic expression of Bcl-2, Bcl-xL, XIAP, and Mcl-1. The free radical scavenger N-acetyl cysteine blocked LAQ824-mediated ROS generation, mitochondrial injury, Mcl-1 down-regulation, ceramide generation, and apoptosis, suggesting a primary role for oxidative injury in LAQ824 lethality. Together, these findings indicate that LAQ824-induced lethality represents a multifactorial process in which LAQ824-mediated ROS generation is necessary but not sufficient to induce apoptosis, and that the degree of XIAP and Mcl-1 down-regulation and ceramide generation determines whether this agent engages a maturation rather than an apoptotic program.
Attention has recently been focused on the role of histone deacetylase inhibitors (HDACIs) as antineoplastic agents, and several of these compounds are currently in various stages of clinical development (Rosato and Grant, 2004). HDACIs comprise a diverse group of agents that act by promoting histone hyperacetylation. Histone acetylation is maintained by the net balance between the opposing actions of histone acetyltransferases and histone deacetylases (HDACs), and represents an important mechanism by which chromatin remodeling and transcriptional activity are regulated (Gray and Teh, 2001). The mechanism of action and the basis for selectivity of HDAC inhibitors toward tumor cells have not yet been fully elucidated, but multiple possibilities exist. For example, HDACIs may interfere with cancer cell proliferation and/or survival through the modulation of cell-cycle traverse, induction of differentiation, or by promoting cell death (i.e., apoptosis) (Rosato and Grant, 2004). In human leukemia cells, HDACIs, when administered at low concentrations, trigger maturation, whereas at higher concentrations, apoptosis ensues (Rosato et al., 2001, 2003b). The factors that determine whether HDACIs engage apoptosis versus differentiation remain the subject of investigation, but they may involve the extent of mitochondrial injury or oxidative damage, up-regulation of death receptors, activation of the extrinsic pathway, interference with 90-kDa heat shock protein function, down-regulation of antiapoptotic Bcl-2 family members, and the generation of ceramide, among others (Yu et al., 2002; Kurita-Ochiai et al., 2003; Rosato et al., 2003b; Guo et al., 2004; Maggio et al., 2004). Recent reports have suggested that the induction of death receptors and perturbations in cellular redox status may underlie the selectivity of these agents toward neoplastic cells (Insinga et al., 2005; Ungerstedt et al., 2005). A variety of compounds with HDAC inhibitory activity have now been identified. On the basis of their structural features, they have been subdivided into four basic groups: 1) short-chain fatty acids (e.g., sodium butyrate, AN-9, phenylbutyrate, phenylacetate, and valproic acid); 2) synthetic benzamide derivatives (e.g., MS-275 and CI-994); 3) cyclic tetrapeptides (e.g., depsipeptide, trapoxin, and apicidin); and 4) hydroxaminic acids (suberoylanilide hydroxamic acid, pyroxamide, trichostatin A, oxamflatin, 3-chloro-4-hydroxyphenylacetates, and LAQ824) (Rosato and Grant, 2004).
NVP-LAQ824 is a structurally novel cinnamic hydroxamic acid that exhibits potent in vitro activity when administered at submicromolar concentrations (Atadja et al., 2004a). LAQ824 has been shown to induce apoptosis in multiple myeloma cell lines resistant to more conventional therapeutic agents (Catley et al., 2003) and in Bcr/Abl+ human leukemia cells resistant to imatinib mesylate (Nimmanapalli et al., 2003). LAQ824 is also active against several additional neoplastic cell types, including breast (Fuino et al., 2003), colon, non-small-cell lung carcinoma, prostate cancer (Atadja et al., 2004a), and other leukemia cells (Guo et al., 2004). In addition to its direct cytotoxic actions, LAQ824 can sensitize transformed cells to other agents. For example, in breast cancer cells, LAQ824 depleted Her-2 mRNA and protein levels and significantly increased the lethality of trastuzumab, taxotere, gemcitabine, and epothilone B (Fuino et al., 2003). In both human SKW6.4 and Jurkat leukemia cells, LAQ824 induced an increase in the expression levels of TRAIL death receptors DR4 and DR5 and down-regulation of FLICE-inhibitory protein, events which sensitize cells to subsequent TRAIL/Apo2-L exposure (Guo et al., 2004).
Relatively little is currently known about the factors that determine whether LAQ824 induces a maturation program rather than apoptosis in human leukemia cells. To address this question, we have examined the dose-dependent induction of apoptosis by LAQ824 in human myelomonocytic U937 and Jurkat T-human lymphoblastic leukemia cells. Here, we report that low concentrations of LAQ824 trigger cell-cycle arrest in G1 phase, most likely through up-regulation of the cyclin-dependent kinase inhibitor p21WAF1/CIP1, the relatively delayed generation of reactive oxygen species (ROS), and cellular maturation. In contrast, higher concentrations of LAQ824 primarily induce apoptosis, an event associated with early ROS generation, G2M arrest, induction of mitochondrial injury, down-regulation/cleavage of antiapoptotic proteins including Bcl-2, Bcl-XL, and Mcl-1, transcriptional repression of XIAP, and the acid sphingomyelinase-dependent generation of the proapoptotic lipid second-messenger ceramide.
Materials and Methods
Cells and Cell Culture. U937 and Jurkat human leukemia cells were obtained from American Type Culture Collection (Manassas, VA). Cells were cultured and maintained in logarithmic growth phase in RPMI 1640 medium supplemented with sodium pyruvate, minimal essential medium essential vitamins, l-glutamate, penicillin, streptomycin, and 10% fetal bovine serum (Invitrogen, Carlsbad, CA). U937 cells stably overexpressing Bcl-2, Bcl-XL, FADD-DN, or XIAP and their empty-vector counterparts were obtained as reported previously (Datta et al., 2000; Cartee et al., 2002) and maintained as described above in the presence of the corresponding selection antibiotics. All experiments were performed using cells in logarithmic phase growth-suspended at 2.5 × 105 cells/ml.
Drugs and Chemicals. LAQ824 was kindly provided by Novartis Pharmaceuticals Inc. (Basel, Switzerland). The pan-caspase inhibitor Boc-d-fluoromethyl ketone was purchased from Enzyme System Products (Livermore, CA) and dissolved in dimethyl sulfoxide. The ceramide synthesis inhibitors were from Calbiochem (San Diego, CA; GW4869), BIOMOL Research Laboratories (Plymouth Meeting, PA; fumonisin B1 and myriocin), and Sigma-Aldrich (St. Louis, MO; desipramine).
Morphological Assessment of Apoptosis. Apoptotic cells were evaluated by both morphological assessment of Wright-Giemsa-stained cytospin preparations and by annexin V/propidium iodide (PI) staining as described previously (Almenara et al., 2002). Annexin V/PI (BD Biosciences PharMingen, San Diego, CA) analysis of cell death was carried out according to the manufacturer's instructions.
Cell-Cycle Analysis. Analysis of cell cycle was made by flow cytometry as described previously (Rosato et al., 2001) using a BD Biosciences FACScan flow cytometer and Verity Winlist software (Verity Software, Topsham, ME).
Assessment of Mitochondrial Membrane Potential. At the indicated intervals, cells were harvested and 2 × 105 cells were incubated with 40 nM DiOC6 for 15 min at 37°C. Analysis was then carried out on BD Biosciences FACScan cytofluorometer. The percentage of cells exhibiting low levels of DiOC6, reflecting loss of mitochondrial membrane potential, was determined as described previously (Almenara et al., 2002).
Determination of Clonogenicity. Pelleted cells were washed extensively and prepared for soft-agar cloning as described previously (Maggio et al., 2004). Cultures were maintained for 10 to 12 days in a 37°C, 5% CO2 incubator after which colonies, defined as groups of ≥50 cells, were scored.
Surface Expression Analysis of DR4 and DR5. After treatment, U937 cells (1 × 106) were pelleted, washed with PBS, resuspended, and blocked for 30 min with 1% bovine serum albumin in PBS and then incubated with 1 μg of antibodies against DR4, DR5, DcR1, or DcR2 (Alexis Corporation, San Diego, CA) in 1% bovine serum albumin in PBS for 30 min, followed by washing with PBS. Finally, cells were incubated with fluorescein isothiocyanate-conjugated goat antimouse IgG (DakoCytomation Denmark A/S, Glostrup, Denmark) for 30 min. After washes, cells were analyzed on a FACScan flow cytometer.
Western Blot Analysis. Whole-cell pellets were washed twice in PBS, resuspended in PBS, and lysed by the addition of one volume of loading buffer (Invitrogen), as described previously (Almenara et al., 2002). total proteins (30 μg per point) were separated by 4 to 12% Bis-Tris NuPage precast gel system (Invitrogen) and electroblotted to nitrocellulose. After incubation with the corresponding primary and secondary antibodies, blots were developed by enhanced chemiluminesence (PerkinElmer Life and Analytical Sciences, Boston, MA). Where indicated, blots were stripped and reprobed with antibodies directed against actin. Cytosolic proteins were obtained by using a technique described previously (Almenara et al., 2002) in which the S-100, or cytosolic fraction, was prepared as described with minor modifications. For each condition, 30 μg of the S-100 fraction was loaded onto the gel and probed with the corresponding antibody.
Antibodies for Western Blot Analysis. Primary antibodies for the following proteins were used at the designated dilutions: Bcl-XL(1:1000; Trevigen, Gaithersburg, MD); PARP (1:1000; BIOMOL); pro-caspase 3, cytochrome c, cyclins D1, A, and E; Mcl-1, pRb and underphosphorylated Rb, and XIAP (1:1000, BD Biosciences PharMingen); Bcl-2 (1:2000; DAKO); caspase 8 (1:2000 Alexia Corporation); Bid (1:1000; Cell Signaling Technology Inc., Beverly, MA); actin (1:2000; Sigma-Aldrich); and Smac/DIABLO and AIF (1:1000, Santa Cruz Biotechnology, Santa Cruz, CA). Secondary antibodies conjugated to horseradish peroxidase were obtained from Kirkegaard and Perry Laboratories (Gaithersburg, MD).
Ceramide Determination. Ceramide was assayed as described previously (Maggio et al., 2004). In brief, after various treatments, lipids were extracted, and mass amounts of ceramide in cellular extracts were measured by the diacylglycerol kinase enzymatic method. Ceramide levels were normalized to total phospholipids present in the extracts.
Real-Time Reverse Transcriptase-Polymerase Chain Reaction. Experiments were performed using the ABI Prism 7900 Sequence Detection System (Applied Biosystems, Foster City, CA) using the TaqMan One Step PCR Master Mix Reagents Kit as described previously (Rosato et al., 2004).
Statistical Analysis. The significance of differences between experimental conditions was determined using the Student's t test for unpaired observations. To assess the interaction between agents, median dose analysis (Maggio et al., 2004) was used with a commercially available software program (CalcuSyn; Biosoft, Ferguson, MO). The combination index was calculated for a two-drug combination involving a fixed concentration ratio. Using these methods, combination index values less than 1.0 indicate a synergistic interaction.
Results
LAQ824 Induces Concentration-Dependent Growth Arrest or Cell Death in Human Leukemia Cells. Whereas concentrations of LAQ824 ≤10 nM had no effect on the number of cells compared with controls, exposure of to 30 to 75 nM LAQ824 induced partial to essentially complete cell growth inhibition, respectively (Fig. 1A). A marked increase in apoptosis was noted at LAQ824 concentrations ≥50 nM, with maximal effects observed after 48-h exposure to 75 nM LAQ824. In contrast, LAQ824 concentrations ≤30 nM resulted in minimal toxicity (Fig. 1B). A clear increase in the percentage of dead cells 24 h after treatment with 75 nM LAQ824 was noted (data not shown), which correlated with cleavage/activation of Bid, caspases-3 and -8, and PARP degradation (Fig. 1C). Results in Jurkat T-human lymphoblastic leukemia cells were similar [i.e., cell growth inhibition at lower drug concentrations (20 nM) and cell death at higher LAQ824 concentrations (50 nM; data not shown)].

LAQ824 induces dose-dependent growth arrest or cell death in human leukemia cells. A, U937 cells in logarithmic phase were suspended at 2.5 × 105 cells/ml in the presence of LAQ824 at the indicated concentrations and were assayed at 24, 48, and 72 h by monitoring the number of cells after treatment using a Beckman Coulter Counter (Beckman Coulter, Fullerton, CA). B, the extent of apoptosis was determined by annexin V/PI analysis as described under Materials and Methods. C, Western blot analysis of lysates of U937 cells exposed to 30 or 75 nM LAQ824 collected at the indicated time intervals. After treatment, cells were pelleted, lysed, and 30 μg of protein was separated by SDS-PAGE as described under Materials and Methods. Blots were then probed with the indicated antibodies directed against PARP, caspases-3 and -8, and Bid. Blots were subsequently stripped and probed with antibodies directed against tubulin to ensure equivalent loading and transfer. The results of a representative study are shown; two additional experiments yielded similar results. D, lysates from U937 cells exposed to either 30 or 75 nM LAQ824 for 24 h were analyzed by Western blot, as described above. E, after incubation with the indicated concentration of LAQ824 for 72 h, the percentage of U937 cells expressing the surface differentiation markers Cd11a and Cd11b was determined by flow cytometry as described under Materials and Methods. Values represent the means ± S.D. for three separate experiments performed in triplicate. F, U937 cells were exposed to either 30 or 75 nM LAQ824 for the indicated intervals after which cells were washed free of drugs and plated in soft agar as described under Materials and Methods. Colonies, consisting of groups of ≥50 cells, were scored after 10 to 12 days.
A low concentration of LAQ824 (30 nM) induced an accumulation of cells in G0/G1 phase at 48 h (57.4 ± 2.6 versus 41.2 ± 2.1%; P < 0.01), a decrease in the S-phase fraction (31.9 ± 1.4 versus 46.8 ± 2.4%; P < 0.01), and no changes in the G2/M phase population. In contrast, G0/G1 arrest was not readily apparent in cells exposed to 75 nM LAQ824, because the large majority of cells were apoptotic by 48 h. It is noteworthy that cells exposed to a lethal LAQ824 concentration displayed a large accumulation in G2M at 24 h (e.g., 39.5 ± 1.8%), whereas this was not observed in cells exposed to 30 nM LAQ824 (data not shown). Western blot analysis revealed a robust concentration-dependent increase of the CDK inhibitor p21WAF1/CIP1 accompanied by increased levels of the hypophosphorylated forms of pRb (Fig. 1D). Bands corresponding to pRb cleavage fragments were also noted, particularly at higher LAQ824 concentrations. It is noteworthy that 30 nM LAQ824 induced a pronounced increase in the expression of the maturation markers CD11a and CD11b, whereas no changes were observed in cells exposed to 75 nM LAQ824 (Fig. 1E). Both differentiation and apoptosis may limit colony formation; furthermore, the extent of apoptosis does not necessarily correlate with the loss of clonogenicity (Yin and Schimke, 1995). Analysis of clonogenic survival showed a marked reduction in clonogenicity in cells exposed to 30 nM LAQ824 (i.e., from 28.3 to 21.3% of controls for 24- or 72-h exposure intervals, respectively), effects that were more pronounced in cells treated with 75 nM LAQ824 (i.e., 3.5 or 0.04% of control values for 24- or 72-h exposure intervals, respectively; Fig. 1F). Thus, despite exerting similar effects on p21WAF1/CIP1 and pRb, the antiproliferative activity of a low concentration of LAQ824 stems primarily from cell-cycle arrest in G0/G1 phase and cellular maturation, whereas higher LAQ824 concentrations largely reflect caspase activation and apoptosis.
LAQ824-Induced Cell Death Does Not Involve Activation of the Extrinsic, Receptor-Mediated Apoptotic Pathway in U937 Cells. LAQ824 has been shown to enhance the expression of death receptors DR4 and DR5 in both Jurkat T-lymphoblastic and SKW6.4 B-lymphoblastic leukemia cells (Guo et al., 2004). To determine whether LAQ824 lethality in U937 cells involved a similar mechanism, U937 and Jurkat cells were treated with 30 or 75 nM LAQ824 for 24 h in the presence or absence of either anti-TRAIL or anti FAS-L antibodies. LAQ824-induced lethality was blocked by the anti-TRAIL antibody in Jurkat cells (Fig. 2A); in contrast, no inhibitory effects were observed in U937 cells, nor were changes observed in either cell line after coincubation with anti-FAS-L antibody. Analysis of DR5 expression by flow cytometry demonstrated a clear concentration-dependent increase in DR5 expression in Jurkat cells and, consistently, no changes in U937 cells (Fig. 2B), suggesting that LAQ824-mediated up-regulation of the death receptor pathway represents a cell-type-specific phenomenon and is unlikely to be involved in LAQ824 lethality in U937 cells.
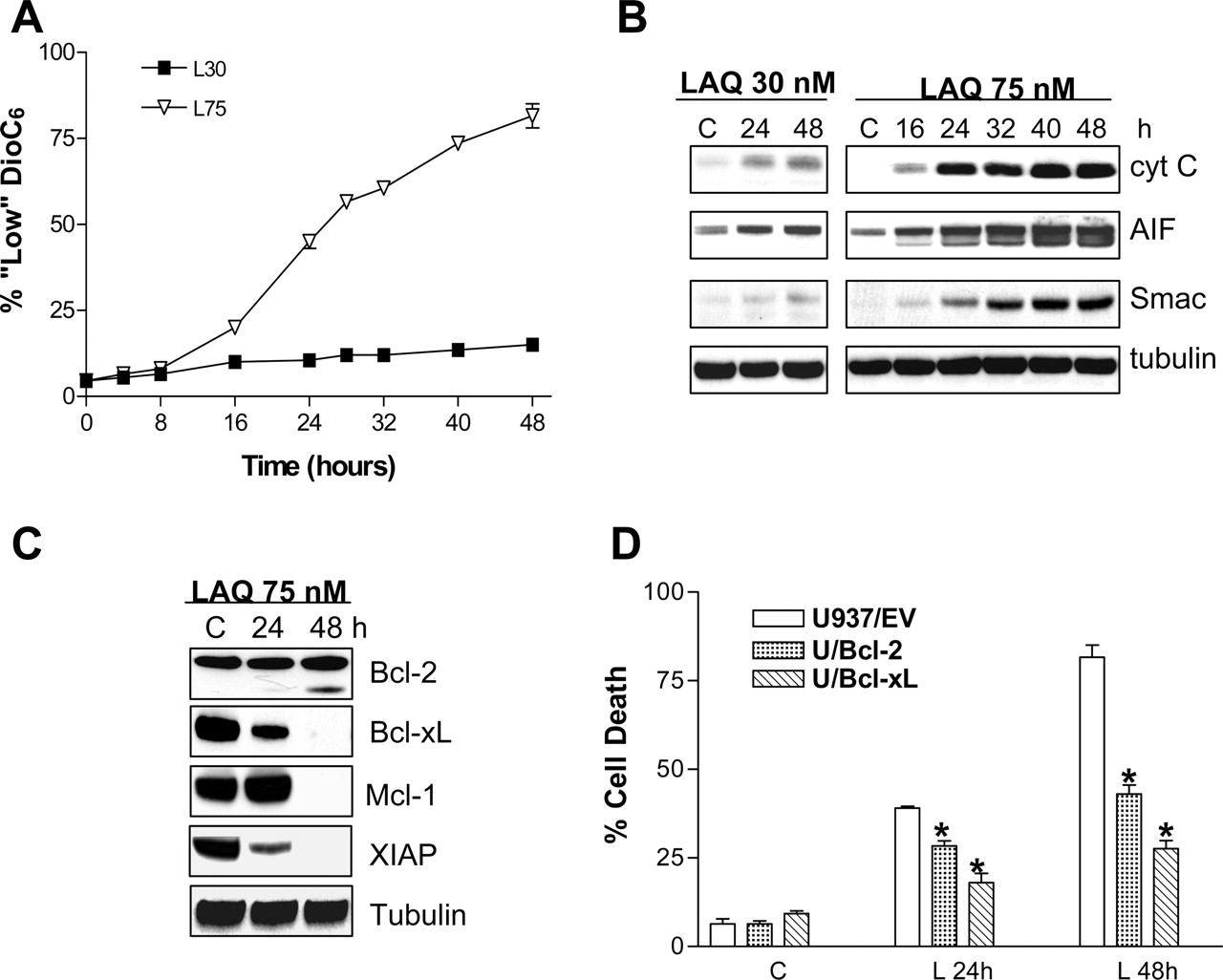
Exposure of U937 Cells to a Lethal Concentration of LAQ824 Results in Pronounced Mitochondrial Injury and Down-Regulation of Antiapoptotic Proteins. As shown in Fig. 3A, 75 nM LAQ824 induced a marked time-dependent loss of mitochondrial membrane potential (ΔΨm) beginning at 16 h of drug exposure, whereas 30 nM LAQ824 had little effect. Loss of ΔΨm was accompanied by cytoplasmic release of cytochrome c, AIF, and Smac/DIABLO after exposure to 75 nM LAQ824, whereas effects were much less pronounced in cells exposed to 30 nM LAQ824 (Fig. 3B). Furthermore, exposure to 75 nM LAQ824 for 48 h resulted in a Bcl-2 cleavage product, which has been associated with proapoptotic activity (Cheng et al., 1997) and down-regulation of Bcl-xL and Mcl-1 (Fig. 3C). Levels of XIAP were markedly reduced after 24 h of drug exposure as well and were essentially absent by 48 h. Furthermore, ectopic expression of either Bcl-2 or Bcl-xL (Rosato et al., 2003a) significantly protected U937 cells from LAQ824-mediated lethality at both 24 and 48 h (P < 0.01; Fig. 3D). Together, these findings indicate that in U937 cells, exposure to a lethal concentration of LAQ824 (75 nM) primarily induces apoptosis via the intrinsic, mitochondrial pathway.
Generation of ROS Plays a Critical Functional Role in LAQ824-Mediated Lethality. The generation of ROS has been shown to represent a primary event in the lethality of several HDACIs, including suberoylanilide hydroxamic acid, MS-275, and butyrate (Ruefli et al., 2001; Kurita-Ochiai et al., 2003; Rosato et al., 2003b). Consequently, ROS levels were monitored in U937 and Jurkat cells exposed to 30 or 75 nM LAQ824. Exposure to either 30 or 75 nM LAQ824 for 2 h induced similar increments in ROS levels in U937 cells (Fig. 4A). However, ROS generation was observed after 30 min of treatment with 75 nM LAQ824 but only after 2 h of exposure to 30 nM LAQ824 (data not shown). Similar effects were detected in Jurkat cells treated with 50 nM LAQ824 in which an early ROS peak was also observed (Fig. 4A). To determine the functional contribution of the early increase in ROS to lethality, U937 and Jurkat cells were exposed to LAQ824 in the absence or presence of the free radical scavenger N-acetyl cysteine (NAC). As shown in Fig. 4A, coexposure of cells to NAC blocked LAQ-induced ROS production. Moreover, treatment of either U937 (Fig. 4B) or Jurkat cells (data not shown) with LAQ824 in the presence of NAC essentially abolished mitochondrial injury (i.e., loss of ΔΨm) and apoptosis. Moreover, cells treated with LAQ824 plus NAC displayed a marked reduction in activation of caspases-3, -8, and Bid, PARP degradation, and release of mitochondrial proteins into the cytoplasm (data not shown). Furthermore, cleavage of Bcl-2 and Bcl-xL and Mcl-1 down-regulation were essentially abrogated by NAC, suggesting that these were secondary, caspase-dependent events. In contrast, XIAP expression was only modestly restored by NAC. The finding that both 30 and 75 nM concentrations of LAQ824 induced a similar increase in ROS levels suggests that this event may be necessary but not sufficient for LAQ824-mediated lethality.

LAQ824-induced cell death does not involve activation of the extrinsic, receptor-mediated apoptotic pathway in U937 cells. A, U937 and Jurkat cells were coincubated with LAQ824 (75 and 50 nM, respectively) in the presence or absence of either anti-TRAIL or anti-FAS-L antibodies, after which the percentage of apoptotic cells was monitored by annexin V/PI staining. B, Jurkat and U937 cells were treated as above, after which expression of DR5 was monitored by flow cytometry as described under Materials and Methods. Results are representative of three separate studies.
Functional Role of XIAP and Mcl-1 Down-Regulation in LAQ824-Induced Lethality. Studies were then undertaken to investigate the functional role of both XIAP and Mcl-1 down-regulation in LAQ824-mediated lethality. Time-course analysis of XIAP expression in U937 cells by Western blot demonstrated a marked reduction in protein expression after treatment of cells with 75 nM LAQ824, which approached ∼90% at 48 h (Fig. 5A). The presence of an XIAP cleavage fragment was also detected at later intervals (Fig. 5A), which may explain the partial restoration of protein levels by NAC (Fig. 4C). In contrast, 30 nM LAQ824 had no effect on XIAP expression. XIAP antiapoptotic activity is exerted in part by blocking the actions of caspases-3, -7, and -9 (Deveraux et al., 1998). XIAP down-regulation mediated by 75 nM LAQ824 was consistently correlated closely with activation of caspases-9, -3, and -7, whereas no activation was observed in cells exposed to 30 nM LAQ824. Enforced expression of XIAP (Datta et al., 2000) significantly blocked LAQ824-induced lethality at the 24- and 48-h intervals (P < 0.02), suggesting that XIAP down-regulation plays an important functional role in LAQ824 lethality. LAQ824 effects on the transcriptional regulation of XIAP were then examined using real time RT-PCR analysis (Fig. 5C). LAQ824 resulted in pronounced transcriptional inhibition of XIAP gene expression (i.e., approximately 50% after 24 h and 90% after 36-48 h; Fig. 5C). Moreover, levels of XIAP mRNA were only partially restored by blocking apoptosis with NAC, further demonstrating that LAQ824-induced XIAP mRNA down-regulation does not solely reflect a secondary, apoptosis-mediated event. In contrast, 30 nM LAQ824 had no effect on XIAP expression at either the protein (Fig. 5A) or mRNA level (Fig. 5C).

Effects of LAQ824 exposure on mitochondrial integrity and antiapoptototic proteins. A, U937 cells were incubated with either 30 or 75 nM LAQ824 for the indicated intervals, after which the percentage of cells exhibiting a loss of ΔΨm was determined by flow cytometry as described under Materials and Methods. Values represent the means for three separate experiments ± S.E.M. B and C, U937 cells were treated as in A: cells were pelleted, lysed, and protein was extracted from either the cytosolic S-100 fraction (AIF, cytochrome c, and Smac; shown in B) or whole-cell lysates (Bcl-2, Bcl-xL, Mcl-1, and XIAP; shown in C). In each case, 30 μg of protein was separated by SDS-PAGE, after which blots were probed with the corresponding antibodies. Blots were subsequently stripped and probed with antibodies directed against tubulin to ensure equivalent loading and transfer. The results of a representative study are shown; two additional experiments yielded similar results. D, U937 cells transfected with an empty vector (EV) or plasmids encoding Bcl-2 or Bcl-XL were incubated with 75 nM LAQ824 for 24 and 48 h, after which the percentage of annexin V/PI cells was determined as described. Values represent the means ± S.D. for three separate experiments performed in triplicate. *, significantly less than values obtained for empty vector control cells, P > 0.05 and >0.01 for L24 and L48, respectively.

Exposure of U937 cells to LAQ824 induces the generation of reactive oxygen species. A, both U937 (top) and Jurkat (bottom) cells were incubated with LAQ824 (30 and 75 nM in U937 cells; 50 nM in Jurkat cells) ± the free radical scavenger NAC (15 mM) for various intervals as indicated, after which they were labeled with an oxidative-sensitive dye (H2DCFDA) and analyzed by flow cytometry to determine the percentage of cells displaying an increase in ROS production (reflected by a rightward shift of the histogram). B, U937 cells were treated for 48 h with LAQ824 as described above in the absence or presence of 15 mM NAC. At the end of the incubation period, the percentage of cells exhibiting the loss of ΔΨm or apoptosis (annexinV/PI+) was determined as described under Materials and Methods. Values represent the means ± S.D. for three separate experiments performed in triplicate. *, significantly less than values obtained for cells treated with LAQ824 in the absence of NAC, P < 0.001. C, in parallel, whole lysates from U937 cells incubated with 75 nM LAQ824 for 48 h in the presence (L48/N) or absence (L48) of NAC were prepared and analyzed by Western blot as described. In each case, 30 μg of protein was separated by SDS-PAGE, after which blots were probed with the corresponding antibodies. Blots were subsequently stripped and reprobed with antibodies directed against tubulin to ensure equivalent loading and transfer. The results of a representative study are shown; two additional experiments yielded similar results. L, incubated in the presence of LAQ824; L48/N, incubated with LAQ824 for 48 h in the presence of NAC.

Role of XIAP down-regulation in LAQ824-induced lethality. A, U937 cells were exposed to either 30 or 75 nM LAQ824 for the indicated intervals after which cells were collected, lysed, and analyzed by Western blot. In each case, 30 μg of protein were separated by SDS-PAGE, blotted, and probed with the corresponding antibodies. The results of a representative study are shown; two additional experiments yielded similar results. Values shown represent the percentage of protein expression determined by densitometry relative to control values (100%). B, both U937 cells overexpressing a cDNA encoding XIAP (U937/XIAP) or the corresponding empty vector (U937/EV) cells were exposed for 24 and 48 h to 75 nM LAQ824 after which the percentage of apoptotic cells (annexinV/PI+) was determined by flow cytometry as described under Materials and Methods. Values represent the means ± S.D. for three separate experiments performed in triplicate. C, U937 cells were exposed to LAQ824 as above, after which expression of XIAP mRNA and 18S rRNA levels (used as a normalization parameter to ensure equivalent loading as specified by the manufacturer) were monitored by RT-PCR as described under Materials and Methods. Values for each condition are expressed as nanograms of specific XIAP mRNA per microgram or total RNA. Values represent the means ± S.D. for three separate experiments performed in triplicate.
Analysis of Mcl-1 expression showed dramatic down-regulation after 32 h exposure to 75 but not 30 nM LAQ824 (Fig. 6A), accompanied by the appearance of a ∼27-kDa cleavage fragment as reported previously (Weng et al., 2005), which was detectable after 24-h exposure to LAQ824 (Fig. 6A). Thereafter, U937 cells stably expressing either the empty vector (U937/pCEP) or Mcl-1 (U937/Mcl-1) cDNAs were exposed to LAQ824 75 nM for 24 to 48 h. As shown in Fig. 6B, U937/Mcl-1 cells displayed a significant reduction in the extent of apoptosis induced by 75 nM LAQ824 (Fig. 6B), which was also reflected by reduced activation of caspases-3, -7, and -9 and PARP degradation (Fig. 6C). Together, these results suggest that down-regulation of both antiapoptotic proteins through either transcriptional (e.g., XIAP) or primarily caspase-dependent mechanisms (e.g., Mcl-1) plays a significant role in the disparate apoptotic responses of U937 cells to low (30 nM) versus high (75 nM) LAQ824 concentrations.
Exposure of U937 Cells to LAQ824 Induces a Dramatic Increase in the Level of the Proapoptotic Lipid Second-Messenger Ceramide. Because ceramide has been implicated in apoptosis induction in human leukemia cells (Jarvis et al., 1994; Kurita-Ochiai et al., 2003), U937 and Jurkat cells were treated with LAQ824 (U937, 30 and 75 nM; Jurkat, 50 nM) for various intervals, after which ceramide levels were monitored. As shown in Fig. 7A, lethal concentrations of LAQ824 induced significant increases in ceramide levels in both U937 and Jurkat cells, which were ∼5-fold higher than controls at 24 h and ∼15-fold higher after 48 h. In contrast, 30 nM LAQ824 only minimally increased ceramide levels (Fig. 7A), raising the possibility that differential increases in ceramide levels may also contribute to the disparate effects of low versus high drug concentrations.
Generation of ceramide may proceed from either de novo synthesis or hydrolysis of sphingomyelin (Pettus et al., 2002). Consequently, U937 cells were coincubated with inhibitors targeting these pathways at different points. The de novo synthesis inhibitors fumonisin B1 and myriocin exerted no effects on LAQ824-induced apoptosis. Likewise, the neutral-sphingomyelinase inhibitor GW4869 was also ineffective. In contrast, desipramine, an inhibitor of acid sphingomyelinase (Pettus et al., 2002), partially but significantly blocked LAQ824-mediated apoptosis (i.e., LAQ824 75 nM, 82.3 ± 2.1 versus 75 nM LAQ824 plus desipramine, 52.6 ± 2.2 at 48 h; P < 0.01; Fig. 7B), suggesting that LAQ824-induced ceramide generation through the hydrolysis of sphingomyelin contributes functionally to the lethal effects of high concentrations of LAQ824. Finally, blockade of LAQ824-induced ROS by NAC essentially abrogated ceramide generation (Fig. 7C), indicating that LAQ824-induced ROS generation leads to the acid sphingomyelinase generation of ceramide and that the latter event represents an important functional mediator of LAQ824-induced lethality.

Mcl-1 degradation contributes to LAQ824-induced lethality. A, U937 cells were exposed to either 30 or 75 nM LAQ824 for the indicated intervals, after which cells were collected, lysed, and analyzed by Western blot. In each case, 30 μg of protein was separated by SDS-PAGE, blotted, and probed with the corresponding antibodies. The results of a representative study are shown; two additional experiments yielded similar results. B, both U937 cells overexpressing a cDNA encoding for Mcl-1 (U937/Mcl-1) or the corresponding empty vector (U937/pCEP) cells were exposed for 24 and 48 h to 75 nM LAQ824, after which the percentage of apoptotic cells (annexinV/PI+) was determined by flow cytometry as described under Materials and Methods. Values represent the means ± S.D. for three separate experiments performed in triplicate. C, Western blot analysis of lysates from U937/pCEP or U937/Mcl-1 cells; cells were exposed to 75 nM LAQ824 for the indicated intervals, after which they were collected, lysed, and 30 μg of protein was separated by SDS-PAGE, blotted, and probed with the corresponding antibodies. The results of a representative study are shown; two additional experiments yielded similar results.
Discussion
Whereas previous studies have demonstrated that HDACIs can induce differentiation and/or apoptosis in human leukemia cells (Vrana et al., 1999; Rosato et al., 2001, 2003b), the factors determining which of these alternative cell fates predominates remain largely unknown. In the case of LAQ824, the present results demonstrate that low concentrations of the drug led to cell-cycle arrest at G0/G1, followed by cellular maturation, whereas higher concentrations induced G2/M arrest accompanied by pronounced mitochondrial injury and apoptosis. These findings are consistent with previous reports of HDACI-mediated tumor cell G2M arrest and subsequent death in mitosis caused by an aberrant G2 checkpoint (Qiu et al., 2000). Furthermore, both induction of p21WAF1/CIP1 and generation of ROS represented a common response to low and high concentrations of LAQ824, although the latter phenomenon occurred earlier in cells exposed to lethal LAQ824 concentrations. However, in addition to increased ROS generation, LAQ824-induced apoptosis was accompanied by pronounced down-regulation of XIAP (at the transcriptional level) and Mcl-1 (through post-transcriptional modifications) as well as the marked acid sphingomyelinase-dependent generation of ceramide. Although differentiation and apoptosis share certain features (e.g., induction of mitochondrial injury), there is evidence that in leukemia cells, these events are mutually exclusive (Selvakumaran et al., 1994). It is therefore likely that when administered above threshold levels, LAQ824 triggers multiple events that culminate in apoptosis and prevent engagement of a differentiation program. It is important to note that whereas differentiation-induction by low LAQ824 concentrations resulted in a marked loss of clonogenic potential, high concentrations of LAQ824 were significantly more inhibitory, reflecting the pronounced increase in apoptosis.

Exposure of U937 cells to LAQ824 results in the acid sphingomyelinase-dependent increase in ceramide levels. A, both U937 and Jurkat cells were exposed to 30 to 75 nM and 50 nM LAQ824, respectively, for the indicated intervals, after which ceramide levels, normalized to levels of total phospholipids, were compared with those of untreated control cells. Values are expressed as fold increases over control levels and represent the means for three separate determinations ± S.D. *, significantly greater than values for U937 cells exposed to 30 nM LAQ824, P < 0.01. B, U937 cells were incubated for either 24 or 48 h with LAQ824 in the absence (L75) or the presence of various inhibitors of ceramide synthesis (FU, 25 μM fumonisin B1; MY, 10 μM myriocin; GW, 10 μM GW4869; DE, 30 μM desipramine) after which apoptosis (annexinV/PI positivity) was monitored as described under Materials and Methods. *, significantly lower than values for U937 cells exposed to 75 nM LAQ824; P < 0.05. C, U937 cells were coexposed to 75 nM LAQ824 in the presence or absence (L75) of the free radical scavenger NAC or the acid sphingomyelinase inhibitor desipramine (Des) for 24 or 48 h, after which ceramide levels were determined as described under Materials and Methods. Values represent the means ± S.D. for three separate experiments performed in triplicate. *, significantly lower than values for U937 cells exposed to 75 nM LAQ824; P < 0.05.
Recent studies have highlighted the role that the extrinsic apoptotic pathway plays in HDACI-mediated lethality, including up-regulation of TRAIL and TRAIL-death receptors DR4 and DR5 in several human leukemia cell lines (Guo et al., 2004; Insinga et al., 2005; Nebbioso et al., 2005). However, activation of the extrinsic/receptor-mediated pathway did not play a significant role in LAQ824-mediated lethality in U937 cells, and in contrast to Jurkat cells, death receptor expression was not up-regulated by LAQ824, suggesting that the latter phenomenon is cell lineage-specific. Furthermore, the finding that Bcl-2 or Bcl-xL overexpression, by blocking the release of cytochrome c, AIF, and Smac, substantially protected cells against LAQ824 lethality, strongly implicates the mitochondrial pathway (Donovan and Cotter, 2004) in LAQ824 lethality. In this regard, LAQ824-induced degradation of several Bcl-2 family proteins including Bcl-2, Bcl-xL, and Mcl-1 which, when overexpressed, significantly protected cells from lethality, support the notion that their down-regulation, although largely secondary events, nevertheless contribute substantially to amplify the cell death process.
Although the mechanisms by which HDACIs induce cell death may vary considerably (Rosato and Grant, 2004), induction of oxidative injury represents a common theme. It is noteworthy that ROS generation was observed in both U937 and Jurkat cells exposed to lethal concentrations of LAQ824. It is noteworthy that ROS production as well as all downstream events, including mitochondrial injury, caspase activation, and apoptosis, were blocked by the free radical scavenger NAC, indicating that oxidative injury represents a primary event in activation of the apoptotic cascade. It is also noteworthy that prevention of ROS generation by NAC blocked LAQ824-mediated induction of ceramide. This finding is consistent with evidence that ceramide generation represents an important downstream signaling event in cells experiencing oxidative damage and mitochondrial injury (Mansat-de Mas et al., 1999). Both low and high LAQ824 concentrations elicited unexpectedly similar increases in ROS levels, although these occurred earlier in the case of 75 nM LAQ824. The functional significance of the early induction of oxidative injury by high LAQ824 concentrations remains to be determined. The present results contrast with those involving the benzamide HDACI MS-275, in which leukemic cell apoptosis correlated closely with ROS generation (Rosato et al., 2003b). This suggests that the functional significance of HDACI-induced ROS production in human leukemia cells may vary with the class of HDACI involved. As a whole, these findings indicate that LAQ824-mediated oxidative damage is necessary but not sufficient to trigger apoptosis in these cells and that for this to occur, other events (e.g., XIAP/Mcl-1 down-regulation, ceramide generation, etc.) must supervene.
Studies performed using U937 cells ectopically expressing either XIAP or Mcl-1 suggest that down-regulation of these proteins plays a significant functional role in LAQ824-induced lethality. For example, a clear correlation between LAQ824-induced XIAP down-regulation and activation of target caspases-9, -3, and -7 was observed; conversely, enforced expression of XIAP blocked activation of these caspases. XIAP down-regulation has been reported previously in human leukemia cells exposed to HDACIs, notably MS-275 (Rosato et al., 2003b), although the molecular mechanisms responsible for this phenomenon have not been explored in detail. The present results suggest that LAQ824-mediated down-regulation of XIAP levels primarily involves transcriptional repression, a phenomenon amplified by caspase-mediated degradation. It is well known that HDAC inhibitors, by modulating histone acetylation and remodeling chromatin architecture, regulate gene expression, which can involve both up- and down-regulation (Peart et al., 2005). Furthermore, two major mechanisms have been implicated in XIAP degradation: ubiquitinization and proteasomal degradation (Yang et al., 2000) and/or caspase degradation (Johnson et al., 2000). The finding that the antioxidant NAC partially blocked XIAP down-regulation by LAQ824 in association with diminished apoptosis suggests that caspase-dependent cleavage contributes to this phenomenon, although the possibility of enhanced proteasomal degradation cannot be excluded. On the other hand, results of real-time RT-PCR studies strongly suggest that transcriptional repression plays an important role in this phenomenon. Finally, the observation that only higher LAQ824 concentrations resulted in transcriptional repression and down-regulation of XIAP supports the notion that differential effects on XIAP contribute to disparate dose-responsive effects of LAQ824 in these cells. In contrast to XIAP, Mcl-1 down-regulation induced by higher LAQ824 concentrations primarily represented a secondary, caspase-mediated process, although in all likelihood, it contributed to lethality. The importance of Mcl-1 in circumventing apoptosis in malignant hematopoietic cells has been documented (Moulding et al., 2000). Furthermore, caspase 3-mediated Mcl-1 degradation promotes apoptosis by two mechanisms (i.e., by reducing Mcl-1 expression levels and by generating an Mcl-1 C-terminal fragment with proapoptotic activity) (Weng et al., 2005). The C-terminal portion of Mcl-1 promotes cytochrome c release from the mitochondria by forming a heterodimer with tBid (Weng et al., 2005). Similar observations have been reported in the case of Bcl-2 (Kirsch et al., 1999), which also experienced cleavage in cells exposed to high concentrations of LAQ824. Thus, although these phenomena represent secondary events in LAQ824-induced apoptosis, both degradation and generation of proapoptotic Mcl-1 and Bcl-2 fragments may amplify the caspase cascade and contribute to the time-dependent increase in lethality.
Another factor implicated in the lethality of high LAQ824 concentrations is ceramide, a lipid second messenger involved in cellular responses to stress and apoptosis induction (Pettus et al., 2002), including that generated by HDACIs such as butyrate and MS-275 (Kurita-Ochiai et al., 2003; Maggio et al., 2004). It is noteworthy that only high concentrations of LAQ824 triggered pronounced increases in ceramide. Furthermore, inhibition of ceramide generation via acid sphingomyelinase attenuated LAQ824-mediated lethality, arguing that increased ceramide generation plays a functional role in LAQ824-induced apoptosis. Acid sphingomyelinase is activated by several proapoptotic stimuli, including induction of death receptors (i.e., CD95/FAS and tumor necrosis factor-R) (Gulbins and Grassme, 2002) and γ and UV radiation (Zhang et al., 2001). It is noteworthy that LAQ824-mediated ceramide generation was dependent on early ROS generation. A close relationship between ROS and ceramide production has been characterized in several models, including U937 cells exposed to daunorubicin and rat primary astrocytes (Mansat-de Mas et al., 1999). It is unclear at present why increases in ROS induced by low LAQ824 concentrations failed to increase ceramide levels but higher LAQ824 concentrations did increase ceramide. One speculative possibility is that high concentrations of LAQ824 might interfere with the function of chaperones such as the 90-kDa heat shock protein (Atadja et al., 2004b) and thus enhance the degradation of proteins that inhibit ceramide generation. On the other hand, LAQ824 might lead to down-regulation of as-yet-to-be-determined genes with similar functions. In any case, the present studies suggest that multiple factors cooperate to disrupt leukemic cell differentiation and promote apoptosis by suprathreshold concentrations of LAQ824. Furthermore, these findings may have implications for the rational development of combination regimens involving this novel HDACI in hematological malignancies.
Footnotes
- Received July 22, 2005.
- Accepted September 27, 2005.
This work was supported by awards CA63753, CA93738, CA100866 (to S.G.), and CA61774 (to S.S.) from the National Cancer Institute, award 6045-03 from the Leukemia and Lymphoma Society of America, an award from the V Foundation, and Department of Defense awards DAMD-17-03-1-0209 (to S.G.) and DAMD-17-02-1-0060 (to S.S.).
Article, publication date, and citation information can be found at http://molpharm.aspetjournals.org.
doi:10.1124/mol.105.017145.
ABBREVIATIONS: HDACI, histone deacetylase inhibitor; ROS, reactive oxygen species; PI, propidium iodide; PARP, poly(ADP-ribose)polymerase; AIF, apoptosis-inducing factor; ΔΨm, mitochondrial membrane potential; PAGE, polyacrylamide gel electrophoresis; PBS, phosphate-buffered saline; NAC, N-acetyl cysteine; HDAC, histone deacetylase; RT-PCR, reverse transcription-polymerase chain reaction; pRb, retinoblastoma protein; AN-9, pivaloyloxymethyl butyrate; MS-275, 3-pyridinylmethyl ((4-(((2-aminophenyl)amino)carbonyl)phenyl)methyl)carbamate; CI-994, 4-(acetylamino)-N-(2-aminophenyl)benzamide; NVP-LAQ824, (2E)-N-hydroxy-3-[4-[[(2-hydroxyethyl)[2-(1H-indol-3-yl)ethyl]amino]methyl]-phenyl]-2-propenamide; GW4869, N,N′-bis[4-(4,5-dihydro-1H-imidazol-2-yl)phenyl]-3,3′-p-phenylene-bis-acrylamide dihydrochloride.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}