Article Text

Abstract
Objective To examine the role of CD10, a characteristic marker of liver metastasis of colorectal cancers (CRCs).
Design The effect of CD10 and Met-enkephalin (MENK) in CD10-positive and -negative human CRC cells was investigated under in vitro and in vivo conditions. Human CRC samples were examined.
Main outcome measure CD10-positive and CD10-knockdown HT29 cells and CD10-negative and CD10-transfected Colo320 cells in nude mice were treated with MENK and/or the CD10 inhibitor (thiorphan). Intracellular signalling of MENK and δ-opioid receptor (DOR) was examined by immunoblotting.
Results MENK inhibited the growth, invasion and survival of CRC cells following thiorphan-induced CD10 inactivation. Thiorphan suppressed liver metastasis of CD10-positive CRC cells. Inoculation of mice with CRC cells induced MENK expression in the liver. Inhibition of hepatic MENK expression by cholesterol-conjugated antisense S-oligodeoxynucleotide increased liver metastasis of CRC cells even when the cells did not express CD10. DOR activation by MENK decreased the phosphorylation of epidermal growth factor receptor and extracellular signal-regulated kinase and increased p38-dependent apoptosis. Nitric oxide was found to induce DOR expression in CRC cells. Co-treatment with thiorphan and a nitric oxide donor had a marked anti-tumour effect on liver metastasis of HT29 cells. Of 68 CRC patients, 19 (28%) showed CD10 expression, which was dependent on the extent of liver metastasis. MENK concentration in metastasis-positive human liver was higher than that in the normal liver.
Conclusion CD10 expression in CRC cells abrogates the anti-tumour effect of hepatic MENK by degrading it, which enhances liver metastasis of CD10-positive CRC cells.
Statistics from Altmetric.com
Colorectal cancer (CRC) is the fourth leading cause of cancer death in Japan, and its incidence is increasing with the progressive Westernisation of the Japanese lifestyle.1 Approximately 24% of CRCs invading beyond the submucosal layer have been shown to metastasise to the liver at the time of operation and/or after the operation.2 Liver metastasis is the cause of death in one-third of all CRC patients.3 Hence, early detection and control of liver metastasis is important for effective treatments of CRCs.
CD10, also known as common acute lymphoblastic leukaemia antigen (CALLA), is a characteristic marker of the various subgroups of B cell type acute lymphocytic leukaemia.4 5 It is a zinc-dependent membrane metalloendopeptidase, also referred to as neutral endopeptidase (EC 3.4.24.11), enkephalinase, or neprilysin.5 CD10 is expressed in CRC and is associated with CRC metastases, particularly liver metastasis.2 6 7 Logistic regression analysis has revealed that lymph node metastasis, expression of CD10, and vascular endothelial growth factor (VEGF) are the most significant factors associated with CRC metastasis.7 However, the role of CD10 in the normal intestinal mucosa and CRC is still unknown. In this study, we aimed to elucidate the role of CD10 in CRC progression.
CD10 has a wide range of substrates, including neuropeptides such as substance P, Leu-enkephalin (LENK), and Met-enkephalin (MENK).5 8 9 Opiates, including MENK, have an antiproliferative effect on normal and cancer cells.10–12 MENK is expressed in hepatocytes exposed to stresses induced by the hepatitis virus, autoimmune hepatitis, primary biliary cirrhosis, and cholestatic liver disease.13–15 It is thought to enhance the replication of the hepatitis C virus in the human liver and to interfere with the therapeutic effect of interferons.13 MENK induces hepatic fibrogenesis by activating hepatic stellate cells,16 and may also modulate T-cell function in the liver in hepatitis, and in CRCs.17 In patients with hepatitis/cirrhosis, hepatocytes expressed MENK but not the MENK-degrading CD10, whereas hepatocellular carcinoma expressed CD10.18
In this study, we tested the hypothesis that MENK produced by liver cells suppresses the metastasis of CRCs. However, MENK-induced suppression does not occur in CD10-positive CRCs because MENK is degraded by CD10. These MENK effects might accelerate liver metastasis of CRCs.
Materials and methods
Cell culture and reagents
HT29 human colon cancer cell line was purchased from Dainihon Pharmaceutical (Tokyo, Japan), and was maintained in Dulbecco's modified essential medium (Sigma, St. Louis, Missouri, USA) containing 10% fetal bovine serum (Sigma) under the conditions of 5% CO2 in air at 37°C. Met-enkephalin (MENK), p38 mitogen-activated protein kinase (MAPK) inhibitor SB239063, thiorphan, Nω-nitro-l-arginine methyl ester (l-NAME) (Sigma), extracellular signal-regulated kinase (ERK) inhibitor (Calbiochem, Darmstadt, Germany), human recombinant EGF (Serotec, Oxford, UK), and sodium nitroprusside (SNP; Sigma) were purchased.
CD10 transfectant
Colo320 human colon cancer cell line was kindly provided by Dr W Yasui (Hiroshima University). CD10 cDNA was amplified by PCR and sub-cloned into pcDNA3.1 (Invitrogen, Carlsbad, California, USA). Colo320 cells were transfected with CD10-pcDNA3.1 using Lipofectamine transfection reagent (Invitrogen). Transfectants were selected by treatment with G418 (100 µg/ml) for 5 weeks. Cells with the highest CD10 expression (Colo320-CD10) were used for further examination. Colo320 cells transfected with empty vector (Colo320-C) were also used.
Antisense phosphorothioate (S)-oligodeoxynucleotide assay
A 20-mer S-oligodeoxynucleotide (ODN) composed of the antisense sequence of nucleotides –5 to 15 (5′-TTCTGACTTGCCCATCACCT-3′) of human CD10 cDNA (GenBank BC101658) was synthesised by Sigma Genosys (Ishikari, Japan). An 18-mer antisense S-ODN for nucleotides 1 to 18 of proenkephalin (PENK; GenBank AK314908) (5′-TGTCAGGAACCGCGCCAT-3′) was also synthesised. Random 18-mers were used as negative controls. The cells were pretreated with 3 µmol l−1 antisense or sense S-ODN for 6 days, with medium exchange and addition of antisense or sense S-ODN every 2 days. The cells were then used in experiments. To inhibit MENK expression in the liver, we used cholesterol-conjugated S-ODN, which is selectively accumulated in the liver.19–21 Mouse preproenkaphalin (PPENK; GenBank NM001002927) antisense S-ODN comprising nucleotides 1 to 18 (5′-CCTCAGGAACCGCGC CAT-3′) was conjugated with cholesterol (Japan BioService, Asaka, Japan). Cholesterol-conjugated PPENK antisense S-ODN (25 µg/kg body weight) was intraperitoneally injected three times a week. Injection of cholesterol-conjugated PPENK antisense S-ODN was started 1 week before cancer cell inoculation.
Short interferent RNA
FlexiTube short interferent RNAs (siRNAs) for DOR, and epidermal growth factor receptor (EGFR) were purchased from Qiagen Genomics (Bothell, Washington, USA). AllStars Negative Control siRNA was used as a control (Qiagen). Cells were transfected with 50 nmol l−1 siRNA for each gene using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions.
Immunoblot analysis
Whole-cell lysates were prepared as described previously.22 Antibodies for CD10 (Novocastra Laboratories, Newcastle upon Tyne, UK), ERK1/2 (Chemicon International, Temecula, California, USA), phosphorylated ERK1/2, p38, phosphorylated p38, EGFR (Santa Cruz Biotechnology, Santa Cruz, California, USA), phosphorylated EGFR (phospho-Tyr992; Cell Signaling Technology, Beverly, Massachusetts, USA), MENK (Progen Biotechnic, Heidelberg, Germany), δ-opioid receptor (DOR; Chemicon), and phosphorylated DOR (phospho-Ser263; Neuromics Antibodies, Edina, Minnesota, USA) were used as primary antibodies. An anti-tubulin antibody was used as a loading control (Oncogene Research Products, Cambridge, Massachusetts, USA). The immune complex was visualised using Enhanced Chemiluminescence Western-blot detection system (Amersham, Aylesbury, UK). Densitometric scanning of immunoblot signals was performed using NIH Image software (SAS Institute, Cary, North Carolina, USA).
Cell growth, apoptosis, and in vitro invasion assay
Cells were seeded at a density of 10 000 cells per well in 12-well tissue culture plates. Cell growth was assessed by the incorporation of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (Sigma).22 Apoptosis were assessed by staining with Hoechst 33258 fluorescent dye (Wako Pure Chemical Industries, Osaka, Japan). The number of apoptotic cells was determined by examining 1000 cells. A modified Boyden chamber assay was performed to examine the in vitro invasion of colon cancer cells.22 The experiments were performed three times.
Animal model
BALB/c nu/nu athymic mice (5 weeks old) purchased from Japan SLC (Shizuoka, Japan) were used as a metastasis model. Single-cell suspensions of CRC cells (1×106) in Hanks' balanced saline solution were injected into the spleen of each mouse. Thiorphan was weekly injected into the peritoneal cavity. The mice were killed to count the number and size of metastatic foci in the liver at 4 weeks after the inoculation. The liver tissues obtained from cancer-free areas were used for MENK examination.
Surgical specimens
Formalin-fixed surgical specimens from 68 patients with CRCs that had invaded the subserosal layer were randomly selected from the Nara Medical University Hospital and the Miyoshi Central Hospital (table 1). Fresh liver tissues were obtained from 12 CRC patients with liver metastasis and 10 non-metastatic patients. The liver tissues obtained were at least 0.5 cm distant from metastatic foci. In the 12 CRCs with liver metastasis, fresh tissues were also obtained from primary tumours and liver metastatic tumours for examining MENK concentration. Because written informed consent was not obtained, identifying information for all samples was removed before analysis for strict privacy protection.
Expression of CD10 and δ opioid receptor (DOR) in specimens from 68 patients with colorectal cancer
Immunohistochemistry
Consecutive 4 µm sections were immunohistochemically stained using the immunoperoxidase technique described previously.23 Antibodies to CD10 (Novocastra) and DOR (Chemicon) were used at a concentration of 0.5 µg/ml. The specimens were colour-developed with diaminobenzidine (DAKO, Corporation, Carpinteria, California, USA) and counter-stained with Meyer's haematoxylin (Sigma). To confirm the specificity of the immunoreactivity for antibody, antibody absorbed with the corresponding antigen was used for immunostaining (absorption test). CD10 (Abcam, Cambridge, UK) and DOR (Biodesign International, Saco, Maine, USA) were used as absorbing antigens.24
After immunostaining, all the slides were assessed according to the immunoreactive score (IRS).25 This score was originally proposed for evaluation of oestrogen receptor expression, although in many studies it has also been applied for evaluation of membranous and/or cytosolic staining.26 27 For the evaluation of the staining patterns, only tumour cells were taken into consideration, thereby excluding potentially positively staining tumour cell debris. The percentage of positive cells (PP; PP1: 0–10%, PP2: 11–50%, PP3: 51–80%, PP4: 81–100%) and the staining intensity (SI; SI1: weak, SI2: moderate, SI3: strong) were determined. PP and SI values were then multiplied to give an IRS score (ranging from 0 to 12) for the staining evaluation. On the basis of the IRS score, the immunoreactivity was classified into three grades: grade 0 (IRS 0), grade 1 (IRS 1–7), and grade 2 (IRS 8–12). Cases with grades 1 and 2 were considered to be positive.
Enzyme-linked immunosorbant assay
Frozen liver tissues were sonicated in lysis buffer.22 The supernatants were used for ELISA. The concentrations of MENK and CD10 (neprilysin) were determined using a Met-enkephalin ELISA kit (Calbiochem) and a Neprilysin Human Duo Kit (R&D Systems, Minneapolis, Minnesota, USA), respectively, according to the provider's instructions.
LASER microdissection
Frozen sections were microdissected after methylene blue staining. Cancer glands in the sections were observed by microscopy. Avoiding cancer cells, the luminal spaces of the cancer glands were captured. Similarly, the stromal spaces surrounding the cancer glands were then captured with the exclusion of inflammatory cells. The target areas were captured using an Arcturus PixCell II-AS1 LASER microdissection system (Biomedical Equipment, Tokyo, Japan) according to the provider's instruction. Captured tissues were lysed with lysis buffer and the lysate was used for ELISA.
Nitrite concentration
For assessing NO production by cells, 1×106 cells were used to seed 1 ml of regular medium and treated with reagents described in the Result section for 24 h. Cultured medium was mixed with the same volume of Griess solution (1% sulfanilamide, 0.1% naphthylethylenediamine dihydrochloride, 2.5% phosphoric acid).
Statistical analysis
Statistical analyses of experimental data were performed using the Mann–Whitney U test, ANOVA, and χ2 test. Non-parametric correlation was examined by the Spearman rank correlation test. Statistical significance was defined as a two-sided p value of less than 0.05.
Results
Expression of CD10, epidermal growth factor receptor, and δ-opioid receptor in CRC cells
Human HT29-CRC cells treated with random S-oligodeoxynucleotide (S-ODN) (HT29R cells) expressed CD10; this expression was inhibited by antisense RNA against S-ODN (HT29AS cells) (figure 1a). The expression of CD10 in CD10-transfected Colo320 cells was higher than that in control vector-transfected Colo320 cells (figure 1a). HT29 and Colo320 cells also expressed the δ-opioid receptor (DOR), which is a MENK-specific receptor, and the epidermal growth factor receptor (EGFR). When MENK was added to the culture medium, it was rapidly degraded by the CD10-positive HT29R and Colo320 cells, which expressed CD10 at high levels (figure 1b). In contrast, MENK was not degraded by the CD10-negative HT29AS cells and was degraded slowly by Colo320-C cells.

Expression of CD10, EGFR and DOR in HT29 and Colo320 human CRC cells. (a) We determined the protein levels of CD10, EGFR and DOR in HT29 cells treated with CD10 antisense S-ODN and in CD10-transfected Colo320 cells. (b) Degradation of MENK added to the culture medium (40 µmol/l by CD10-positive and CD10-negative CRC cells. HT29R, random S-ODN-treated HT29 cells; HT29AS, CD10 antisense S-ODN-treated HT29 cells; Colo320-C, control vector-transfected Colo320 cells; Colo320-CD10, CD10 expression vector-transfected Colo320 cells. CRC, colorectal cancer; DOR, δ opioid receptor; EGFR, epidermal growth factor receptor; MENK, Met-enkephalin; ODN, oligodeoxynucleotide.
Effect of MENK on cell growth, apoptosis, and invasion of CD10-negative CRC cells
We examined the effect of MENK on the cell growth, apoptosis, and invasion of CD10-positive HT29 cells and CD10-negative Colo320 cells by treating the cells with the CD10-specific inhibitor thiorphan (figure 2). CD10-negative HT29AS and Colo320-C cells treated with MENK exhibited a decrease in cell number by 40–50% and increased apoptosis regardless of whether the cells were treated with thiorphan (figure 2a–d). CD10-positive HT29R and Colo320 cells treated with MENK and thiorphan exhibited a thiorphan-dependent reduction in cell number and an increase in apoptosis. An in vitro invasion assay revealed that MENK treatment markedly suppressed the invasiveness of CD10-negative cells regardless of thiorphan treatment (figure 2e,f). In the case of CD10-positive cells, MENK treatment suppressed the invasion in a thiorphan-dependent manner. Hence, MENK inhibits cell growth and invasion, and induces apoptosis in both CD10-negative and CD10-inactivated (thiorphan-treated) CRC cells.

Effect of CD10 inhibitor and MENK on HT29 and Colo320 cells. The effects of MENK and thiorphan, a CD10 inhibitor, on HT29 and Colo320 cells were analyzed on the basis of cell growth (a,b), apoptosis (c,d), and an in vitro invasion assay with a type IV collagen-coated membrane (e,f). Cells were treated with MENK (20 µmol/l) and various concentrations of thiorphan for 48 h. Error bars, SD of three independent assays.
Effect of MENK expression in the liver on CRC metastasis
We examined the effect of CD10 inactivation on liver metastasis using a nude mouse model (figure 3a,b). At 4 weeks after intraperitoneal administration of thiorphan into the mouse spleen, the number and size of the metastatic foci of CD10-positive HT29 and Colo320 cells in these mice were reduced in a dose-dependant manner compared to those in the untreated mice. The liver cells constitutively expressed MENK at very low levels in the metastasis-negative livers, whereas MENK expression was upregulated in the metastasis-positive livers (figure 3c). Hence, MENK was thought to inhibit the metastasis of CD10-inactivated CRC cells.
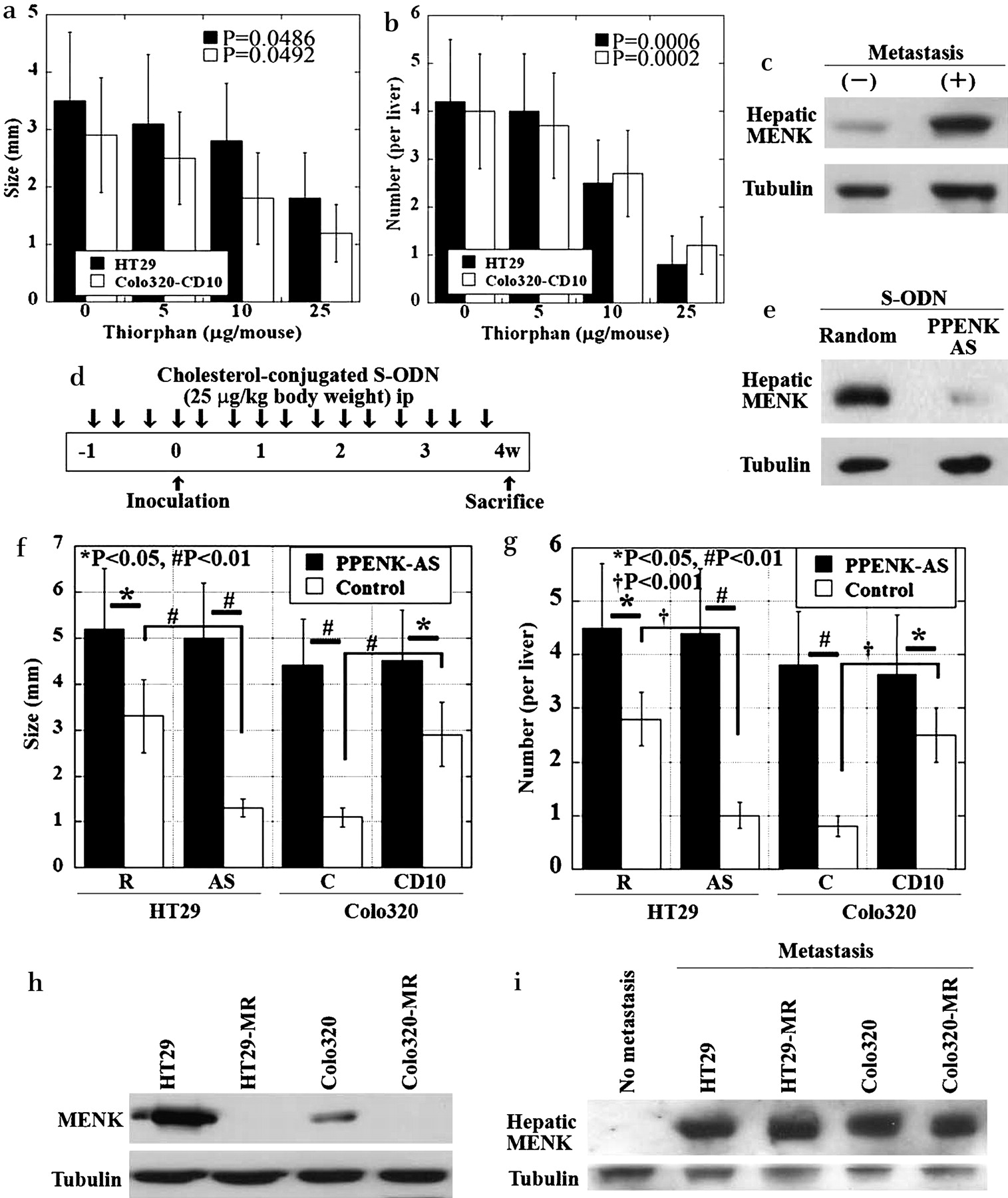
Effect of MENK production in the liver on metastasis of HT29 and Colo320 cells in nude mice. (a,b) Effect of thiorphan on liver metastasis of HT29 and Colo320 cells. The mean tumour size (a) and number (b) of metastatic foci in the liver of the nude mice were determined. (c) Immunoblotting was used to study MENK expression in livers with or without CRC metastasis. CRC cells were inoculated into the spleen, and liver metastases were examined at 4 weeks after inoculation. (d) Protocol of the cholesterol-conjugated S-ODN experiment. Cholesterol-conjugated preproenkephalin (PPENK) antisense S-ODN or cholesterol-conjugated random 20-mer S-ODN was administered intraperitoneally at a dose of 25 µg/kg body weight three times a week. (e) Effect of cholesterol-conjugated PPENK antisense S-ODN on MENK expression in the liver. Immunoblotting was used to study the MENK expression in antisense S-ODN-treated and untreated mouse livers. (f,g) Effect of MENK repression in the mouse liver by cholesterol-conjugated PPENK antisense S-ODN on liver metastasis of HT29R, HT29AS, Colo320-C, and Colo320-CD10 cells. Mean tumour size (f) and number (g) of metastatic foci in the mouse liver. PPENK-AS, PPENK antisense S-ODN treatment; Control, random S-ODN treatment. Error bars, SD of six mice. (h) MENK expression in CRC cells. HT29-MR, MENK antisense S-ODN-treated HT29 cells; Colo320-MR, MENK antisense S-ODN-treated Colo320 cells. (i) MENK expression in mouse livers with CRC cell embedding. CRC cells were inoculated into the spleen. The liver tissues were examined by immunoblotting at 2 days after inoculation. No metastasis, PBS injection into the spleen. CRC, colorectal cancer; MENK, Met-enkephalin; ODN, oligodeoxynucleotide; PPENK, preproenkaphalin.
We also examined the effect of suppression of MENK expression in the liver (figure 3d–g). Mice were injected with antisense S-ODN against cholesterol-conjugated preproenkephalin (PPENK; 25 µg/kg body weight) three times a week for 5 weeks (figure 3d). Antisense S-ODN-treated mice exhibited reduced MENK expression in the liver (figure 3e). CD10-positive cells (HT29R and Colo320-CD10) and CD10-negative cells (HT29AS and Colo320-C) were inoculated into the spleen (figure 3f,g). In livers treated with control S-ODN, the size and number of metastatic foci were significantly higher in CD10-positive cells than in CD10-negative cells. In contrast, in livers treated with PPENK antisense S-ODN, the size and number of metastatic foci of CD10-negative cells were increased to levels comparable to those of CD10-positive cells. Moreover, the size and number of metastatic foci were increased in MENK-suppressed livers by inoculation of CD10-positive cells. Thus, the suppression of hepatic MENK enhanced liver metastasis of CD10-positive CRC cells and even that of CD10-negative cells.
We confirmed the effect of cancer cell implant in the liver on hepatic CD10 expression (figure 3h). HT29 and Colo320 cells expressed MENK, which was inhibited by PENK antisense S-ODN (HT29-MR and Colo320-MR cells). To confirm the effect of cancer cell-derived MENK on hepatic MENK expression, HT29, Colo320, HT29-MR, and Colo320-MR cells were used for embedding in the liver. MENK expression was examined in the liver tissues at 2 days after inoculation (figure 3i). Non-metastatic liver tissues exhibited no MENK expression, whereas the cancer cell-embedded liver tissues exhibited MENK expression regardless of MENK expression in cancer cells. Thus, hepatic MENK expression was independent of tumoural MENK expression.
Effect of MENK on mitogen-activated protein kinase signalling in HT29 cells
The MENK receptor DOR is a G-protein-coupled receptor that can activate proteins of the mitogen-activated protein kinase (MAPK) family.28 The G-protein-coupled receptors may also modulate other receptor kinases that activate MAPKs, such as EGFR. The effect of MENK on the phosphorylation of MAPKs was investigated in MENK-repressed HT29 cells by treating the cells with antisense S-ODN against human proenkephalin (PENK) (HT29-MR cells) (figure 4a).

Effect of siRNAs on DOR- or EGFR-induced phosphorylation of MAPK proteins in HT29 cells. (a) Protein levels of EGFR, DOR, ERK1/2, and p38, and their phosphorylated forms (pEGFR, pDOR, pERK1/2, and pp38, respectively) in DOR- and EGFR-knockdown HT29 cells treated with MENK (20 µmol/l) and EGF (10 ng/ml). (b) Effects of inhibitors of p38 (p38-I), JNK (JNK-I), ERK1/2 (ERK-I), and siRNA for EGFR and DOR on the growth of HT29 cells. Error bars, SD of three independent assays. (c) Effects of inhibitors of p38 (p38-I) and DOR siRNA on the apoptosis of HT29 cells. DOR, δ opioid receptor; EGFR, epidermal growth factor receptor; ERK, extracellular singal-related kinase; JNK, cJun N-terminal kinase; MAPK, mitogen-activated protein kinase; MENK, Met-enkephalin.
Treatment with MENK and EGF induced phosphorylation of EGFR and DOR in HT29-MR cells. Knockdown of EGFR and DOR was confirmed by the absence of the EGFR and DOR proteins. DOR knockdown increased EGFR phosphorylation in the HT29-MR cells to 147% of that in the control HT29 cells, whereas EGFR knockdown increased DOR phosphorylation to 155% of that in the control HT29 cells. The protein levels of ERK1/2 and p38 were unaffected by knockdown of either EGFR or DOR. The phosphorylation of ERK1/2 increased to 210% after DOR knockdown and decreased to 38% after EGFR knockdown. The phosphorylation of p38 decreased to 44% after knockdown of DOR, but increased to 127% after knockdown of EGFR. Phosphorylation of JNK was not altered by the knockdown of either EGFR or DOR (data not shown). These data suggest that EGFR and DOR might interfere with one another's phosphorylation. Furthermore, DOR might act as a partial agonist as well as an antagonist of EGFR in ERK1/2 phosphorylation.
To evaluate the significance of DOR signals and activation of the MAPK proteins, HT29AS cells were subjected to DOR knockdown and inhibition of MAPKs by treatment with MENK and EGF (figure 4b). DOR knockdown induced higher growth (183%) in the HT29AS cells than in the HT29 control cells (p=0.0093). Inhibition of p38 increased cell growth (126%, p=0.0446), whereas inhibition of ERK1/2 reduced cell growth (p=0.0485).
To confirm the MENK-induced activation of the antiproliferative effect of p38, we counted the number of apoptotic cells among MENK-treated HT29 cells exposed to p38-inhibitor or DOR siRNA (figure 4c). DOR knockdown inhibited p38 signals and decreased the number of apoptotic cells among MENK-treated HT29 cells.
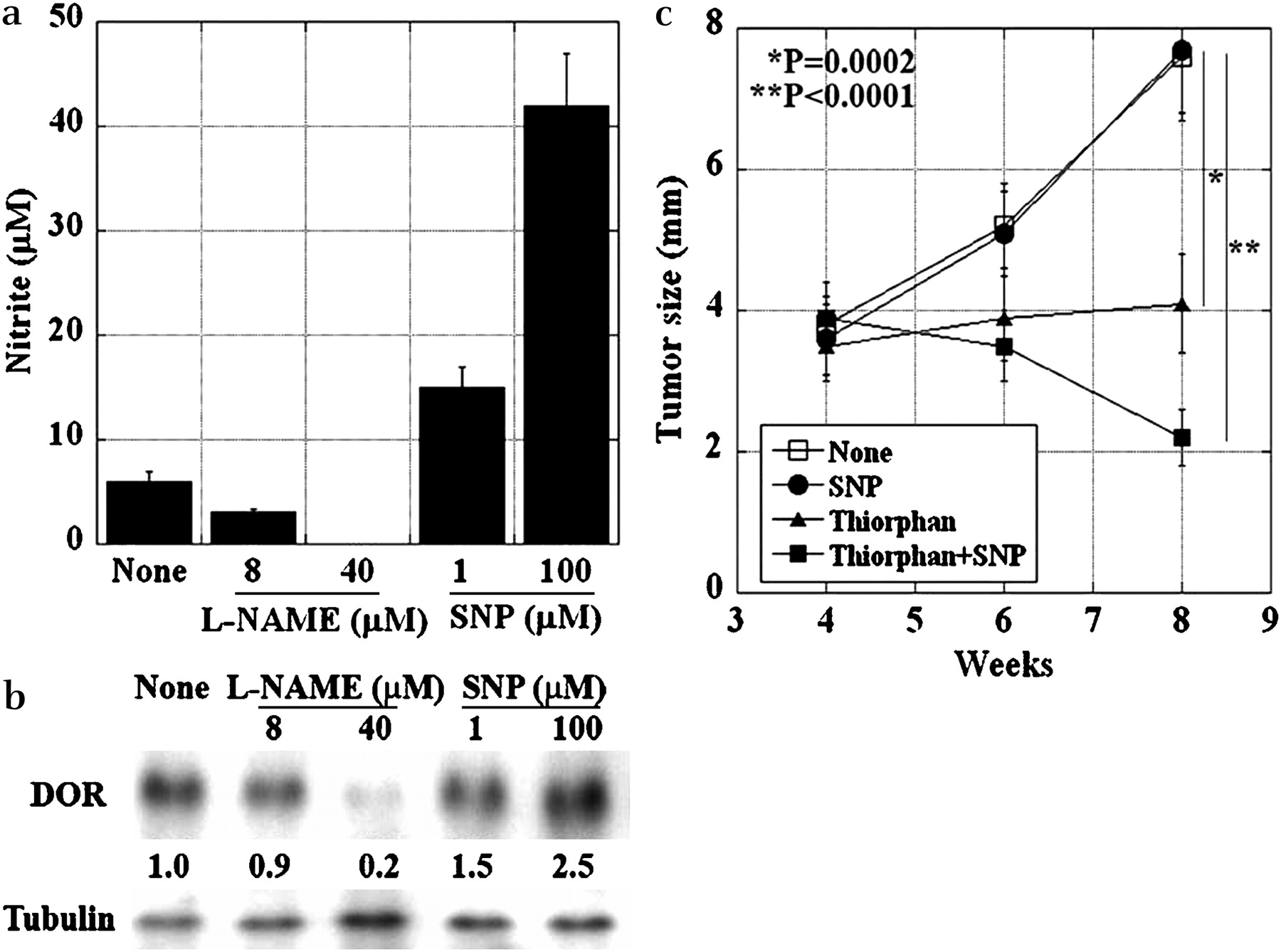
Effect of nitric oxide on DOR expression
Since DOR expression has been reported to be induced by nitric oxide (NO),29 we examined the effects of NO on DOR expression (figure 5). l-NAME treatment of HT29 cells decreased nitrite concentration in a dose-dependent manner. In contrast, SNP treatment increased the nitrite concentration dose dependently (figure 5a). Similarly, l-NAME decreased DOR expression in HT29 cells, whereas SNP increased DOR expression (figure 5b). The therapeutic application of the DOR-inducing effect of SNP was examined in the mouse liver metastasis model (figure 5c). HT29 liver tumours were established at 4 weeks after intrasplenic inoculation, and thereafter SNP (3 mg/kg, i.p.) and/or thiorphan (1 mg/kg, i.p.) were administered to the mice twice a week. At 4 weeks after the treatment, tumour growth was inhibited in thiorphan-treated mice (p=0.0002). SNP alone did not affect the tumour growth; the combination of thiorphan and SNP was the most effective method of suppressing tumour growth (p<0.0001). Thus, the anti-tumour effect of thiorphan in the liver was enhanced via DOR induction by an NO donor (SNP).

Effect of NO on DOR expression in HT29 cells. (a) NO production was assessed by determining the nitrite concentration. HT29 cells (2×104) were treated with l-NAME or SNP for 24 h. The nitrite concentration in the culture media was determined. Error bars, SD of three independent assays. (b) Immunoblotting was used to assess the DOR expression in HT29 cells treated with l-NAME or SNP. The numbers below the bands indicate signal intensities standardised by tubulin. The value of DOR expression in control cells (None) was set to 1.0. (c) The anti-tumour effect of the treatment with a combination of thiorphan and SNP on the growth of liver metastasis in HT29 cells. The mice were treated with SNP (3 mg/kg, i.p.) and/or thiorphan (1 mg/kg, i.p.) twice a week, 4 weeks after intrasplenic inoculation of HT29 cells. Error bars, SD of five mice. DOR, δ opioid receptor; l-NAME, Nω-nitro-l-arginine methyl ester; NO, nitric oxide; SNP, sodium nitroprusside.
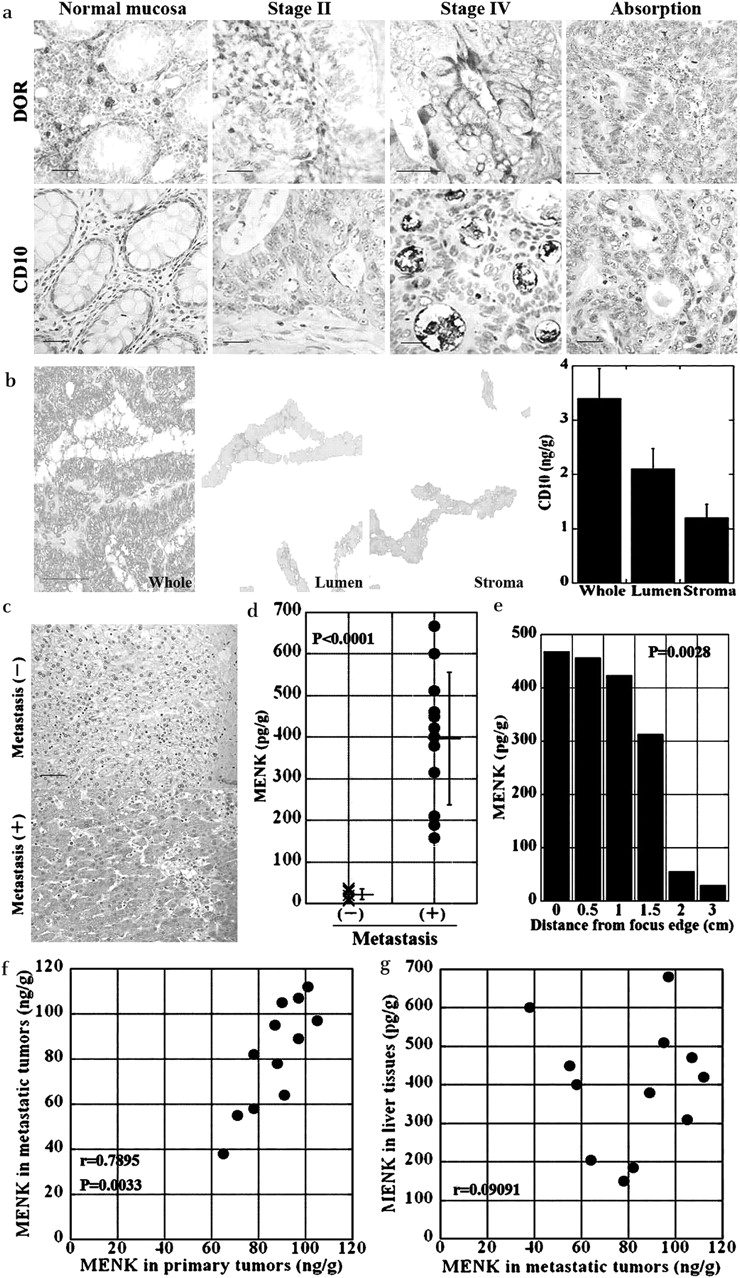
Expression of DOR and CD10 in human CRCs
We studied the expression of CD10 and DOR in the primary tumours of human CRCs by immunohistochemistry (figure 6, table 1). The luminal surface of cancer cells was found to be immunoreactive against CD10 (figure 6a), and the cytoplasm and cell membrane of cancer cells were found to be immunoreactive against DOR. No staining was observed when immunostaining was performed using the respective antigen-absorbed antibodies. CD10 and DOR were also expressed in infiltrating inflammatory cells. CD10 and DOR were expressed in 19 (28%) and 34 (50%) of the 68 patients, respectively (table 1). Expression of CD10 was associated with the TNM stage of the cancer, particularly liver metastasis. In contrast, DOR expression was not associated with any clinicopathological feature. We also examined 12 CRCs with liver metastasis to compare expression of CD10 and DOR between primary and metastatic tumours (table 2). Expression of CD10 and DOR in metastatic tumours did not differ from that in the primary tumours. Thus, the comparison of CD10 and DOR expressions failed to show any correlation with primary or metastatic tumours.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of CD10 and DOR in human CRCs and expression of MENK in the liver. (a) Expression of CD10 and DOR was examined by immunohistochemical analysis of a normal colonic mucosa (normal mucosa), a stage II case, and a stage IV case (with liver and nodal metastasis). Antigen-absorbed antibodies were used for confirming the specificity of the immunoreactivity (Absorption). Bar, 100 µm. (b) The intraluminal and stromal tissues were carefully separated from the frozen sections of colon cancer tissue by laser microdissection and were used to prepare cell lysates. Whole frozen tissue stained with methylene blue. Bar, 100 µm. Lumen, intraluminal tissues; stroma, stromal tissues. Lower right panel, CD10 concentration determined by ELISA. Error bars, SD of three independent assays. (c) Immunohistochemistry of MENK in the human liver with or without CRC metastasis. Bar, 100 µm. (d) MENK concentrations in the human liver tissues with or without CRC metastasis using 12 metastasis-positive and 10 metastasis-negative cases. The MENK concentration was determined by ELISA. Error bars, SD. (e) The relationship between MENK concentration and distance from the metastatic focus in a metastasis-positive case. The MENK concentration was determined by ELISA. Error bars, SD. (f) MENK concentration was compared between primary and metastatic tumours in 12 CRCs with liver metastasis. r: Spearman's r. (g) MENK concentration was compared between metastatic tumours and the liver tissues in 12 CRCs with liver metastasis. r: Spearman's r. CRC, colorectal cancer; DOR, δ opioid receptor; MENK, Met-enkaphalin.
Expression of CD10 and δ opioid receptor (DOR) in 12 colorectal cancers with liver metastasis
To degrade MENK, CD10 needs to be located in the stromal tissue surrounding cancer nests. To confirm the localisation of CD10, we compared CD10 concentrations in the stromal and luminal areas (figure 6b). The CD10 concentrations in the microdissected tissues of the luminal and stromal areas were 2.1 (SD 0.3) and 1.1 (SD 0.2) ng/g, respectively. Thus, localisation of CD10 was observed both at the luminal surface and in the stromal area of the cancer cells.
We also examined the MENK concentrations in the human liver tissues with and without CRC metastasis. Immunohistochemical analysis of the liver tissue adjacent to metastatic foci demonstrated MENK immunoreactivity; however, liver tissue obtained from a normal non-metastatic site did not show MENK immunoreactivity (figure 6c). MENK concentration in livers with CRC metastasis was higher than that in livers without CRC metastasis (figure 6d) (p=0.0012). In one metastasis-positive case, MENK concentrations in the liver tissue were compared with the distance from the metastatic focus (figure 6e). MENK concentration was found to be inversely correlated with the distance (p=0.0028).
Finally, we compared MENK expression among primary tumours, metastatic tumours, and the liver tissues in 12 CRCs with liver metastasis (figure 6f,g). MENK concentrations in metastatic tumours were correlated with those in primary tumours (p=0.0033), but not with those in liver tissues. However, MENK concentrations were correlated with neither the expression of CD10 nor that of DOR, in either primary or metastatic tumours (data not shown).
Discussion
CD10 is known to be associated with CRC progression and liver metastases.2 6 7 30 In the present study, we confirmed that CD10 expression is associated with the metastatic potential of HT29 and Colo320 human CRC cells.
We attempted to identify a CD10 substrate that is expressed in cancer cells in order to evaluate the role of CD10 in the progression of CRC. CD10 is a neutral endopeptidase having many substrates, including substance P, LENK, and MENK.5 8 9 Substance P is associated with development of colitis, and CD10-induced degradation of substance P completely suppresses inflammation.31 However, we found substance P had no effect on the growth and invasion of HT29 cells in vitro (data not shown). MENK inhibits tumour progression of colon cancer cells9 32 through activation of an opioid receptor.33 The normal adult liver does not express MENK; however, in primary biliary cirrhosis, viral hepatitis, and cholestasis, MENK is expressed in the liver.14 15 34 35 Therefore, we hypothesised that the expression of MENK is high in liver tissue with CRC metastasis, and that CD10 degrades MENK to abrogate the suppression of CRC metastasis. In the present study, we found that MENK directly inhibited the growth, invasion, survival, and metastasis of cancer cells, but that CD10 blocks the inhibitory effects of MENK by degrading it.
In order to study the inhibitory effects of MENK, we focused on the intracellular signalling pathway of MENK. DOR, which belongs to the family of G-protein-related receptors, is a high-affinity receptor of MENK.36 We examined the activation of MAPK proteins in HT29 cells. We found that the level of ERK phosphorylation in EGFR(+)/DOR(−) cells was higher than that in EGFR(+)/DOR(+) cells. This finding is supported by a report that suggests that signals from both EGFR and DOR are transduced via the MAPK pathway, and that DOR activation modulates EGFR activation and ERK1/2 phosphorylation.28 The finding that EGFR knockdown increased the phosphorylation of DOR and that DOR knockdown increased the phosphorylation of EGFR suggests that EGFR and DOR exhibit mutual interference. Both DOR and EGFR recruit G-proteins for intracellular signalling; hence, their signals may interfere with each other.28 The expression of DOR and EGFR has been studied in the G6 glioma cell line, and it was demonstrated that activation of DOR inhibited the subsequent activation of EGFR, whereas activation of EGFR did not inhibit the activation of DOR.37 Consequently, the findings of the present study regarding the relationship between DOR and EGFR require further investigation.
Surprisingly, DOR, which has an anti-tumoural effect, is found to be expressed in one-third of all patients with CRC. In response to NO, DOR expression is upregulated in the mucosa in inflammatory bowel disease.29 38 NO, which is expressed at high levels in CRC tissues, is a bifunctional biomodulator and induces cell injury and influences cell survival.39 NO induces the expression of BCL-2 and VEGF; however, CRC cells are resistant toward NO owing to over-expression of haem oxygenase-1.40 Our data showed that the inhibition of inducible NO synthase decreased DOR expression in both HT29 and Colo320 cells. Nuclear factor kappa B (NF-κB) and Sp-1 also induce the transcription of DOR,41 42 which is over-expressed in HT29 and Colo320 cells (data not shown). These common factors (NO, Sp-1, NF-κB), which are found in CRCs, are thought to be associated with DOR expression in CRCs.
Our data suggested that MENK induction in the liver tissue might be a response to CRC metastasis. Metastasis and embedding of CRC cells induced MENK expression regardless of MENK expression in CRC cells. In human material, MENK concentration in liver tissues was not correlated with that in metastatic tumours. A volume effect of metastasis resulting in local ischaemia, hypoxia and cholestasis might be one possible factor contributing to MENK induction. Bioactive substances produced by cancer cells might be associated with the induction of MENK.
In the present study, we found that CD10 degrades the MENK produced in livers with metastasis, which suppresses tumour cells through DOR activation. Liver metastasis is more commonly observed in CD10-expressing CRC cells, and MENK is produced in response to such metastasis. Our results suggest that CD10 is a potential target for CRC treatment. We investigated the anti-metastatic effect of a CD10 inhibitor, namely, thiorphan, by systemic administration of thiorphan in mice. The anti-tumoural effects of thiorphan on Lewis lung carcinoma metastases have been previously reported.43 Moreover, we confirmed that an NO donor and CD10 inhibitor had a synergic effect, inducing the activity of DOR. In the future, it will be necessary to evaluate the efficacy of administering a CD10 inhibitor with chemotherapeutic agents containing NO donors for preventing and treating liver metastasis following colon cancer treatment.
References
Footnotes
Funding This work was supported in part by Grant-in-Aid for Scientific Research from Ministry of Health, Labour and Welfare, Japan.
Competing interests None.
Ethics approval The procedure regarding the human tissue used in this study was in accordance with the Ethical Guidelines for Human Genome/Gene Research enacted by the Japanese Government. The mice used in this study were maintained according to the institutional guidelines approved by the Committee for Animal Experimentation of Nara Medical University.
Provenance and peer review Not commissioned; externally peer reviewed.