


Abstract
Amentoflavone, isolated from an ethyl acetate extract of the whole plant of Selaginella tamariscina, a traditional herb, may exhibit antitumor activity. The aim of this study was to investigate the anticancer mechanism(s) of amentoflavone, such as mitochondria-mediated apoptotic cell death, in typical breast cancer MCF-7 cells. Cells treated with amentoflavone exhibited a series of cellular alterations related to apoptosis, including DNA and nuclear fragmentation, and de-regulation of intracellular reactive oxygen species (ROS) and calcium. In addition, markers of mitochondrial-mediated apoptosis, including the reduction of mitochondrial inner-membrane potential, the release of cytochrome c from mitochondria, and activation of caspase 3, were observed. In conclusion, our results present, to our knowledge, the first evidence that amentoflavone induces apoptosis of MCF-7 breast cancer cells, and that this is closely related to mitochondrial dysfunction. Amentoflavone may be a potential therapeutic agent for breast cancer treatment.
Breast cancer is the most common type of invasive cancer among women worldwide, responsible for over 458,503 deaths (13.7% of cancer deaths in women and 6.0% of cancer deaths in both genders), in 2008 (1, 2). The fight against breast cancer attracts attention from all who are searching for more potent anticancer drugs for cancer patients, including medical doctors and basic scientists. The failure of apoptosis induction in breast cancer cells is the primary obstacle that limits the therapeutic efficacy of anticancer agents, and hence the development of powerful apoptosis-inducing agents for cancer has become an urgent mission for translational researchers.
Flavonoids are naturally-occurring compounds present in a variety of fruits, vegetables and seeds, and in Chinese herbal medicines (3, 4). They have many biological properties including antioxidative, anti-inflammatory and antifungal effects. Growing lines of evidence have demonstrated that flavonoids are neuroprotective in a variety of cellular and animal models of neurodegenerative diseases, primarily due to their antioxidative properties (5-8). However, the antitumor capacity of flavonoids has not been extensively evaluated, and their mechanisms of action are not well-elucidated. Amentoflavone, 8-(5-(5,7-dihydroxy-4-oxo-4H-chromen-2-yl)-2-hydroxyphenyl)-5,7-dihydroxy-2-(4-hydroxyphenyl)-4H-chromen-4-one, belonging to the biflavonoid class of flavonoids, is abundant in Selaginella tamariscina and has been used for the treatment of cancer in traditional Chinese medicine. It has also been used as an antioxidant, vasorelaxant, anti-HIV and anti-angiogenic agent (9-12). Nonetheless, to our knowledge there have been no reports on the possible effects of amentoflavone on apoptosis, which plays an important role in cancer cell death.
In this study, we investigated the growth-inhibition and cell-cycle arrest effects of amentoflavone on MCF-7 cells, as well as on the induction of apoptosis. Furthermore, we examined the intracellular mechanisms involved in the process of apoptosis induction.
Materials and Methods
Chemicals. Amentoflavone, dimethyl sulfoxide (DMSO), propidium iodide (PI), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) and 4’-6-diamidino-2-phenylindole (DAPI) were purchased from Sigma-Aldrich Corp. (St. Louis, MO, USA). All primary and secondary antibodies were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA). The fluorescent probes 2’,7’-dichlorofluorescin diacetate (DCFH-DA), Indo 1/AM and 3, 3’-dihexyloxacarbocyanineiodide (DiOC6) were from Invitrogen Life Technologies (Carlsbad, CA, USA). Dulbecco's modified Eagle's medium (DMEM) and penicillin/streptomycin were obtained from GIBCO/BRL Life Technologies (Cambrex, Walkersville, MD, USA).
MCF-7 cell culture. MCF-7 human breast cancer cell line was obtained from the American Type Culture Collection (Manassas, VA, USA). The cells were cultured in DMEM supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT, USA), 100 mM non-essential amino acid, 2 mM glutamate, 100 U/ml penicillin, and 100 μg/ml streptomycin. The cultures were incubated at 37°C in a humidified atmosphere with 5% CO2. Cells were sub-cultured every 2-3 days to obtain exponential growth.
Observation of morphological alterations. Approximately 2×105 MCF-7 cells/well in 24-well plates were treated with 250 μM amentoflavone, or with vehicle (DMSO, 0.1% in culture media)-only and all cells were incubated for 6 to 72 h. For the examination of morphological changes, cells were photographed under a phase-contrast microscope.
Cell viability measurement. For determining the cell viability, the MTT assay was performed as previously published (13). Briefly, cells were plated into 24-well plates at a density of 3×104 MCF-7 cells/well, and grown for another 24 h, then treated with 0.1% DMSO or different concentrations of amentoflavone. After treatment, MTT was added to each well at a final concentration of 0.5 mg/ml and the mixture of MTT and cells was further incubated at 37°C for 4 h. The viable cell number was directly proportional to the production of formazan following its solubilization with isopropanol. The color intensity was measured at 570 nm in a Multiskan MS ELISA reader (Labsystems, Helsinki, Finland). The experiments were performed in triplicate.
Cell-cycle distribution determination. About 2×106/ml MCF-7 cells were seeded in 10-cm dishes with and without 250 μM amentoflavone, and all cells were incubated for 0, 6, 12, 24, 48 and 72 h. After incubation, the cells were harvested and fixed gently with 70% ethanol, washed twice with PBS and then incubated with PI buffer (4 μg/ml PI, 0.5 μg/ml RNase, and 1% Triton X-100 in PBS) for 30 min in the dark at room temperature. The cells were then filtered through a 40-μm nylon filter and the PI-stained cells were analyzed for cell-cycle distribution and apoptosis (sub-G1 phase) using a FACS Calibur instrument (BD Biosciences, San Jose, CA, USA) equipped with Cell Quest software, as described previously (14-19). The sub-G1 group was representative of mean apoptosis.
Nucleic acid condensation and DNA damage. For DNA damage investigation, the comet assay was performed as previously published (14, 15), and the comet moment was calculated according to the formula “comet moment=Σ0→n [(amount of DNA at distance X)×(distance X)]/total DNA”, as given in our previous publications (20, 21).
For the DNA fragmentation assay, MCF-7 cells were plated at 2×105 cells/ml in 12-well plates and amentoflavone added for 48 h. Both attached and detached cells were collected and lysed in 100 mM Tris (pH 8.0), 20 mM EDTA and 0.8% N-laurylsarcosine sodium salt on ice. The lysates were incubated with 0.2 mg/ml RNase A for 1 h at 37°C, then with 4 mg/ml proteinase K overnight at 50°C. The DNA was precipitated by adding an equal volume of isopropanol, and loaded onto 2% DNA agarose gels and electrophoresed in 1× Tris-Borate-EDTA (TBE) buffer for 3 h. Gels were photographed under UV light.
DNA ladder observation. MCF-7 cells at 2×105 cells/well, in 12-well plates were cultured with 0, 100, 150, 200, 250 and 300 μM amentoflavone for 24 h. DNA was isolated (Genomic DNA purification kit; Genemark Technology Co., Ltd., Tainan, Taiwan) and the ladder formation assay was carried out, as previously described (14, 15).
Analysis of mitochondrial membrane potential loss, reactive oxygen species (ROS) and Ca+2 release. About 2×105 MCF-7 cells/well in 12-well plates were incubated with 250 μM amentoflavone for 0, 24, 48, and 72 h, to determine the change in mitochondrial membrane potential (ΔΨm) and the productions of ROS and cytosolic Ca+2. After cells were incubated for different time periods, all the cells under each treatment were harvested, washed twice with PBS, then re-suspended in 500 μl of 1 μM DiOC6 for determination of ΔΨm, 500 μl of 10 μM DCFH-DA for ROS and in Indo 1/AM (3 μg/ml) for cytosolic Ca+2 production at 37°C in the dark for 30 min. Then all samples were analyzed immediately by flow cytometry, as previously described (14, 15).
Western blotting. About 5×106 MCF-7 cells/well in 6-well plates were incubated with 10 μM amentoflavone for 0, 6, 12, 24, and 48 h. The cells from each treatment were harvested and washed twice with PBS for the determination of levels of caspase-3, apoptosis inducing factor (AIF), B-cell lymphoma 2 (BCL2), BCL2-associated X protein (BAX), BH3 interacting domain death agonist (BID) and p53 proteins, associated with apoptosis which were determined by western blotting. Lysates of treated cells from each well were prepared using lysis buffer, as described previously (14, 15). Briefly, each sample was incubated with primary and secondary antibodies, detected by the ECL reagent kit (Millipore, Billerica, MA, USA) and then by autoradiography using X-ray film. Each membrane was re-probed with anti-β-actin antibody to ensure equal protein loading. The image is the outcome of protein quantification by NIH image processing and analysis in java software (ImageJ).
Statistical analysis. The quantitative data are shown as the mean±SD. The statistical differences between the amentoflavone-treated and control samples were calculated by Student's t-test. p-Values of less than 0.05 were considered statistically significant. Results are representative of at least three independent experiments.

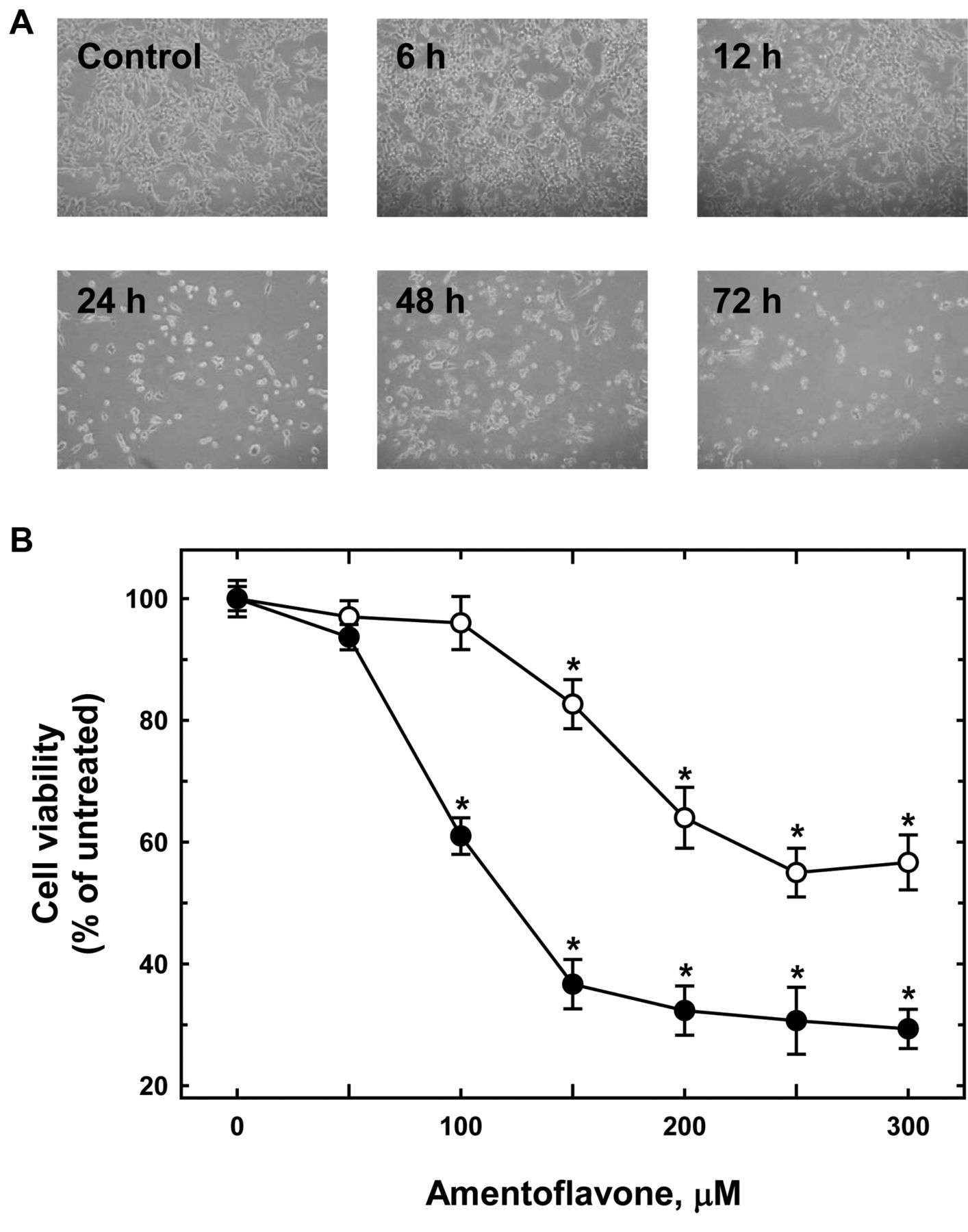
Amentoflavone induced cell morphological changes and reduced the percentage of viable MCF-7 cells. A: Cells were cultured with 250 μM amentoflavone for 6 to 72 h. The cell morphological changes were examined and photographed by phase-contrast microscopy after 6, 12, 24, 48 and 72 h treatments. B: The percentage of viable MCF-7 cells was determined as described in the Materials and Methods. Each point is the mean±SD of three experiments. *p<0.05, significantly different comparing the DMSO-treated control and amentoflavone treatment.
Results
Amentoflavone induced morphological changes and reduced the percentage of viable MCF-7 cells. MCF-7 cells were treated with different doses of amentoflavone for 6, 12, 24, 48 and 72 h, and followingly the morphological changes were investigated using photographs taken under a phase-contrast microscope. The results showed that amentoflavone induced obvious morphological changes time-dependently (Figure 1A). Incubation of cells with amentoflavone for both 24 and 48 h dose-dependently reduced the cell viability of MCF-7 cells. This cycotoxic effect of amentoflavone to MCFR-7 cells was significant at concentrations of 150 μM and more for both 24 and 48 h (Figure 1B).
Amentoflavone induced apoptosis in MCF-7 cells. MCF-7 cells were treated with 250 μM of amentoflavone for 6 to 72 h, and harvested to determine alterations in the cell-cycle distribution and investigate the increase of the typical sub-G1 (apoptosis) phase. The number of cells at each stage of the cell cycle were calculated as a percentage of the overall number of cells and the results are shown in Figure 2. The results indicate that amentoflavone induced apoptosis time-dependently, with an obvious decrease of cells in the G0/G1 and G2/M phases, especially after 24 h. The percentage of apoptotic MCF-7 cells reached almost 50% after 72-h exposure to amentoflavone (Figure 2).
Amentoflavone–induced chromatin condensation and DNA strand breaks in MCF-7 cells. MCF-7 cells were treated with 100 to 300 μM of amentoflacone for 24 h and then harvested for determination of the apoptosis by DAPI staining, for DNA damage by comet assay and for DNA fragmentation by DNA gel electrophoresis. The data are presented in Figure 3. DAPI staining assay demonstrated that amentoflavone induced chromatin condensation and apoptosis in a dose-dependent manner (Figure 3A). The comet assay demonstrated that 100-300 μM amentoflavone for 24 h also induced DNA damage in a dose-dependent manner (Figure 3B). DMSO- and H2O2-treated cells served as negative and positive controls for comet observation, respectively (Figure 3B and C). DNA ladder gel electrophoresis indicated that amentoflavone induced DNA fragmentation in a dose-dependent manner (Figure 3D).
Amentoflavone affected ΔΨm, ROS and cytosolic Ca+2 in MCF-7 cells. MCF-7 cells were treated with 250 μM amentoflavone for different periods of time and the levels of ΔΨm, ROS and cytosolic Ca+2 release were measured by flow cytometry. The results are presented in Figure 4. There was a significant decrease in ΔΨm (Figure 4A and B) and an increase in intracellular ROS (Figure 4C), and cytosolic Ca+2 (Figure 4D) observed in 250-μM amentoflavone-treated cells. Figures 4A and B indicate that amentoflavone significantly reduced ΔΨm in MCF-7 cells in a time-dependent manner (Figure 4A and B).
After 6 h of treatment with amentoflavone, cytosolic the Ca+2 levels significantly increased. After the rapid increase, the cytosolic Ca+2 levels slowly decreased and returned to the levels of the control at 72 h (Figure 4D). It is interesting that there was a biphasic change of ROS levels. The ROS levels were elevated at 1 h after amentoflavone treatment, and reached a plateau at 3 to 12 h. The ROS levels were significantly lower at 24 h after amentoflavone treatment, and this decreasing trend was maintained until 72 h (Figure 4C).
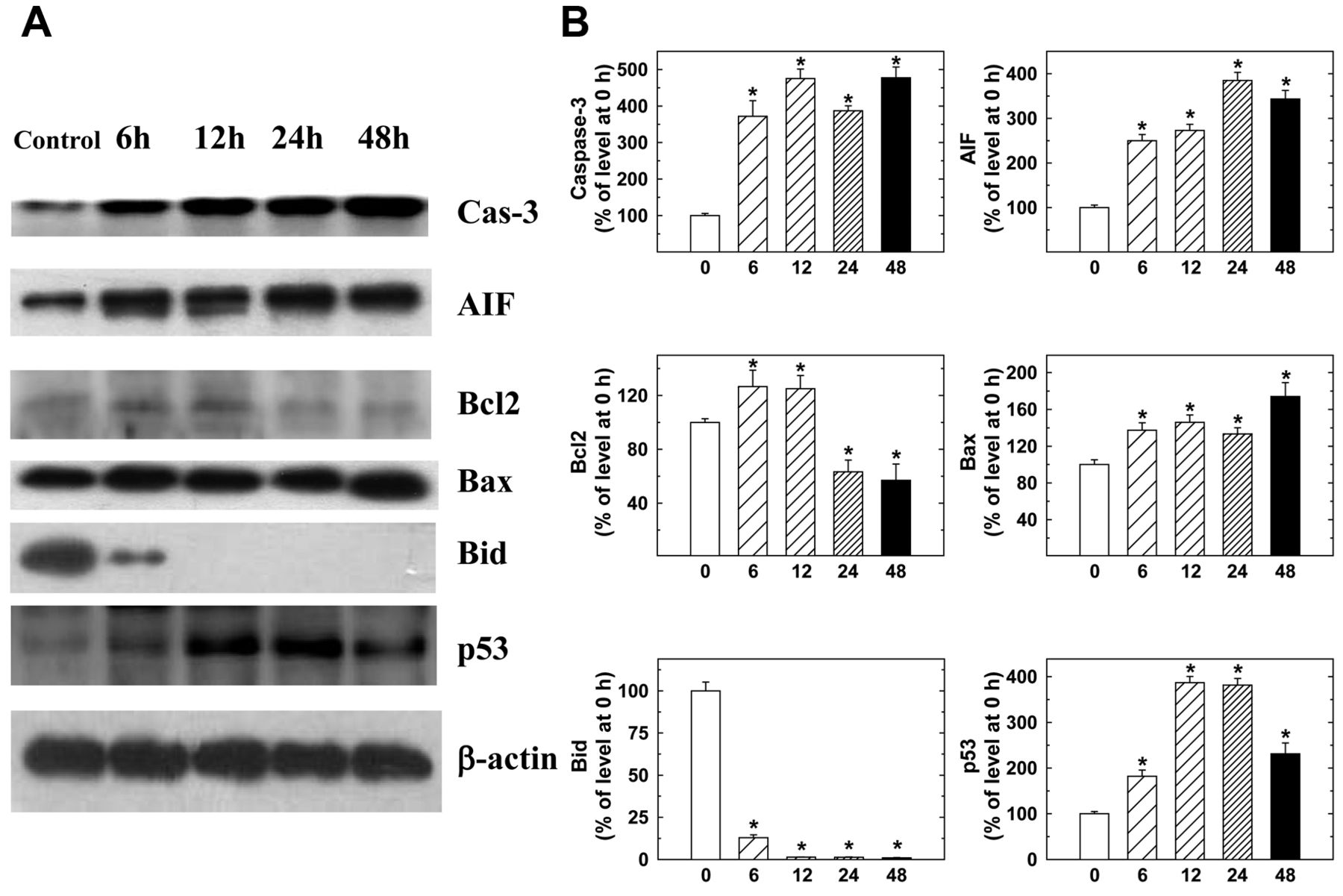
Amentoflavone altered the expression profile of apoptosis-associated proteins in MCF-7 cells. MCF-7 cells were treated with 250 μM amentoflavone for 0, 6, 12, 24, and 48 h and were then harvested for determination of apoptosis-associated proteins caspase-3, AIF, BCL2, BAX, BID and p53 (Figure 5). The level of the antiapoptotic protein BCL2 was decreased after 24 h, and that of pro-apoptotic protein BAX was up-regulated in MCF-7 cells, together with AIF, after exposure to amentoflavone. p53 expression was also increased, which could contribute to amentoflavone-induced programmed cell death of MCF-7 cells. At the same time, BID was decreased following 6 h of treatment (Figure 5).

Amentoflavone affected the cell-cycle distribution and apoptosis of MCF-7 cells. Cells were cultured with 250 μM amentoflavone for 0, 6, 12, 24, 48 and 72 h. The cells were examined and analyzed for cell-cycle distribution and sub-G1 phase cells (apoptosis) by flow cytometry as described in the Materials and Methods. Each point is the mean±SD of three experiments. *p<0.05, significantly different comparing the DMSO-treated control and amentoflavone-treated groups.
Discussion
In the present study, we have provided the first evidence that amentoflavone is an apoptosis-inducing agent in MCF-7 cells via a mitochondria-dependent pathway. It might be a potential anticancer drug for breast cancer prevention and therapy based on the observations that amentoflavone induced morphological changes and reduced the viability of MCF-7 cells, increased the proportion of sub-G1 MCF-7 cells, induced chromatin condensation and DNA strand breaks in MCF-7 cells, reduced mitochondrial membrane potential, increased ROS (within 12 h) and cytosolic Ca+2 release, and altered the expression of apoptosis-associated proteins.
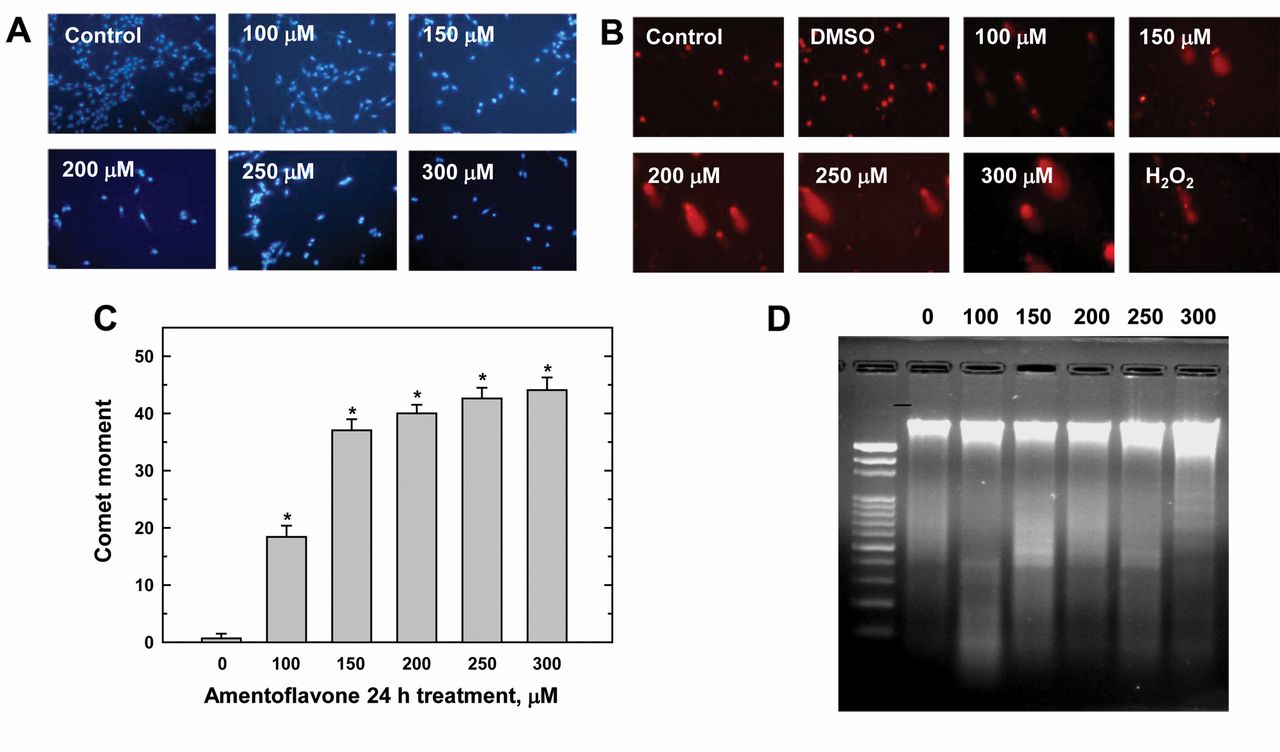
Amentoflavone induced chromatin condensation and DNA strand breaks in MCF-7 cells. Cells were incubated with different concentrations of amentoflavone for 24 h. The cells were harvested and were examined for chromatin condensation by 4’, 6-diamidino-2-phenylindole (DAPI) staining (A) and for DNA damage by comet assay (B) with quantification of fluorescence intensity (fold-difference between control and amentoflavone treatment) and comet tail (percentage difference between control and amentoflavone treatment) (C) DNA fragmentation (D) was assessed by DNA gel electrophoresis as described in the Materials and Methods. Each point is the mean±SD of three experiments. *p<0.05, significantly different comparing DMSO-treated control and amentoflavone-treated groups.
The treatment with 250 μM amentoflavone induced almost half of the MCF-7 cells to undergo programmed cell death after 72 h (Figure 2). Unlike the G0/G1 or G2/M phases, which were dramatically reduced by 250 μM amentoflavone, the S phase persisted at about 20% during the 72-h treatment period (Figure 2). Thus, it is reasonable to propose that amentoflavone may induce the cell cycle to be arrested at the S phase, rather than at G0/G2 or G2/M, as other anticancer drugs do. The detailed mechanisms remain of action to be further investigated.
Caspases play important roles in programmed cell death (22, 23). Caspase-8 and -9 are recognized as the extrinsic initiator and mitochondria-associated caspase, respectively (24). Both caspase-8 and -9 can activate the downstream caspase-3 (25), but in a few cases, only one of these induced activation of caspase-3 (26). In this study, we also examined the activation of caspase-8 and -9 in amentoflavone-treated MCF-7 cells. The results showed that the protein levels of caspase-9 were increased but not as significantly as those of caspase-3 (Figure 5). However, caspase-8 expression was not obviously altered (data not shown). The activation of caspase-3 and -9, but not caspase-8 was also confirmed by flow cytometry (data not shown). The almost complete loss of mitochondria membrane potential provides another piece of evidence that the mitochondria are involved in amentoflavone-induced MCF-7 apoptosis (Figure 4).
To sum up, we can conclude that amentoflavone-induced apoptosis in MCF-7 human breast cancer cells occurs via the mitochondria-dependent pathway. Recently, amentoflavone was found to be effective in suppressing fatty acid synthase expression in human epidermal growth factor receptor 2 (HER2)-positive human breast cancer SKBR3 cells, together with inducing apoptosis (27). It is interesting that amentoflavone may induce apoptosis via different mechanisms in different breast cancer cell lines. Similar to our findings, amentoflavone was also reported to induce cell-cycle arrest at the sub-G1 phase, and apoptosis via mitochondria-emanated intrinsic pathways in SiHa and CaSki human cervical cancer cells (28). In an animal model, amentoflavone was found to significantly inhibit B16F-10 melanoma-induced solid tumor development in C57BL/6 mice (29) and the alteration of BCL2, p53 and caspase-3 expression pattern in amentoflavone-treated B16F-10 melanoma cells (29) was consistent with our findings in MCF-7 cells.
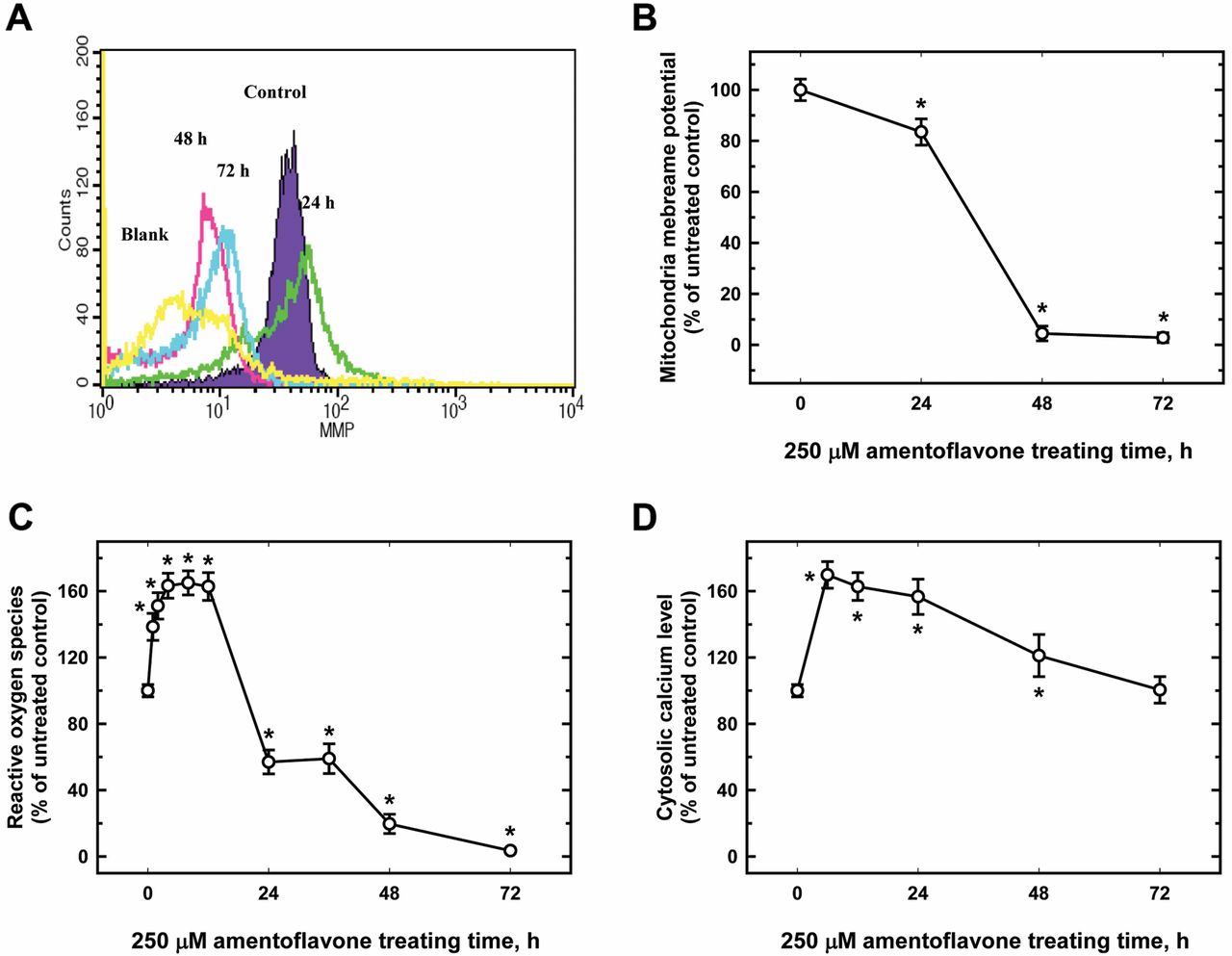
Amentoflavone affected the mitochondria membrane potential (ΔΨm), reactive oxygen species (ROS), and cytosolic Ca+2 in MCF-7 cells. Cells were treated with 250 μM amentoflavone for 0, 24, 48, or 72 h before being collected, and stained with 3, 3’-dihexyloxacarbocyanineiodide (DiOC6) (1 μM) for ΔΨm (A) 2’, 7’-dichlorofluorescein diacetate (DCFH-DA) (10 μM) for ROS (B) and Indo 1/AM (3 μg/ml) for cytosolic Ca+2 production (C) as described in the Materials and Methods. Each experiment was carried out with triple sets (data are the mean±SD): *p<0.05, significantly different comparing DMSO-treated control and amentoflavone-treated groups.
In conclusion, this study provides evidence that amentoflavone may induce MCF-7 cells to undergo cell-cycle arrest and programmed cell death via the ROS- and Ca+2-involved mitochondria-dependent pathway. Our findings provide evidence for amentoflavone being a potential anti-breast cancer agent by confirming its efficacy towards MCF-7 breast cancer cells.

Representative western blotting showing changes in the levels of apoptosis-associated proteins in MCF-7 cells after amentoflavone exposure. Cells were treated with 250 μM amentoflavone for 0, 6, 12, 24 and 48 h before the total proteins were prepared and determined as described in the Materials and Methods. The expression of apoptosis-related proteins caspase-3, apoptosis inducing factor (AIF), B-cell lymphoma 2 (BCL2), BCL2-associated X protein (BAX), BH3 interacting domain death agonist (BID), and p53 were estimated by western blotting analysis and quantified as described in the Materials and Methods.
Acknowledgements
This study was supported by a research grant from TaoYuan General Hospital (number PTH10031). The assistance from Sue-Fung Chen, Yi-Ting Chang, Hong-Xue Ji at the Terry Fox Cancer Research Laboratory of the China Medical University was highly appreciated by the Authors.
Footnotes
-
↵* These Authors contributed equally to this work.
- Received July 11, 2012.
- Revision received September 25, 2012.
- Accepted September 27, 2012.
- Copyright © 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Lenvatinib Suppresses Protein Kinase B Signaling and Induces Apoptosis in Osteosarcoma Cells
- Inactivation of AKT/ERK Signaling and Induction of Apoptosis Are Associated With Amentoflavone Sensitization of Hepatocellular Carcinoma to Lenvatinib
- Amentoflavone Induces Apoptosis and Reduces Expression of Anti-apoptotic and Metastasis-associated Proteins in Bladder Cancer
- Amentoflavone Inhibits ERK-modulated Tumor Progression in Hepatocellular Carcinoma In Vitro
- Amentoflavone Induces Apoptosis and Inhibits NF-ĸB-modulated Anti-apoptotic Signaling in Glioblastoma Cells
- Baicalin Scavenged Reactive Oxygen Species and Protected Human Keratinocytes Against UVB-induced Cytotoxicity
- Amentoflavone Induces Anti-angiogenic and Anti-metastatic Effects Through Suppression of NF-{kappa}B Activation in MCF-7 cells
- Protective Effects of Pyridoxamine Against UVC-induced Programmed Cell Death in HaCaT Cells
- Baicalin Scavenges Reactive Oxygen Species and Protects Human Keratinocytes Against UVC-induced Cytotoxicity