


Abstract
Tumor-specificity (TS) and anti-inflammatory activity of benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one, generally known as benzoxazinotropone (BOT), have been reported. In order to find a new biological activity, the combination effect of BOT and three apoptosis-inducing agents was investigated. Cytotoxicity against four human oral squamous cell carcinoma (OSCC) cell lines and five human oral normal cells (gingival fibroblasts, periodontal ligament fibroblasts, pulp cells, oral keratinocytes and primary gingival epithelial cells) was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) method. TS was evaluated by the ratio of the mean 50% cytotoxic concentration (CC50) against normal oral cells to the one against OSCC cell lines. Synergy was evaluated by CompuSyn software program. Expression of cleaved forms of poly ADP-ribose polymerase and caspsase-3 was evaluated by western blot analysis. BOT induced activation of caspase 3, suggesting the apoptosis induction in HSC-2 OSCC cells. BOT enhanced the cytotoxicity of doxorubicin (DXR) additively and that of curcumin and resveratrol synergistically. On the other hand, BOT did not enhance, but rather inhibit the cytotoxicity of DXR against normal keratinocytes. The present study suggests that BOT may enhance the anti-tumor activity of apoptosis-inducing agents, while reducing its cytotoxicity against normal cells.
Heterocylic compounds are known to display diverse biological activities (1). Hinokitiol and its related derivatives with a tropolone skeleton (2-4) have been reported to exhibit various biological activities, such as antimicrobial (5), antifungal (6) and phytogrowth-inhibitory activity (7, 8), cytotoxic effects on mammalian tumor cells (9, 10) and inhibitory effects on catechol-O-methyltransferase (11) and metalloproteases (5). Hinokitiol acetate did not exhibit cytotoxic activity (10), antimicrobial activity nor metalloprotease inhibition (5), suggesting that these biological effects of hinokitiol-related compounds may result from the formation of metal chelates by the carbonyl group at C-1 with the hydroxyl group at C-2 in the tropone skeleton.
We have recently reported that benzo[b]cyclohept[e] [1,4]oxazin-6(11H)-one (here referred to as benzoxazinotropone (BOT)) (structure shown in Figure 1) showed slightly (2.2-fold) higher cytotoxicity against human oral squamous cell carcinoma (OSCC) cell lines (HSC-2, HSC-3, HSC-4) than human mesenchymal normal oral cells (human gingival fibroblasts (HGF); human periodontal ligament fibroblasts (HPLF); human pulp cells (HGF)) (12) (Table I) and potently inhibited the production of nitric oxide by lipopolysaccharide-activated mouse macrophage-like RAW264.7 cells via inhibition of inducible nitric oxide synthase and cyclooxygenase-2 protein expression (13). In order to obtain more information of its antitumor potential, we investigated here whether BOT induces apoptosis in HSC-2 cells and whether BOT potentiates cytotoxicity of apoptosis-inducing agents, that is, doxorubicin (DXR), an anthracycline antitumor antibiotic (14), curcumin, a natural component of the rhizome of turmeric (15) and resveratrol, a class of polyphenolic compounds called stilbenes (16) against HSC-2 cells, but not against human oral keratinocytes (HOK).
Cytotoxicity of these compounds was evaluated using both human normal oral cells derived from mesenchymal (HGF, HPLF, HPC) and epithelial tissues (HOK, primary human gingival epithelial cells (HGEP)), as well as human (OSCC) cell lines (Ca9-22, HSC-2, HSC-3, HSC-4). The tumor-selectivity index (TS) was calculated by dividing the mean 50% cytotoxic concentration (CC50) against normal oral cells by that against OSCC cell lines. We have already demonstrated that the TS value determined by this method reflects the antitumor potential of test compounds, although these normal oral cells and OSCC cell lines are classified as different types of cells (mesenchymal or epithelial) (17).
Cytotoxic activity of BOT, curcumin, resveratrol and 5-FU. Each value represents the mean±S.D. of three independent experiments performed in triplicate, except for 5-FU (just one experiment performed in triplicate).
Materials and Methods
Materials. The following chemicals and reagents were obtained from the indicated companies: Dulbecco's modified Eagle's medium (DMEM) from GIBCO BRL, Grand Island, NY, USA; fetal bovine serum (FBS), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), doxorubicin-HCl (DXR), resveratrol from Sigma-Aldrich Inc., St. Louis, MO, USA; dimethyl sulfoxide (DMSO) from Wako Pure Chem. Ind., Osaka, Japan; curcumin from Tokyo Kasei Kogyo Co., Tokyo, Japan); 5-fluorouracil (5-FU) from Kyowa, Tokyo, Japan; Culture plastic dishes and plates (96-well) were purchased from Becton Dickinson (Franklin Lakes, NJ, USA). Benzoxazinotropone (BOT) was synthesized by the reaction of 3-bromo-2-methoxytropone with o-aminophenol as described previously (18, 19). BOT, doxorubicin, curcumin and resveratrol were dissolved in DMSO at 80, 1, 40, 40 mM and stored at −20°C before use. Protease and phosphatase inhibitors were purchased from Roche Diagnostics (Tokyo, Japan). Antibodies against cleaved caspase-3 (Cell Signaling Technology Inc., Beverly, MD, USA), PARP (Cell Signaling Technology Inc.) and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Trevigen, Gaithersburg, MD, USA) were used as primary antibodies. As secondary antibodies, α-rabbit IgG (DAKO Japan) antibodies, which were conjugated with horseradish peroxidase, were used.

Structure of tropone and benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one (BOT).
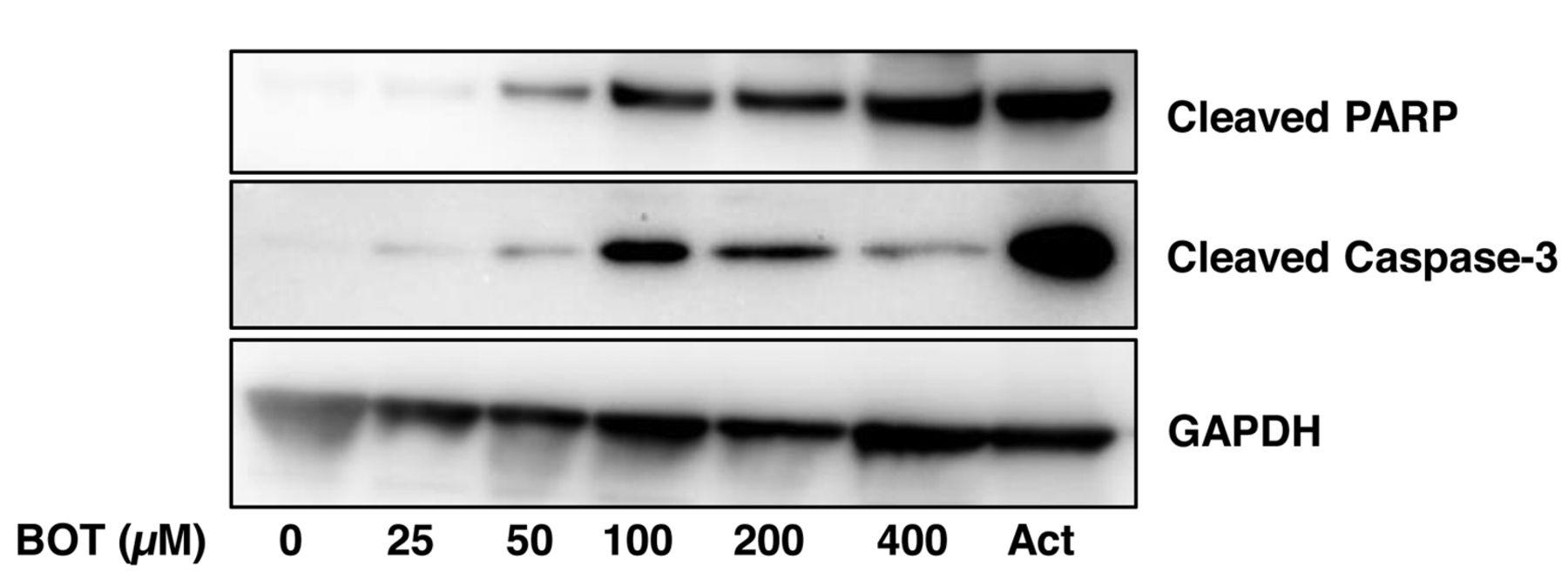
Induction of apoptosis markers by benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one (BOT) in human OSCC HSC-2 cells. Near-confluent HSC-2 cells were incubated for 24 h with the indicated concentrations (25~400 μM) of BOT and expression of cleaved products of PARP and caspase-3 was visualized by western blot analysis. Each sample contains 0.1% DMSO. Act, Actinomycin D (2 μM), was used as an apoptosis-positive control.
Cell culture. HGF, HPLF and HPC cells, established from the first premolar tooth extracted from the lower jaw of a 12-year-old girl (20) and OSCC cell lines (Ca9-22, HSC-2, HSC-3, HSC-4) purchased from Riken Cell Bank, Tsukuba, Japan were cultured at 37°C in DMEM supplemented with 10% heat-inactivated FBS, 100 units/ml penicillin G and 100 μg/ml streptomycin sulfate under a humidified 5% CO2 atmosphere. HOK (purchased from COSMO BIO Co/ Ltd., Tokyo, Japan) was cultured in keratinocyte growth supplement (OKGS, Cat, No. 2652; Science Research laboratories, Carlsbad, CA, USA). Primary human gingival epithelial cells (HGEP) (purchased from CELLnTEC Advanced Cell Systems AG, Bern, Switzerland) were grown in CnT-PR medium (CELLnTEC). Cells were then harvested by treatment with 0.25% trypsin-0.025% EDTA-2Na in phosphate-buffered saline without calcium and magnesium [PBS(−)] and subcultured or used for experiments.
Assay for cytotoxic activity. Cells were inoculated at 2.5×103 cells/0.1 ml in a 96-microwell plate (Becton Dickinson Labware, Franklin Lakes, NJ, USA). After 48 hours, the medium was removed by suction with an aspirator and replaced with 0.1 ml of fresh medium containing different concentrations of test compounds in single or in combination with BOT. Control cells were treated with the same amounts of DMSO present in each diluent solution. Cells were incubated for 48 hours and the relative viable cell number was then determined by the MTT method. In brief, the treated cells were incubated for another 3 hours in fresh culture medium containing 0.2 mg/ml MTT. Cells were then lysed with 0.1 ml of DMSO and the absorbance at 540 nm of the cell lysate was determined using a microplate reader (Sunrise Rain bow RC-R; TECAN, Mannedorf Switzerland). The CC50 was determined from the dose–response curve and the mean CC50 for each cell type was calculated from three independent experiments.
Calculation of TS. When normal oral mesenchymal cells (HGF, HPLF, HPC) were used, TS was calculated by the following equation: TS=mean CC50 against HGF, HPLF and HPC cells / mean CC50 against OSCC cell lines (shown as (B/A) in Table I).
When normal oral epithelial cells (HOK, HGEP) were used, TS was calculated by the following equation: TS=mean CC50 against HOK and HGEP/mean CC50 against OSCC cell lines (shown as (C/A) in Table I).
Western blot analysis. The cells were washed with phosphate-buffered saline (PBS) and re-suspended in 50 mM Tris-HCl (pH 7.6), 150 mM NaCl, 1 mM EDTA, 0.1% sodium dodecyl sulfate (SDS), 0.5% deoxycholic acid, 1% NP-40 and protease inhibitors (RIPA buffer). After ultrasonication using Bioruptor (UCD-250; Cosmo Bio) for 12.5 min (10 sec on, 20 sec off) at the middle level of output (250 W) at 4°C, the soluble cellular extracts were recovered after centrifugation for 10 min at 16,000 × g. The protein concentration of each sample was determined using the BCA Protein Assay Reagent Kit (Thermo Fisher Scientific) and cell extracts were subjected to Western blot (WB) analysis. The blots were probed with the primary antibody followed by a horseradish peroxidase-conjugated secondary antibody. The immune complexes were visualized using Pierce Western Blotting Substrate Plus (Thermo Fisher Scientific). WB results were documented and quantified using ImageQuant LAS 500 (GE Healthcare, Tokyo, Japan).
Statistical treatment. Each experimental value is expressed as the mean±standard deviation (SD) of three independent experiments that were performed in triplicate in most of experiments. The CompuSyn software program (ComboSyn, Inc., Paramus, NJ, USA) was used to calculate the combination index (CI) (21). Briefly, CI value was determined by the following equation: CI=(% decrease of viable cell number by compound A + % decrease of viable cell number by compound B)/(% decrease of viable cell number by combination of compound A and B). The combination effect was judged “synergistic”, “additive” or “antagonistic” when CI is <1, 1 and >1, respectively.
Results
Apoptosis induction. WB analysis demonstrated that BOT dose-dependently induced the activation of caspase 3 (detected by the active form of caspase 3, that is, cleaved caspase and the cleavage of PARP, one of the substrates of caspase 3) (22). The effects of BOT became discernible above 25 μM and reached plateau at 50~400 μM. These results suggest that BOT induced apoptosis in HSC-2 cells but at a slightly lower level than that achieved by actinomycin D, a positive control of apoptosis (Figure 2).
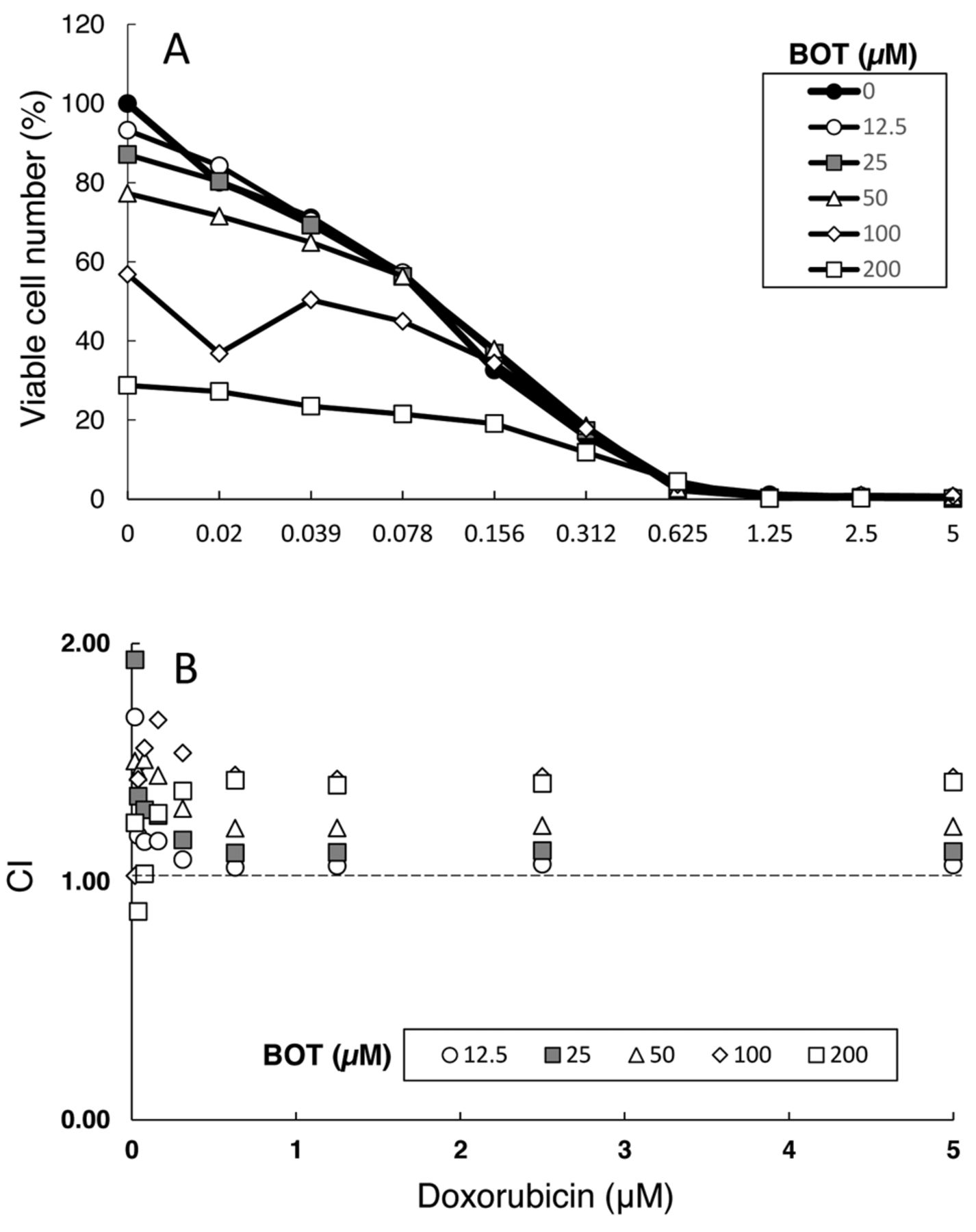
Combination effect of doxorubicin and benzo[b] cyclohept[e][1,4]oxazin-6(11H)-one (BOT) on the viable cell number of HSC-2 cells. (A) Near-confluent HSC-2 cells were incubated for 48 h with the indicated concentrations (0.02~5 μM) of doxorubicin (DXR) in the presence of 12.5, 25, 50, 100 or 200 μM of BOT and, then, viable cell number was determined by MTT method. Each value represents mean of three independent experiments. (B) Combination index (CI), as calculated from data of A.
Combination with DXR. DXR (0.02~5 μM) dose-dependently reduced the viable cell numbers of HSC-2 cells, while addition of increasing concentrations (12.5~200 μM) of BOT further reduced the viable cell numbers of HSC-2 cells (Figure 3A). Calculation of combination index (CI) revealed that lower concentrations (12.5 and 25 μM) of BOT showed a somewhat additive effect (CI=1), whereas higher concentrations were rather antagonistic (CI>1) (Figure 3B).
DXR showed bi-phasic effect on growth of HOK cells, known as hormesis (23), stimulating the growth at lower concentrations and inhibiting the growth at higher concentrations (Figure 4). Simultaneous addition of BOT did not enhance, but rather tended to inhibit, the cytotoxicity of DXR (Figure 4). This suggests that BOT protected the cells from DXR-induced cytotoxicity.

Protective effect of benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one (BOT) on doxorubicin (DXR)-induced cytotoxicity against HOK cells. HOK cells were incubated for 48 h with the indicated concentrations (0.02~5 μM) of DXR in the presence of 12.5, 25, 50, 100 or 200 μM of BOT and, then, viable cell number was determined by MTT method. Each value represents mean of triplicate assays.
Combination with curcumin and resveratrol. Since there was no detailed study of tumor-specificity of curcumin and resveratrol, we first investigated their cytotoxicity against human oral OSCC (Ca9-22, HSC-2, HSC-3, HSC-4) and human oral normal cells (HGF, HPLF, HPC, HOK, HGEP) (Table I). We found that curcumin and resveratrol gave TS (B/A) values of 3.9 and 3.1, respectively, about 3~4-fold less than that of 5-FU (TS>12.0) when normal mesenchymal cells (HGF, HPLF, HPC) were used (Table I). These polyphenols gave very low TS (C/A) values of 1.5 and 1.3, respectively, about one half that of 5-FU (TS=3.0), when normal epithelial cells (HOK, HGEP) were used (Table I).
We next investigated the combination effect of BOT and these polyphenols. BOT and curcumin or resveratrol gave additive cytotoxic effects on the growth of HSC-2 cells (Figure 5A and B). In particular, when the concentrations of these polyphenols were below 12.5 μM, synergistic inhibition of the growth of HSC-2 cells was observed.
Discussion
The present study demonstrated, for the first time, that relatively lower concentrations (12.5 and 25 μM) of BOT enhances the cytotoxicity of DXR against OSCC cells (HSC-2) in an additive fashion, while BOT slightly protected HOK cells from the cytotoxicity of DXR. This suggests that appropriate concentrations of BOT may potentiate the antitumor action of DXR.

Combination effect of benzo[b]cyclohept[e][1,4]oxazin-6(11H)-one (BOT) and curcumin or resveratrol on viable cell numbers of HSC-2 cells. (A) Near-confluent HSC-2 cells were treated (in triplicate) for 48 h with 0, 12.5, 25, 50, 100 or 200 μM of curcumin (A) or resveratrol (B) in the presence of 12.5, 25, 50, 100 or 200 μM of BOT and, then, viable cell numbers were determined by MTT method. From the dose-response curve, combination index (CI) was determined and plotted as a function of concentration of curcumin or resveratrol.
The other new finding of the present study is that BOT dose-dependently enhanced the cytotoxicity of curcumin and resveratrol against HSC-2 cells, additively or synergistically (Figure 5). Additivity was found when relatively higher concentrations of curcumin (25~200 μM) and resveratrol (12.5~200 μM) were used, whereas synergy was observed when relatively lower concentrations of curcumin (3.1~12.5 μM) and resveratrol (3.1~6.3 μM) were employed. The biological significance of the present study is unclear. Further study is needed to discriminate whether the present finding is unique only to BOT or universally observed in all benzoxazinotropone family.
We recently reported that human oral epithelial cells (HOK, HGEP) were highly sensitive to DXR and 5-FU (24-26). This finding was supported by the present study. Studies are underway to search for natural and/or synthetic compounds that alleviate the cytotoxicity of these antitumor agents against normal oral epithelial cells.
Acknowledgements
This work was supported by Grant-in-Aid for Challenging Exploratory Research from The Ministry of Education, Culture, Sports, Science and Technology (Sakagami H. 25670897), research funds from Meikai University School of Dentistry.
Footnotes
↵* These Authors contributed equally to this work.
Conflicts of Interest
We confirm that there are no known conflicts of interest associated with this publication and there was no significant financial support for this work that could have influenced its outcome.
- Received April 27, 2016.
- Revision received June 5, 2016.
- Accepted June 6, 2016.
- Copyright © 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Potentiation of Anticancer Activity of G2/M Blockers by Mild Hyperthermia
- In Vitro Assessment of Antitumor Potential and Combination Effect of Classical and Molecular-targeted Anticancer Drugs
- Quantitative Structure-Cytotoxicity Relationship of Azulene Amide Derivatives
- Quantitative Structure-Cytotoxicity Relationship of 3-(N-Cyclicamino)chromone Derivatives
- Quantitative Structure-Cytotoxicity Relationship of Pyrano[4,3-b]chromones
- Quantitative Structure-Cytotoxicity Relationship of 2-(N-cyclicamino)chromone Derivatives
- In Vitro Antitumor Activity of Alkylaminoguaiazulenes
- In Vitro Anti-tumor Activity of Azulene Amide Derivatives