


Abstract
The copper efflux transporters ATP7A and ATP7B sequester intracellular copper into the vesicular secretory pathway for export from the cell. The influence of these transporters on the pharmacodynamics of cisplatin, carboplatin, and oxaliplatin was investigated using human Menkes' disease fibroblasts (Me32a) that do not express either transporter and sublines molecularly engineered to express either ATP7A (MeMNK) or ATP7B (MeWND). Cellular copper levels were significantly higher in the Me32a cells than in the MeMNK and MeWND sublines. These transporter-proficient sublines were resistant to the cytotoxic effect of copper, cisplatin, and carboplatin but were hypersensitive to oxaliplatin. Whole-cell accumulation of platinum after a 24-h exposure was significantly increased in the MeMNK and MeWND cells for all three platinum drugs, but this was accompanied by an increase in the amount of platinum reaching the DNA only for oxaliplatin. Vesicles isolated from MeMNK cells contained more platinum after exposure to cisplatin and carboplatin, whereas the platinum content of vesicles from MeWND cells was increased after exposure to all three drugs. Although copper triggered relocalization of ATP7A from the perinuclear region to more peripheral locations, the platinum drugs did not. These results demonstrate that both ATP7A and ATP7B modulate the pharmacodynamics of all three clinically used platinum drugs. The data are consistent with the hypothesis that these copper exporters sequester the platinum drugs into subcellular compartments, limiting their cytotoxicity, similar to their effect on copper. However, in this model system, although copper is readily exported after vesicular sequestration, the platinum drugs are not.
Cisplatin (DDP) and carboplatin (CBDCA) are widely used in the treatment of human cancers, and oxaliplatin (l-OHP) has recently been marketed for the treatment of colon cancer. However, resistance to all of these agents emerges commonly during therapy and is a major cause of treatment failure. The mechanisms underlying clinical resistance to the platinum-containing drugs are poorly characterized. Past studies have noted reduced cellular drug accumulation, increased cytoplasmic detoxification, and increased DNA repair in resistant cells (Siddik, 2003). The cytotoxicity of DDP and its analogs is a function of how much drug enters the cell and how much avoids intrinsic detoxification mechanisms and successfully reaches the nucleus to react with DNA (Andrews and Howell, 1990). The mechanisms involved in transporting DDP across the plasma membrane and through the cytoplasm to the nucleus are not well-defined. DDP, CBDCA, and l-OHP are all relatively polar molecules that do not cross lipid bilayer membranes easily. Previous studies have suggested that one or more transporters are involved in DDP uptake (Andrews and Albright, 1991).
ATP7A and ATP7B are copper transporters that sequester copper from the cytoplasm into the TGN for loading onto ceruloplasmin and other copper-requiring enzymes and subsequent export from the cell. ATP7A is expressed in many tissues, whereas the expression of ATP7B is limited to liver. Mutations that disable these exporters cause human disease. Loss of ATP7A function results in Menkes' disease, and loss of ATP7B function produces Wilson's disease. In both diseases, copper accumulation and distribution among tissues is perturbed (Suzuki and Gitlin, 1999). When copper levels are low, ATP7A and ATP7B are localized to the TGN, in which they function to incorporate the metal into copper-dependent enzymes. When copper levels are increased, a substantial amount of ATP7A relocalizes from the TGN to the plasma membrane, whereas ATP7B relocates primarily to intracellular vesicular compartments (Pena et al., 1999).
Recent studies from our laboratory have demonstrated that in vitro selection of human ovarian carcinoma cells for DDP resistance results in cross-resistance to copper and vice versa (Katano et al., 2002; Safaei et al., 2003). ATP7A was found to be overexpressed at the protein level in some DDP-resistant ovarian carcinoma cell lines (Katano et al., 2002). The concept that copper exporters might mediate DDP resistance was introduced by Komatsu et al. (2000), who reported DDP resistance in prostate carcinoma cells overexpressing ATP7B. Subsequent studies have reported that overexpression of ATP7B is found in a variety of DDP-resistant tumors (Nakayama et al., 2001, 2002; Kanzaki et al., 2002; Higashimoto et al., 2003; Miyashita et al., 2003; Ohbu et al., 2003). In addition, studies in yeast have demonstrated a role for the copper-uptake transporter CTR1 in mediating both the cellular pharmacology of DDP and its ability to kill cells (Ishida et al., 2002; Lin et al., 2002).
We have examined the effect of ATP7A and ATP7B on the cellular pharmacology of DDP, CBDCA, and l-OHP and on their cytotoxicity in a well-characterized isogenic panel of human fibroblasts molecularly engineered to express these proteins. The studies were performed using the human fibroblast cell line Me32a, established from a patient with Menkes' disease that does not express either ATP7A or ATP7B, and Me32a sublines transfected with an ATP7A or ATP7B expression vector (La Fontaine et al., 1998). We report here that both copper exporters regulate sensitivity to the cytotoxic effects of the three clinically used platinum-containing drugs and that these changes in sensitivity are accompanied by substantial alterations in their cellular pharmacology and intracellular trafficking.
Materials and Methods
Drugs. DDP and CBDCA were generously provided by Bristol-Myers Squibb Co. (Stamford, CT), and l-OHP was provided by SANOFI Research Center (Montpellier, France). DDP was stored as 1 mM stock solution in 0.9% NaCl in the dark at room temperature, CBDCA was stored as 27 mM stock solution in water at 4°C, and l-OHP was stored as 12.6 mM stock solution in water at -20°C. Cupric sulfate was obtained from Fisher Scientific Co. (Tustin, CA).
Cell Lines. The ATP7A- and ATP7B-nonexpressing fibroblast cell line Me32a, the isogenic ATP7A-transfected cell line MeMNK, and the isogenic ATP7B-transfected cell line MeWND were generously provided by Michael J. Petris (Columbia, MO) and were established as described previously (La Fontaine et al., 1998). For growth-enrichment studies, Me32a cells were transfected with a vector expressing EYFP, and MeMNK and MeWND cells were transfected with a vector expressing ECFP (BD Biosciences Clontech, Palo Alto, CA). Clones that expressed these fluorescence proteins at high levels were recovered by flow cytometry and were used in subsequent experiments. All cell lines were maintained at 37°C in a humidified incubator containing 5% CO2 in α-minimal essential medium supplemented with 10% fetal bovine serum (Invitrogen, Carlsbad, CA). MeMNK and MeWND cells and fluorescent protein-transfected Me32a, MeMNK, and MeWND cells were grown in the presence of 500 μg/ml G418.
Measurement of Drug Sensitivity. Me32a-EYFP, MeMNKECFP, and MeWND-ECFP cells were separately plated in 96-well plates at a density of 5000 cells/200 μl/well and grown overnight in α-minimal essential medium without phenol red (Invitrogen). drug (50 μl) diluted in medium from the stock concentration identified above was added to each well to produce the desired final concentration in a total of 250 μl, and the fluorescence intensity was measured immediately (t = 0) as well as after 72 h (t = 72) using a Safire microplate reader (Tecan, Zurich, Switzerland). Excitation/emission wavelengths were set at 434/476 and 518/535 nm for ECFP and EYFP, respectively. Because Me32a-EYFP, MeMNK-ECFP, and MeWND-ECFP cells grew at different rates, the growth rate in the presence of drug was expressed as a fraction of that in the absence of drug for each concentration tested.
Copper Accumulation. Cells were grown to ∼80% confluence in 100-mm dishes. For ICP-OES measurements, five plates were used for each data point, and the cells were incubated in fresh medium containing 0 to 100 μM Cu2+ for 24 h. The cells were then washed three times with cold PBS (4°C), scraped, and pelleted. The pellets were dried, resuspended in 70% nitric acid, and dissolved at 65°C for 2 h, after which the samples were diluted with water to a final concentration of 5% acid. Measurements were made using a PerkinElmer ICP-OES (model 3000DV; PerkinElmer Life and Analytical Sciences, Boston, MA) from the Analytical Facility at the Scripps Institute of Oceanography and normalized to protein levels for each cell line.
DDP Whole-Cell Uptake and Accumulation in DNA. Cells were grown to 80% confluence in either six-well plates (whole-cell uptake) or 145-mm plates (DNA accumulation). For whole-cell uptake studies, three wells were used for each data point, and the cells were incubated in fresh medium containing DDP for 24 h. The cells were then washed with PBS and were lysed directly by the addition of 215 μl of 70% nitric acid into each well. The cells were then collected and dissolved at 65°C for 2 h, after which the samples were diluted with water to a final concentration of 5% acid. For DNA platinum accumulation studies, three plates were used for each data point, and the cells were incubated in fresh medium containing DDP for 24 h. The cells were then washed with PBS, scraped, and pelleted before isolation of genomic DNA using a Wizard Genomic DNA Purification Kit (Promega, Madison, WI). Harvested DNA was originally resuspended in a small volume of water for quantification. After quantification, 215 μl of 70% nitric acid was added to the DNA, and the samples were dissolved at 65°C for 2 h, after which the samples were diluted with water to a final concentration of 5% acid. Measurements were made using a Thermo Finnigan ICP-MS (model Element2; Thermo Finnigan, San Jose, CA) from the Analytical Facility at the Scripps Institute of Oceanography and normalized to protein levels or DNA amounts for each cell line. Indium was added to each sample at 1 part per billion as a control for flow variation.
Synthesis of fDDP. fDDP was prepared using a modification of the method reported by Molennar et al. (2000). Briefly N3-tBoc-1,2,3-propanetriamine was prepared from 3-amino-1,2-propanediol and then reacted with potassium tetrachloroplatinate to obtain [1-(tBocaminomethyl)-1,2-ehtylenediamine] dichloroplatinum(II). The tBoc group was removed with 0.1 N HCl at 70°C, and the resulting platinated salt was neutralized with 2 N NaOH and then reacted with 5(6)-carboxyfluorescein preactivated by treatment with 1-ethyl-2-(2-dimethylaminopropyl)carbodiimide and N-hydroxysuccinimide in dimethyl formamide. After centrifugation and successively washing with water, ethanol, and ether, the final product was characterized for DDP and fluorescein content by 1H NMR and mass spectrometry.
Assessment of Accumulation of fDDP in Whole Cells by Flow Cytometry. Me32a, MeMNK, and MeWND cells were plated in T-75 flasks and grown to ∼80% confluence. Cells were then treated with medium alone (to account for autofluorescence) or fDDP for 2 h. Treated and untreated cells were harvested in trypsin, and the cell pellets were resuspended in 1 ml of 5% bovine serum albumin in PBS. A FACSCalibur analyzer with CellQuest acquisition software (BD Biosciences Immunocytometry Systems, San Jose, CA) was used to collect and analyze data on at least 24,000 events per sample. Events were gated by forward and side scatter to exclude aggregates and debris, and the fluorescence intensity excited by the 488-nm laser was measured in the FL1 channel (530 ± 15 nm).
Vesicle Preparation. Cells were plated in 145-mm plates and grown to ∼80% confluence. All subsequent steps were performed with ice-cold solutions, and all spins were performed at 4°C. The cells were rinsed once with PBS and then rinsed twice with 1 mM sodium bicarbonate solution. Each plate was then scraped with 1 mM sodium bicarbonate solution with added Complete Mini, EDTA-free protease inhibitor cocktail tablets (Roche Diagnostics, Mannheim, Germany) into a 15-ml conical tube (three plates were combined for each data point), and the cells were homogenized for 5 min. The lysate was spun at 3000g for 5 min, and the postnuclear fraction was recovered and added to a 2-ml cushion of 38% sucrose in 100 mM sodium carbonate. The samples were spun in an ultracentrifuge at 25,000 rpm for 35 min, and the vesicular band was carefully recovered and transferred to a new ultracentrifuge tube. The tubes were filled with a 50 mM Tris solution containing 250 mM sucrose with added Complete Mini, EDTA-free protease inhibitor cocktail tablets. The samples were again spun in an ultracentrifuge at 26,000 rpm for 45 min to pellet the vesicles. Each pellet was resuspended in 100 μl of standard radioimmunoprecipitation assay buffer (150 mM NaCl, 1% Nonidet P-40, 0.5% deoxycholate, 0.1% SDS, and 50 mM Tris, pH 7.5) and then prepared for ICP-MS analysis as described above. For quantitative purposes, each sample was also analyzed for sulfur content using ICP-OES as described above, and sulfur levels were used to normalize total platinum accumulation.
Digital Microscopy. Cells were cultured for 3 to 4 days in non-selective media on round coverslips (Fisherbrand 12-545-81, 12CIR.-1.5) in 24-well plates and treated for 1 h at 37°C with drug in Opti-MEM media (Invitrogen). All subsequent steps were performed at room temperature unless otherwise indicated. After three rinses in PBS, the cells were fixed for 30 min in 3.7% formaldehyde in PBS, rinsed twice with PBS, permeabilized for 15 min in 0.3% Triton X-100 in PBS, and rinsed twice with 0.1% Tween in PBS. Cells were then blocked for 1 h in PBS containing 1% bovine serum albumin and 0.1% Tween 20, followed by incubation with a 1:50 dilution of anti-MNK (Clone 34; BD Biosciences, San Jose, CA) for 2 h at 37°C. Cells were then rinsed three times with PBS containing 0.1% Tween and incubated with 1:2000 dilution of Hoechst 33342 (Molecular Probes) and Alexa 594 anti-mouse (#A21125; Molecular Probes) at 1:1000 dilution for 2 h at 37°C. Cells were then rinsed three times with PBS and twice with water before mounting on glass slides with gelvatol. Slides were kept in the dark at room temperature. Digital microscopic examination was performed at University of California San Diego Cancer Center Digital Imaging Shared Resource using a DeltaVision deconvoluting microscope system (Applied Precision, Inc., Issaquah, WA). Images were captured by 100×, 60×, and 40× lenses using SoftWorx software (Applied Precision) on a Silicon Graphics Octane workstation (SGI, Mountain View, CA).
Statistics. All comparisons were made using Student's t test with the assumption of unequal variance.
Results
Basal Copper Content and Accumulation. These studies were conducted using Me32a human fibroblasts established from a patient with Menkes' disease that expressed no detectable levels of either ATP7A or ATP7B, and sublines transfected with vectors expressing one or the other of these proteins (MeMNK and MeWND cells, respectively). These cell lines have been characterized previously with respect to the effect of ATP7A and ATP7B expression on copper transport (La Fontaine et al., 1998), and differences in copper homeostasis between these lines were further documented as part of the studies reported here. First, basal copper levels were measured in cells grown in standard medium to which no additional copper had been added. Under these conditions, the Me32a cells contained 56.7 ± 4.1 ng of Cu2+/mg of protein (S.E.M.), whereas the MeMNK cells contained only 19.2% of this amount (10.9 ± 0.9 ng of Cu2+/mg of protein, S.E.M.; p < 0.0003). The basal copper level in the MeWND cells was only 23.7% of that in the Me32a cells (13.5 ± 1.7 ng of Cu2+/mg of protein, S.E.M.; p < 0.0002). Copper levels were also measured in the Me32a, MeMNK, and MeWND cells after a 24-h exposure to increasing concentrations of CuSO4. As shown in Fig. 1, copper levels were concentration-dependent in all three lines. However, the exporter-deficient Me32a cells contained significantly more copper at all copper concentrations tested (p < 0.002 for all concentrations). These results were confirmed in studies in which the cells were exposed to 2 μM 64Cu for either 1 or 24 h (data not shown). Thus, the members of this panel of cell lines have well-defined differences in the cellular pharmacology of copper that are directly attributable to differences in the expression of ATP7A and ATP7B.

Copper content in whole cells after a 24-h exposure to CuSO4 as a function of copper concentration. ▪, Me32a cells; □, MeMNK cells;  , MeWND cells. Each data point represents the mean of a single experiment performed with 10 cultures per data point. Vertical bars, ±S.E.M. (in the MeMNK and MeWND data points, the error bar is contained within the symbols).
, MeWND cells. Each data point represents the mean of a single experiment performed with 10 cultures per data point. Vertical bars, ±S.E.M. (in the MeMNK and MeWND data points, the error bar is contained within the symbols).
Sensitivity to the Growth-Inhibitory Effects of Copper, DDP, CBDCA, and l-OHP. Me32a cells and its sublines do not form well-separated independent colonies when seeded sparsely on plastic; therefore, sensitivity to the cytotoxic effect of copper and the platinum drugs was assessed by measuring change in growth rate rather than by using clonogenic assays. Each cell line was rendered fluorescent by stable transfection with a vector expressing either EYFP or ECFP, and growth rates were measured during a period of 72 h of drug exposure by quantifying total fluorescence in the culture. The data presented in Fig. 2 and Table 1 show that the MeMNK and MeWND cells were significantly more resistant to copper, DDP, and CBDCA than the Me32a cells. It is interesting that the MeMNK and MeWND cells exhibited hypersensitivity rather than resistance to l-OHP. Thus, under circumstances in which ATP7A and ATP7B expression mediated reduced sensitivity to copper, DDP, and CBDCA, both exporters actually increased sensitivity to l-OHP. This is of particular interest given that l-OHP exhibits a different profile of clinical activity than DDP and CBDCA (Rixe et al., 1996; Raymond et al., 1998).
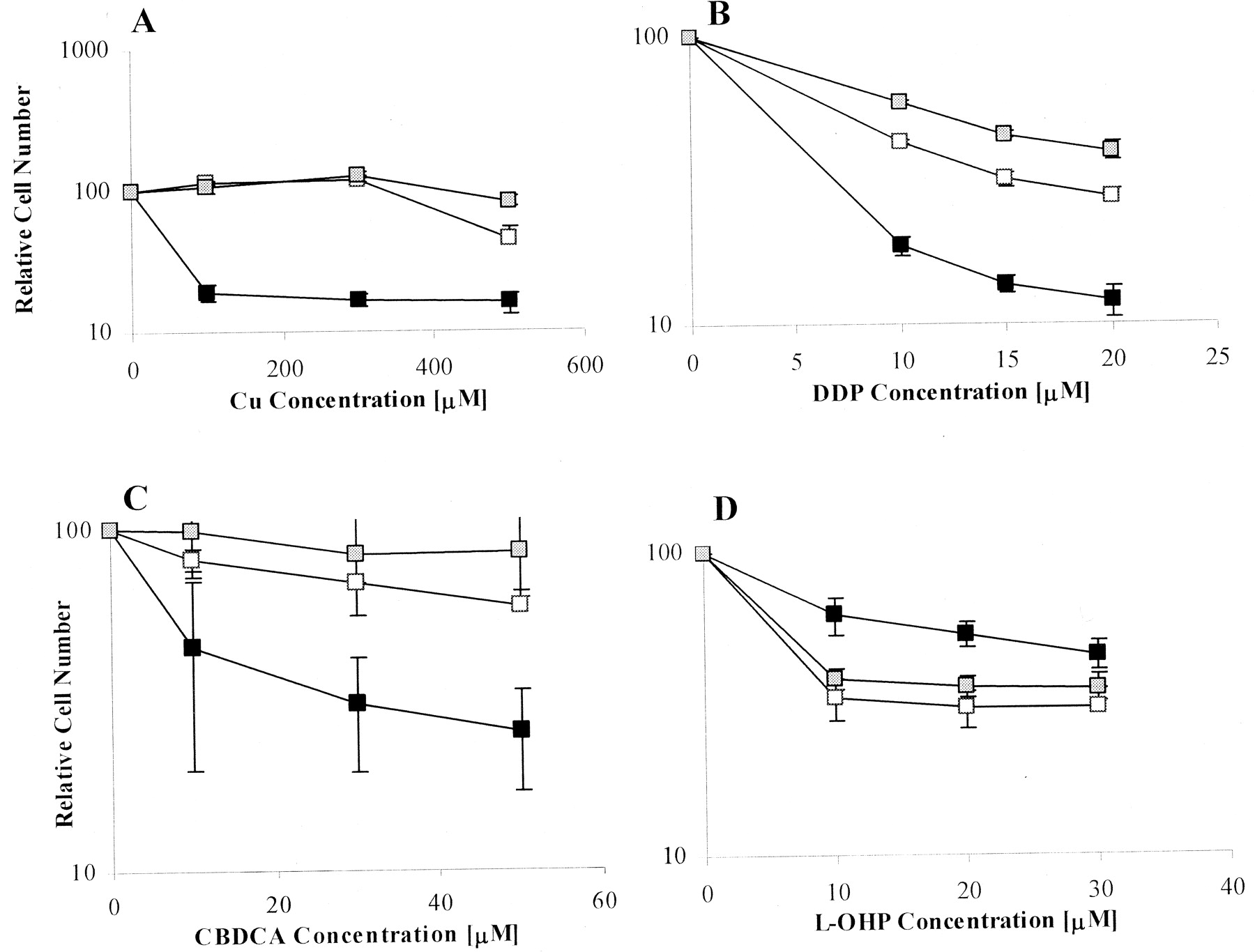
Sensitivity to the growth-inhibitory effects of CuSO4 (A), DDP (B), CBDCA (C), or l-OHP (D) as a function of drug concentration. ▪, Me32a cells; □, MeMNK cells;  , MeWND cells. Each data point represents the mean of three independent experiments, each performed with triplicate cultures. Vertical bars, ±S.E.M.
, MeWND cells. Each data point represents the mean of three independent experiments, each performed with triplicate cultures. Vertical bars, ±S.E.M.
IC50 values for growth inhibition
DDP Whole-Cell Accumulation. Experiments were conducted to determine whether the changes in sensitivity to the platinum drugs were accompanied by altered net whole-cell platinum accumulation after a 24-h drug exposure. Each cell line was tested at several drug concentrations spanning the levels found in the plasma of patients undergoing treatment. As shown by the data presented in Fig. 3, A to C, the MeMNK and MeWND cells accumulated higher levels of platinum at all three drug concentrations tested for each of the three platinum drugs (p < 0.02 for all comparisons relative to accumulation in Me32a cells). Thus, despite the fact that the MeMNK and MeWND cells were clearly less sensitive to the growth-inhibitory effects of DDP and CBDCA, they accumulated significantly more platinum during a relatively long period of exposure. This result suggests the possibility that, rather than functioning to export DDP and CBDCA from the cell, these copper transporters serve to sequester them from the cytoplasm into vesicular structures, thus effectively detoxifying the drugs while at the same time increasing total cell accumulation. It is noteworthy that apparently l-OHP is also subject to this mechanism, because MeMNK and MeWND cells exposed to this drug also accumulated more total cellular platinum than did the Me32a cells. However, in the case of l-OHP, the drug was not as effectively detoxified.

Platinum content in the whole cell after a 24-h exposure as a function of DDP (A), CBDCA (B), or l-OHP (C) concentration. ▪, Me32a cells; □, MeMNK cells;  , MeWND cells. Each bar represents the mean of a single experiment performed with triplicate cultures. Vertical bars, ±S.E.M.
, MeWND cells. Each bar represents the mean of a single experiment performed with triplicate cultures. Vertical bars, ±S.E.M.
Accumulation of fDDP in Whole Cells. To confirm the increased accumulation of DDP in the MeMNK and MeWND cells compared with Me32a cells, the cell lines were treated with a fluorescein-tagged form of DDP, fDDP. After a 2-h treatment with either medium alone or 2 μM fDDP, cells were harvested and intracellular fluorescein-specific fluorescence levels were measured by flow cytometry. As shown in Table 2, the MeMNK and MeWND cells accumulated significantly higher fluorescent intensities after treatment with fDDP compared with the Me32a cells (p < 0.003 for MeMNK cells and p < 0.0002 for MeWND cells), indicating greater accumulation of fDDP in these cells. These results provide further evidence that the expression of ATP7A and ATP7B in the MeMNK and MeWND cells is associated with increased accumulation of DDP.
Whole-cell accumulation of fDDP after a 2-h exposure to 2 μM drug
Values are presented as the mean ± S.E.M. of three independent experiments.
Platinum Accumulation in DNA. The platinum drugs are believed to injure cells primarily through the formation of DNA adducts. The extent of adduct formation, as reflected by the amount of platinum in DNA, was measured after a 24-h exposure to 10 μM DDP, 50 μM CBDCA, and 15 μM l-OHP, concentrations that yielded clear differences in the extent of whole-cell platinum accumulation. As shown in the top of Fig. 4, no differences were detectable in the amount of platinum in the DNA between any of the three cell lines exposed to DDP or CBDCA. Thus, for DDP and CBDCA-treated MeMNK and MeWND cells, the increased whole-cell accumulation of drug was not accompanied by more extensive adduct formation, further suggesting that the effect of ATP7A and ATP7B expression in these fibroblasts was to mediate the accumulation of the platinum drugs in some compartment of the cell in which it did not exert cytotoxicity. It is noteworthy that the MeMNK and MeWND cells did exhibit increased platinum levels in DNA after treatment with l-OHP (p < 0.01 and p < 0.02, respectively), which is consistent with the greater sensitivity of the MeMNK and MeWND cells to the cytotoxic effect of l-OHP than with Me32a cells, a feature that again distinguishes l-OHP from DDP and CBDCA.

Ratio of the accumulation in platinum DNA (top) and vesicles (bottom) isolated from MeMNK and MeWND relative to that in Me32a cells after exposure to 10 μM DDP, 50 μM CBDCA, or 15 μM l-OHP. Each bar represents the mean ratio of accumulation in the MeMNK cells (□), or MeWND cells ( ) to that in the Me32a cells as determined in two to four independent experiments; vertical bars, ± S.E.M.
) to that in the Me32a cells as determined in two to four independent experiments; vertical bars, ± S.E.M.
Platinum Accumulation in Vesicular Compartments. The results of the whole-cell and DNA platinum-accumulation studies suggested that, as has been well-established for copper, ATP7A and ATP7B act to sequester DDP and CBDCA into one or more subcellular vesicular compartments and thus limit their access to DNA and their cytotoxicity. To directly demonstrate increased vesicular sequestration as a consequence of ATP7A and ATP7B expression, the ratio of the platinum content of the vesicular fraction from the MeMNK and MeWND cells to that in the Me32a cells was determined by ICP-MS after exposure to equitoxic concentrations of 2 μM DDP, 50 μM CBDCA, or 6 μM l-OHP for 24 h. The results, shown at the bottom of Fig. 4, demonstrate significantly increased vesicular accumulation of DDP in both transporter-proficient cell lines (p < 0.008 for MeMNK cells and p < 0.006 for MeWND cells). There was also increased vesicular accumulation of CBDCA, although this reached statistical significance only for the MeMNK cells (p < 0.02 for MeMNK cells and p < 0.09 for MeWND cells). In the case of cells exposed to l-OHP, there was an increase in the ratio of vesicular platinum accumulation in the MeWND cells (p < 0.02) but not in the MeMNK cells. These results provide further support for the notion that, in this cell system, ATP7A and ATP7B function to sequester and detoxify DDP and CBDCA in intracellular vesicles, resulting in increased whole-cell accumulation while conferring resistance to the drugs. l-OHP does not seem to be as efficiently sequestered and detoxified, at least by ATP7A, supporting our findings that l-OHP-treated MeMNK and MeWND cells accumulate increased levels of platinum in their DNA and are hypersensitive to drug treatment.
Immunolocalization of ATP7A in MeMNK Cells by Microscopy. The ability of ATP7A to regulate intracellular copper levels is dependent in part on its ability to relocalize to the plasma membrane in response to copper exposure. We sought to determine whether ATP7A also relocalizes in response to platinum drug treatment. MeMNK cells were treated with equitoxic concentrations of copper, DDP, CBDCA, or l-OHP for 1 h, and the localization of ATP7A was compared with that in untreated cells. Figure 5, A to C, show that in untreated MeMNK cells, ATP7A was found in the perinuclear region consistent with localization in the trans-Golgi (Pena et al., 1999). Figure 5, D to F, show that upon treatment with 500 μM Cu2+, ATP7A redistributed away from the trans-Golgi, verifying the copper-induced relocalization of ATP7A in this experimental model. However, upon treatment with 2 μM DDP (Fig. 5, G-I), 50 μM CBDCA (Fig. 5, J-L), or 10 μM l-OHP (Fig. 5, M-O), ATP7A remained localized mainly to the perinuclear region similar to its location in untreated cells. Thus, although overexpression of ATP7A in this cell system was sufficient to confer resistance to copper, DDP, and CBDCA and to concentrate these platinum drugs into vesicular compartments when exposed to these same drug concentrations, all three of the platinum drugs failed to trigger the trafficking of ATP7A that seems to be essential for its ability to actually export copper from the cell.

Immunofluorescence localization of ATP7A in MeMNK cells after exposure to copper, DDP, CBDCA, or l-OHP. The nuclear marker Hoechst 33342 stains the nucleus blue. Alexa fluor 594-tagged anti-mouse antibody indirectly stains ATP7A red. A-C, perinuclear localization of ATP7A in untreated MeMNK cells; D-F, relocalization of ATP7A in MeMNK cells after a 1-h exposure to 500 μM copper; G-I, perinuclear localization of ATP7A in MeMNK cells after a 1-h exposure to 2 μM DDP; J-L, perinuclear localization of ATP7A in MeMNK cells after a 1-h exposure to 50 μM CBDCA; M-O; perinuclear localization of ATP7A in MeMNK cells after a 1-h exposure to 10 μM l-OHP.
Discussion
Although numerous studies suggest that the development of DDP resistance is mediated in part by changes in DDP influx and/or efflux, the mechanisms by which DDP is transported into and out of cells have remained largely unknown. Recent studies provide evidence that the copper exporter ATP7B (Komatsu et al., 2000; Nakayama et al., 2001, 2002; Kanzaki et al., 2002; Katano et al., 2002; Miyashita et al., 2003; Ohbu et al., 2003) and the major copper influx transporter CTR1 (Ishida et al., 2002; Lin et al., 2002) mediate sensitivity to DDP and modulate its cellular accumulation. The results of the current study confirm the ability of ATP7B to modulate sensitivity to DDP and indicate that this transporter can also render cells resistant to CBDCA. In addition, they provide evidence that ATP7A can also mediate resistance to both DDP and CBDCA.
The approach used in this study has the advantage of permitting comparisons of drug sensitivity and accumulation between isogenic cell lines. The measurement of growth rates during copper exposure and of cellular copper levels confirmed the results of prior studies of copper homeostasis in these cells (La Fontaine et al., 1998). The MeMNK and MeWND cells were more resistant to copper, and during growth in basal or copper-supplemented medium, the copper content was markedly lower in the MeMNK and MeWND cells than in the Me32a cells. Although a detailed comparison of copper efflux kinetics was attempted, the MeMNK and MeWND cells accumulated such small amounts of copper that, even using 64Cu, accurate estimates of initial and terminal rate constants could not be made. Nevertheless, the available data indicate that this group of cell lines constitutes a robust system with which to investigate the effect of ATP7A and ATP7B expression on the cellular pharmacology of the platinum drugs.
Engineering the Me32a cells to express ATP7A or ATP7B clearly rendered them less sensitive to the growth-inhibitory effects of DDP and CBDCA as well as copper. In the case of copper, resistance was accompanied by decreased copper accumulation consistent with the known ability of these exporters to sequester copper into the TGN and vesicles of the secretory pathway and to actually export copper out of the cell. In the case of DDP and CBDCA, resistance was paradoxically accompanied by increased rather than decreased whole-cell accumulation. The increase in whole-cell accumulation was not associated with a similar increase in platinum in DNA for either DDP or CBDCA, indicating that the excess drug did not have access to this critical target. Thus, both ATP7A and ATP7B must increase accumulation of DDP and CBDCA into some other compartment, and this sequestration may serve to detoxify these drugs and to limit the amount of drug that reaches the nucleus. Direct measurement of the platinum levels in vesicles did in fact disclose increased platinum content in vesicles isolated from DDP- and CBDCA-exposed MeMNK and MeWND cells. This compartment is likely to be similar to that into which ATP7A and ATP7B sequester copper. If this is the case, it indicates that there are some fundamental differences in the way in which copper and the platinum drugs are handled, because this export pathway is effective at actually removing copper but not the platinum drugs from the cell. One possibility is that although the platinum drugs serve as substrates for ATP7A- and ATP7B-mediated sequestration, they are not good substrates for the later steps required for actual export from the cell, or that they disable the export machinery because of their ability to react directly with thiol groups in proteins. It is not surprising that that DNA platinum levels after exposure to DDP and CBDCA were not measurably lower in the MeMNK and MeWND cells despite their relative resistance to these drugs, given that the magnitude of the resistance is small, and even large changes in sensitivity are accompanied by only small changes in adduct levels in other cell systems (Johnson et al., 1997).
Given the different spectrum of clinical activity and pattern of toxicity produced by l-OHP (Misset et al., 2000), it is perhaps not surprising that ATP7A and ATP7B expression was found to have different effects on the cellular pharmacology of this drug relative to their effects on DDP and CBDCA. Although both exporters produced an increase in l-OHP whole-cell accumulation in a manner similar to that for DDP and CBDCA, they both rendered cells hypersensitive rather than resistant to l-OHP. The magnitude of this effect was not large, but it was reproducible in multiple experiments, and it was clearly in the opposite direction from the effect of ATP7A and ATP7B on sensitivity to DDP and CBDCA. The implication is that although the copper exporters are capable of sequestering all three of the platinum drugs into some subcellular compartment, in the case of l-OHP, the drug is not inactivated by sequestration. This model is further supported by the finding of increased platinum levels in the DNA of the MeMNK and MeWND cells after l-OHP treatment compared with that for the Me32a cells. Whether some of the excess drug that accumulates in the whole cell finds its way to the nucleus or whether the sequestration of l-OHP into vesicles of the secretory pathway results in some other form of growth-inhibiting injury is currently uncertain.
In another experimental model developed in our laboratory, the human ovarian carcinoma cell line 2008 was transfected with an ATP7B expression vector, and studies of CBDCA sensitivity and accumulation were conducted on the resulting subline (Katano et al., 2003). In this epithelial cell system, increased expression of ATP7B produced resistance to the cytotoxic effect of copper and CBDCA but decreased rather than increased CBDCA accumulation. Increased expression of ATP7B also rendered these cells resistant rather than hypersensitive to l-OHP (K. Katano, unpublished observation).
The reason for the different effects on drug accumulation and sensitivity in the 2008 and Me32a/MeWND/MeMNK systems is not known, but one factor likely to be of importance to the function of ATP7B is the polarity of the cells under study. Previous reports have suggested that ATP7A and ATP7B expression and function are dependent on the polarity of the cells (Harris et al., 1998). There is increasing evidence that the ability of ATP7A and ATP7B to mediate copper export is dependent on their ability to undergo relocalization within the cell after exposure to copper, and it would not be surprising to discover that relocalization occurs differently in fibroblasts and epithelial cells. Another possibility is that although the 2008 cells were transfected with a vector expressing human ATP7B, the MeWND cells express murine ATP7B, and it is conceivable that the murine and human ATP7B proteins function in subtly different ways.
The results of the current study support the notion that expression of ATP7A and ATP7B can alter the sensitivity of cells to all three platinum drugs. It is important to note that neither ATP7A nor ATP7B has formally been shown to be able to transport DDP, CBDCA, or l-OHP, and it remains possible that the effects of these proteins on drug sensitivity and accumulation are mediated indirectly through effects on other transporters or other detoxification mechanisms. Nevertheless, the results reported here provide additional support for the concept that the uptake, intracellular distribution, and efflux of DDP and its analogs is regulated in part by transporters and chaperones that evolved to manage copper homeostasis.
Acknowledgments
We thank Dr. Michael J. Petris of the University of Missouri-Columbia for providing the Me32a, MeMNK, and MeWND cell lines used in this study. We thank Dr. Kevin Walda of the Scripps Institute of Oceanography Analytical Facility for technical assistance and the laboratory of Roger Tsien at the University of California San Diego (UCSD) for use of a Safire microplate reader. We also thank Dennis J. Young in the UCSD Cancer Center's Flow Cytometry Shared Resource for expert advice and Dr. Murray Goodman and Mika Tomioka for synthesis of fDDP.
Footnotes
-
Supported by grants CA95298 from the National Institutes of Health and DAMD17-03-0158 from the Department of Defense. This work was conducted in part by the Clayton Foundation for Research-California Division. R.S. and S.B.H. are Clayton Foundation Investigators. The production of 64Cu at Washington University School of Medicine was supported by the National Cancer Institute grant R24 CA86307.
-
ABBREVIATIONS: CBDCA, carboplatin; DDP, cisplatin; ECFP, enriched cyan fluorescent protein; EYFP, enriched yellow fluorescent protein; fDDP, fluorescein-tagged cisplatin; ICP-MS, inductively coupled plasma mass spectroscopy; ICP-OES, inductively coupled plasma optical emission spectroscopy; l-OHP, oxaliplatin; PBS, phosphate-buffered saline; TGN, trans-Golgi network; Boc, N-tert-butoxycarbonyl.
- Received January 21, 2004.
- Accepted March 17, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}