


Abstract
Background/Aim: Advanced renal cancer still has a very poor prognosis. In this regard recent investigations demonstrated a constitutive activation of the Wnt signaling pathway in renal cell carcinoma (RCC) thereby promoting an exaggerated cell proliferation. Especially, β-catenin overactivation and the functional loss of endogenous Wnt antagonists are associated with RCC carcinogenesis and progression. Thus, influencing Wnt signaling might represent a promising target in RCC treatment. Materials and Methods: It was recently confirmed that ethacrynic acid (EA), ciclopirox olamine (CIC) and piroctone olamine (PO) can inhibit Wnt signaling in various cancer cell lines. Herein we investigated their cytotoxic potential towards human RCC cells and their influence on the Wnt pathway concerning apoptosis as determined by 3,3’-dihexyloxacarbocyanine iodide (DiOC6) and propidium iodide (PI) staining in flow cytometry and immunoblotting. Results: All three agents, EA, CIC and PO triggered a significant apoptotic activity in tested RCC cell lines in a time- and concentration-dependent manner. Moreover, exposure to CIC and PO decreased the expression of β-catenin as the pivotal feature within the canonical Wnt pathway. However, β-catenin expression increased upon the treatment with EA. Conclusion: These results reveal a significant selective induction of apoptosis by EA, CIC and PO and suggest a suppression of RCC survival in part due to inhibition of Wnt/β-catenin signaling. The development of targeted-therapies affecting the Wnt signaling pathway might therefore lead to novel treatment options for RCC patients.
Renal cell carcinoma (RCC) represents the most common neoplasm in adult kidney accounting for approximately 3-5% of all adult malignancies in the United States, with an increasing incidence over the last decades (1). An early diagnosis of RCC is associated with favorable prognosis. Unfortunately, up to 30% of patients with RCC are diagnosed at an advanced metastatic stage (2) since typical clinical symptoms like flank pain, mass and hematuria turned-out to be ineffective for early diagnosis (3). At present, treatment is characterized by a multi-modal strategy, combining systemic therapy with surgical nephrectomy (4). Recent advances in understanding the molecular tumor biology in RCC led to the establishment of new therapeutic regimens including targeted agents such as sorafenib, sunitinib and temsirolimus, to name a few. Thereby outcome and patient survival could be additionally improved. Despite, however, this major progress, RCC eventually remains an incurable disease with a 5-year relative survival rate of ~9% in advanced renal cancer (2), still underlining the urgent need to investigate new therapeutic approaches.
The Wnt/beta-catenin pathway turned-out to be a promising target as it is involved in the regulation of cell proliferation, differentiation and apoptosis induction. In several types of cancers, including RCC, an enhanced activation of the Wnt signaling pathway is a major etiological factor (5-7). As an important downstream effector, β-catenin is the key player in the canonical Wnt signaling pathway. When Wnt signaling is in its resting state, cytosolic β-catenin forms a destruction complex composed of axin, adenomatous polyposis coli (APC), casein kinase (CK) and glycogen synthase kinase-3 β(GSK-3 β), phosphorylating β-catenin. This phosphorylated β-catenin is ubiquitinated by cellular β-transducin repeat-containing proteins (β-TrCP) and subsequently degraded by the proteasome (8). In the canonical Wnt-pathway, binding of Wnt-ligands to Frizzled (Fzd) receptors and the co-receptor low density lipoprotein receptor-related protein 5 (LRP5) or LRP6 results in the phosphorylation of the cytoplasmic adaptor protein disheveled (Dvl), leading to inhibition of glycogen synthase kinase (GSK-3 β) activity and thereby promoting a de-phosphorylation and stabilization of β-catenin. Unphosphorylated β-catenin is enabled to accumulate within the nucleus. Its interaction with members of the lymphoid enhancer-binding factor (LEF) and T-cell factor (TCF) family (7, 9) promotes the transcription of critical Wnt target genes governing cell proliferation such as c-myc and cyclin D1. Therefore, influencing the Wnt signaling pathway could lead to novel targeted strategies in cancer therapy.
It was recently confirmed that the commonly used diuretic agent ethacrynic acid (EA), the anti-fungal agent ciclopirox olamine (CIC) and the pyridone derivate piroctone olamine (PO) inhibit the Wnt/beta-catenin pathway and may be effective agents in the therapy of various, especially hematopoietic types of cancer (10-13). Similar results were obtained with other newly-discovered biological Wnt inhibitors as well, corroborating an efficacy in apoptosis induction in cancer cells due to alterations in Wnt-mediated protein expression (14). Additionally, in our previous studies Koller et al. first revealed an exceptional impact of Wnt inhibitors in renal cancer when used in combination with the bi-functional peptide TP-tox (15). For this reason, we investigated the cytotoxic effects of EA, CIC and PO as single agents and also their potential influence on the Wnt/beta-catenin pathway in RCC. We demonstrate that all tested substances exhibit selective toxicity and trigger apoptosis induction in RCC cells. In addition, they all had an influence on the β-catenin expression and could therefore prevent a constitutive activation of Wnt signaling.
Materials and Methods
Cell lines and culture conditions. All cell lines were obtained from ATCC (LGC Standards, Wesel, Germany), Cell Line Service (Eppenheim, Germany) or DSMZ (Braunschweig, Germany) and incubated at 37°C with 5% CO2 at 90% humidity. The human kidney cancer cell line A704 was cultured in Dulbecco's Modified Eagle Medium (DMEM; Invitrogen, Darmstadt, Germany), supplemented with 15% heat-inactivated fetal calf serum (FCS) (Invitrogen), 1% penicillin/streptomycin (Seromed, Jülich, Germany) and 0,2% insulin. Human kidney cancer cell lines A498 and Caki-2 were cultured in Roswell Park Memorial Institute (RPMI)-1640 medium (PAA, Pasching, Austria), supplemented with 2.5% FCS and 1% penicillin/streptomycin. The human colon fibroblast cell line CCD-18Co was cultured in ATCC-formulated Eagle's minimum essential medium (EMEM; LCG Standards, Wesel, Germany) supplemented with 10% FCS and 1% penicillin/streptomycin. The medium was renewed every 3 days and the cells were harvested using a 0.05% trypsin-EDTA solution (Invitrogen), centrifugated at 1200 rpm for 7 minutes and then resuspended in 1mL medium to define cell count.
Human samples. Peripheral blood lymphocytes (PBLs) were isolated from blood samples of healthy volunteers using Ficoll density gradient centrifugation (Lymphoprep, Nycomed, Oslo, Norway). Blood from buffy coats was diluted 1:2 with phosphate buffered saline (PBS)/1% bovine serum albumin (BSA) (both from PAA) and used for a ficoll gradient (Lymphoprep). The leukocyte layer was transferred to new tubes after centrifugation at 800 ×g for 30 min. Cells were washed three times with PBS/1%BSA and resuspended in RPMI-1640 medium supplemented with 10% FCS, 1% penicillin/streptomycin and 2.5% 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) buffer solution (PAA). Ethics approval had been obtained according to the guidelines of the host Institution and all samples were taken after signed informed consent.
Drugs and chemical reagents. The following drugs were used in this study: EA, CIC and PO. EA and CIC were purchased from Sigma-Aldrich (Steinheim, Germany), and PO was obtained from Spinnrad (Bonn, Germany).
For the flow cytometry experiments, the drugs were tested at different concentrations with incubation times of 24, 48 and 72 h. EA and CIC were used at concentrations ranging from 10-200 μM and PO was used at a concentration range of 1-50 μM on A-498, A-704 and Caki-2 cells. On PBLs, EA and CIC were used at concentrations ranging from 0.1-200 μM while PO was administered at a concentration range of 0.1-66 μM with an exposure time of 72 h.
3,3’-dihexyloxacarbocyanine iodide (DiOC6) and Propidium iodide (PI)-staining. We assessed mitochondrial transmembrane potential which occurs late in the apoptotic process by using DiOC6 staining and flow cytometry. Therefore, 1×105 cells were plated in 3-ml medium in 6-well plates. EA, CIC and PO were dissolved in Dimethyl sulfoxide (DMSO; Invitrogen) and added to the medium in different concentrations for 1-3 days as described above. For the apoptosis assay, staining with DiOC6 for detecting mitochondrial membrane potential in viable cells and with propidium iodide (PI) that binds to DNA in necrotic cells was used. Measurements were performed on a fluorescence-activated cell sorter (BD FACSCanto, Franklin Lakes, New Jersey, USA). The medium, containing drug-treated cells, was transferred from each well into a glass tube. Cells were then centrifuged at 800 ×g for 7 min, washed with PBS (pH 7.4) (Roti-Stock 10×; CarlRoth, Karlsruhe, Germany) and stained after repeated centrifugation by adding 500 μl staining solution (RPMI-1640, 0.5% BSA, 80 nM DiOC6) for 15 min at 37°C. After an additional washing step with PBS/1% BSA, cells were resuspended in 500 μl PBS/1% BSA. FACS analysis followed immediately after the addition of 5 μl PI solution (100 μg/ml) with a BD FACSCanto flow cytometer. Approximately 10,000 counts were made for each tested sample. In our experiments viable cells show high fluorescence intensity for DiOC6 and low expression for PI. On the contrary, necrotic cells exhibit high fluorescence intensity for PI and low for DiOC6. Apoptotic cells show a low expression for both DiOC6 and PI. Cells with high fluorescence intensity for both, DiOC6 and PI, however, may be derived from either cell debris or apoptotic bodies.
Immunoblotting. Cells were treated with EA, CIC and PO at concentrations of 150, 100 and 50 μM, respectively, for 24 h. Therefore, more than 1×106 cells were collected by centrifugation at 800 g for 7 min. Cells were washed twice with ice-cold PBS, transferred in a centrifuge tube, and resuspended in 1 ml lysis buffer (25 mM Tris–HCl; pH7.6), 150mM NaCl, 1% NP-40, 1% sodium deoxycholate and 0.1% sodium dodecyclsulfate (SDS), all purchased from ThermoScientific, Bonn, Germany). This suspension was then placed on ice for 45min interrupted by occasional vortexing and finally centrifuged at 13,000 g for 20 min at 4°C. The supernatant was stored at −80°C. Protein concentrations were determined using a detergent compatible protein assay (Bio-Rad Laboratories, Munich, Germany). Samples containing 80 μg of protein were boiled in SDS sample buffer (Roti-Load 1, Carl Roth, Karlsruhe, Germany), loaded on 10% SDS/PAGE and subsequently blotted onto polyvinylidene difluoride (PVDF) membranes (Immobilon-P, EMD Millipore Corporation, Billerica, MA, USA). Membranes containing transferred protein were blocked by incubating the membrane in blocking solution (Roti-Block) for 30 min followed by immunoblotting using anti-β-catenin antibody (1:1000) and anti-β-actin antibody (1:1000) (all antibodies purchased from Santa Cruz Biotechnology, CA, USA) overnight at 4°C. Appropriate horseradish peroxidase-conjugated anti-IgG was used as secondary antibody (Santa Cruz Biotechnology. Membranes were developed using a chemiluminescence system (Luminata forte, EMD Millipore Corporation, Billerica, MA, USA).
IC50 concentrations of EA, CIC and PO on renal cell carcinoma cell lines.
Statistical analysis. The Student's t-test was used for statistical analysis and values are given as mean±standard deviation (SD). Different sample sizes (n) were chosen for different cell lines as mentioned in the respective description. A p-value of less than 0.05 was considered significant.
Results
Titration of EA, CIC and PO. We showed that all three drug agents used exhibit selective toxicity to a wide spectrum of cancers. However, the obtained effects were dependent on the respective cell line and concentration administered. Therefore, as a first step, we determined the optimal concentrations of EA, CIC and PO which led to a significant decrease in viability of RCC cells. As control, human PBLs and CCD-18Co colon fibroblasts were investigated by FACS analysis. The mean 50% inhibitory concentration (IC50) after 72 h was detected by titration. IC50 values of each single agent employed after 72 h of incubation are shown in Table I.

Effect of EA on viability of A-498 (A), A-704 (B) and Caki-2 (C) RCC cells. Cells were cultured with different compounds for one to three days. Viability was measured by DiOC6 and PI staining using flow cytometry. Results represent data from three separate experiments. Data are shown as mean±SD.
Effect of EA on viability of RCC cells and control cell lines. Unlike our findings in hematopoietic malignancies, the application of EA used in concentrations less than 100 μM did not significantly influence the viability of RCC cells in vitro. However, administered concentrations ranging from 100-200 μM significantly decreased the viability of RCC cells in a time- and concentration-dependent manner. Maximum efficacy occurred after an incubation time of 72 h (Figure 1). In contrast, the effect of EA used at concentration levels higher than 100 μM was more toxic towards normal PBLs derived from healthy volunteers than compared with renal cancer cells (Table I). After 24 h, the IC50 for CCD-18Co colon fibroblasts was determined as 45 μM (Table I). Figure 4 (Panel A) shows the corresponding flow cytometry results.
Effect of CIC on viability of RCC cells and control cell lines. CIC showed a cytotoxic effect on all tested cell lines. For A-498 and Caki-2 cells, the application of CIC caused a significant decrease in viability at concentrations starting from 10 μM after 24 h. A-704 cells, however, required higher doses for significant apoptosis induction. The observed decrease in viability was time- and concentration-dependent (Figure 2). Interestingly, the concentrations concerning RCC cells had merely marginal toxic effects on PBLs and CCD-18Co colon fibroblasts that served as controls (Table I). Figure 4B presents the respective flow cytometry results.
Effect of PO on viability of RCC cells and control cell lines. Exposure to PO also strongly decreased RCC cells' viability and triggered a significant selective induction of apoptosis in all tested cell lines. To note that after 48 h, incubation with 10 μM of PO was sufficient to induce apoptosis. This effect was more obvious 72 h post-exposure (Figure 3). Healthy PBLs and CCD-18Co colon fibroblasts, however, were only marginally affected by PO when treated with the same concentrations administered to RCC cells (Table I). Exemplary flow cytometric results are shown in Figure 4C.
Effect of EA, CIC and PO on the Wnt/beta-catenin pathway in RCC. As a final step, we investigated the effect of EA, CIC and PO on the Wnt/beta-catenin pathway leading to β-catenin's accumulation in the cytoplasm and its eventual translocation into the nucleus, an important feature within the Wnt pathway. β-catenin levels were analyzed after incubation of 24 h. After exposure to EA, β-catenin expression was up-regulated in all tested RCC cell lines. When exposed to CIC, β-catenin levels decreased in A-498 and A-704 cells while there was no noticeable alteration in Caki-2 cells. The use of PO, however, strongly impaired the β-catenin expression in all RCC cells employed as evidenced from the Western blot experiments performed (Figure 5).
Discussion
Renal cell carcinoma, due to its resistance to chemotherapy and radiation therapy, is the most lethal type of genitourinary cancer. As yet, surgical nephrectomy of the tumor remains the only way for curative therapy. A number of drugs developed in recent years have shown limited-to-significant efficacy in treating RCC (1-4).

Effect of CIC on viability of A-498 (A), A-704 (B) and Caki-2 (C) RCC cells. Cells were cultured with different compounds for one to three days. Viability was measured by DiOC6 and PI staining using flow cytometry. Results represent data from three separate experiments. Data are shown as mean±SD.
Besides Von Hippel-Lindau (VHL), Vascular endothelial growth factor (VEGF) and mammalian target of rapamycin (mTOR) signaling, additional pathways, in particular Wnt/β-catenin, have been implicated to be decisive factors in RCC pathogenesis (16).
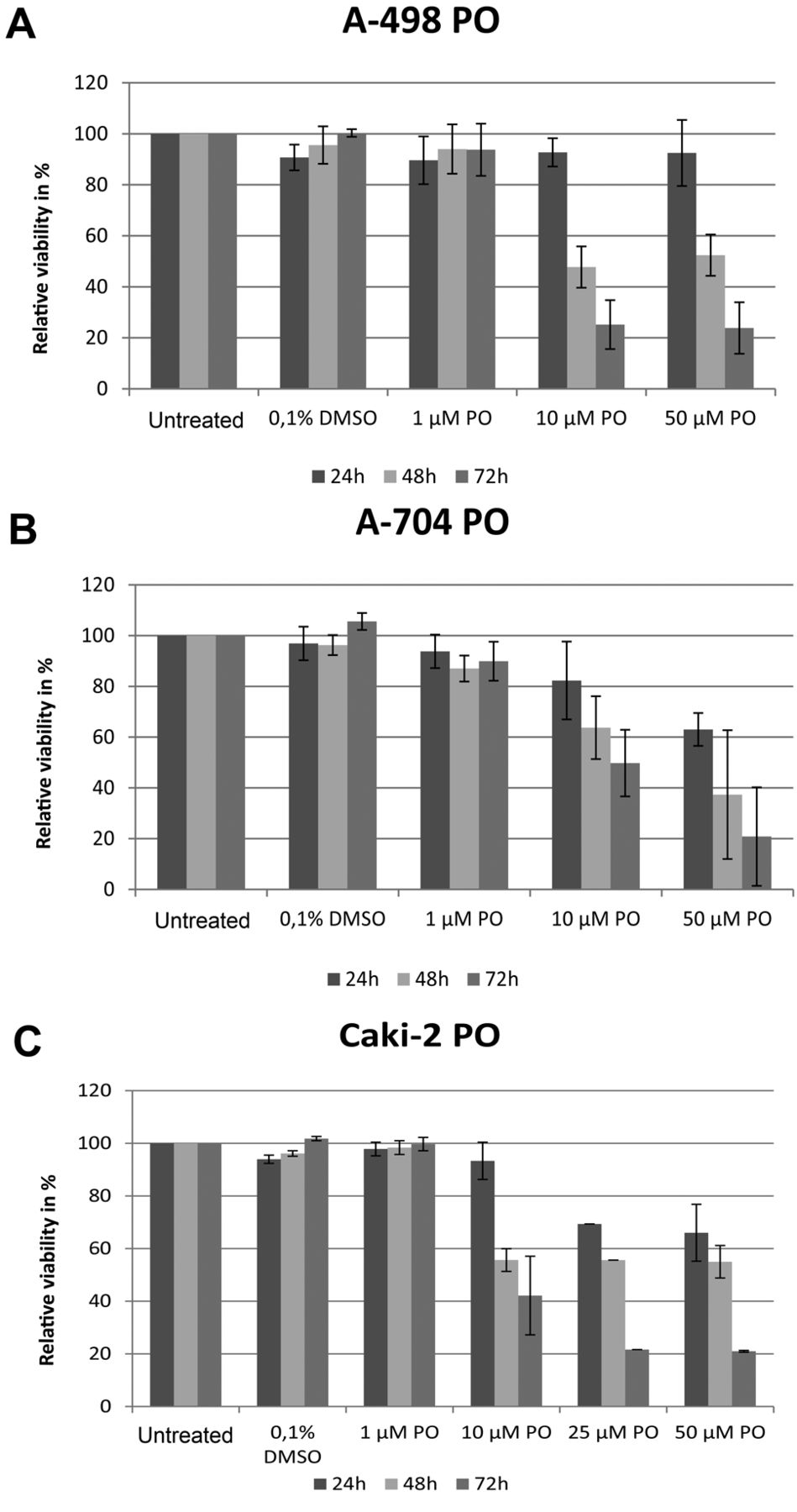
Effect of PO on viability of A-498 (A), A-704 (B) and Caki-2 (C) RCC cells. Cells were cultured with different compounds for one to three days. Viability was measured by DiOC6 and PI staining using flow cytometry. Results represent data from three separate experiments. Data are shown as mean±SD.
WNT/β-catenin signaling dysregulation, especially β-catenin overactivation and the functional loss of endogenous Wnt antagonists conduce to constitutive activation of the Wnt pathway and are associated with RCC carcinogenesis and progression through exaggerated cell proliferation and differentiation (17, 18). Most interestingly in this regard, Kruck et al. postulated the cytoplasmic β-catenin fraction as the most promising candidate associated with unfavorable clinicopathology and impaired survival in RCC (19). Hence, it follows that inhibition of Wnt/β-catenin signaling suppresses RCC growth (17). Thus, the Wnt signaling pathway emerged as an attractive therapeutic target for RCC.
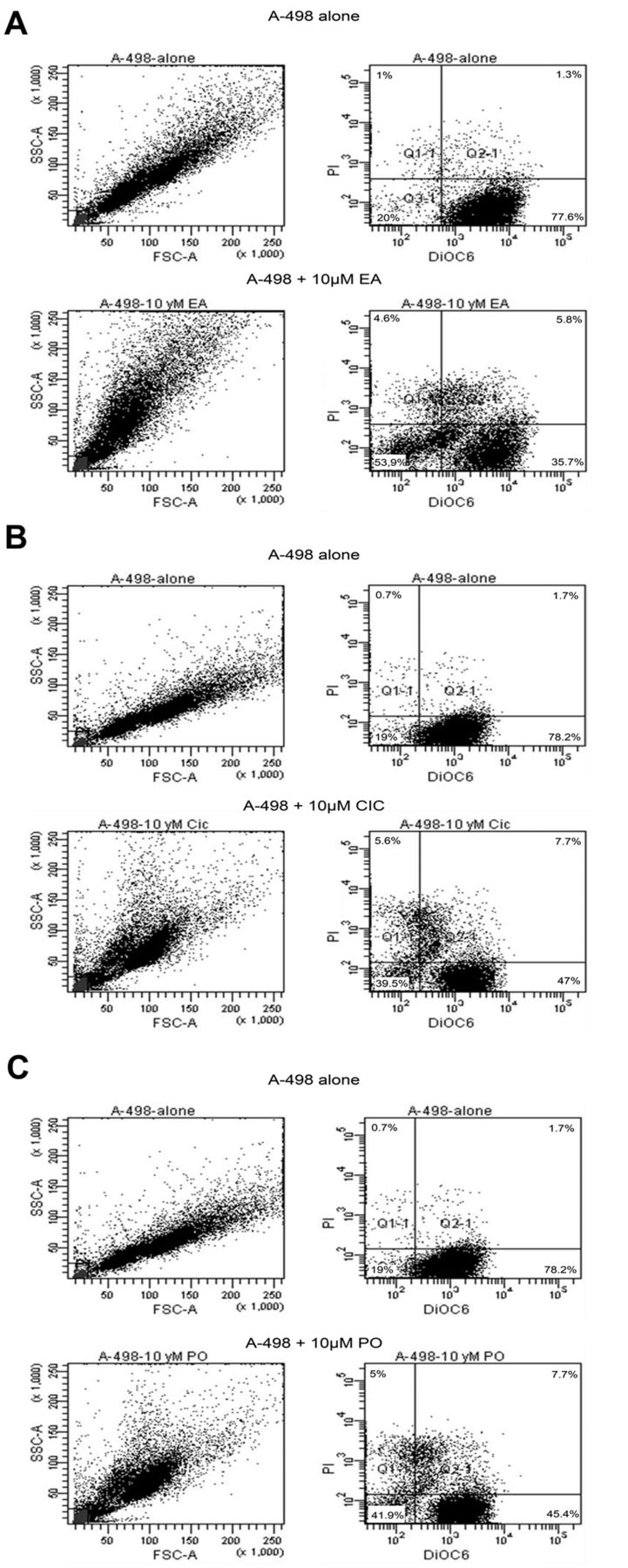
Exemplary results generated by flow cytometry. Within the quarters the relative number of cells is given in percentages. A-498 cells were treated with EA, CIC and PO each at a concentration of 10 μM. Seventy-two hours after incubation, flow cytometry was performed. (A) Cells before and after treatment with EA. (B) Cells before and after treatment with CIC. (C) Cells before and after treatment with PO.

Western blot analysis was performed under untreated conditions and under treatment with either EA, CIC or PO for 24 h. EA was used at a concentration of 150 μM for A-498, A-704 and Caki-2 cells. CIC was used at a concentration of 10 μM for A-498, A-704 and Caki-2 cells while PO was administered on A-498, A-704 and Caki-2 cells at a concentration of 50 μM. After cells were lysed, western blot analysis was performed by using 80 μg of whole-cell lysates for detecting β-catenin. α-Actin immunoblotting served as the loading control.
Recently, our workgroup revealed three drugs, EA, CIC and PO to be efficient inducers of apoptosis in lymphoma and myeloma cells in vitro. In vivo, tumor growth was significantly reduced and overall survival was prolonged in myeloma-bearing mice. All three drugs rendered the tested cell lines more sensitive to other agents and influenced the Wnt pathway through targeting either β-catenin itself or its downstream factors (10, 11, 13, 20, 21). These promising effects on both cancer cell survival and Wnt signaling encouraged us to determine whether EA, CIC or PO display cytotoxicity towards RCC cells.
EA, commonly used as loop diuretic agent, revealed a variety of interesting properties besides inhibition of the Na+-K+-2Cl− kidney import. Several studies showed EA's cytotoxicity towards different cancer cells while, additionally, EA led to an increase of other drugs' cytotoxicity (11, 22). The inhibition of glutathione-S-transferase (GST), causing increased cellular oxidative stress through higher glutathione (GSH) levels, was supposed to be the biological effect by which EA induces apoptosis. However, this finding, i.e., independence between GSH levels and EA-related induction of apoptosis, has not yet been confirmed by other investigators. N-acetyl-L-cystein (NAC) could protect cancer cells from apoptosis upon treatment with EA while GSH levels remained elevated (23). EA has also been characterized as an inhibitor of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and mitogen-activated protein kinase (MAPK) signaling (23, 24).
In the present study, EA decreased the viability of all cultured RCC cell lines in a dose- as well as time-dependent manner, by probably influencing the Wnt/β-catenin pathway as β-catenin expression was up-regulated in all tested RCC cells after exposure to EA. In a previous study we demonstrated, that EA could influence the Wnt signaling pathway by interacting with the LEF-1 protein and subsequently inducing a de-stabilization of the LEF-1/β-catenin complex. The expression of the Wnt target genes cyclin-D1, LEF-1 and fibronectin was also diminished (21). Based on our observations, it is tempting to speculate that β-catenin accumulates before its degradation in the proteasome due to an EA-mediated inhibition downstream β-catenin. The observed cytotoxicity on healthy PBLs was surprising as EA is in common clinical use. The doses we used, however, might have been too high and this is a point necessitating further experimentation.
The second drug under study, CIC, is a synthetic anti-fungal agent, usually applied to treat mycoses. CIC recently revealed many-faceted anti-carcinogenic properties (10, 11, 25-28). However, relatively little is known so far about the exact mechanisms of CIC's antitumor effects. Iron-chelation has been considered to inhibit iron-dependent enzymes like ribonucleotide reductase (26). In addition, CIC has been proven able to slow down the cell-cycle progression from G1/G0 to S phase in tumor cells and induced apoptosis (27). Recent investigations, including our previous studies, have demonstrated a down-regulation of Wnt signaling in colon, breast cancer, lymphoma and myeloma cell lines upon treatment with iron chelators such as CIC (10, 11, 28, 29).
Herein, we confirm on the anti-proliferative effects of CIC and demonstrated its potent selective cytoxicity in RCC cells. As it resulted from a flow cytometry assay, viability was significantly decreased in all tested RCC cells, and apoptosis induction was triggered in a time- and concentration-dependent manner. From similar findings in other cancer cells, the present study also indicates an influence of CIC on the Wnt signaling pathway in RCC by altered levels of β-catenin as the key protein within the Wnt pathway.
PO, is a pyridine derivate, as the case with CIC is, which was first synthetized by Hoechst AG, Frankfurt/ Main, Germany and is widely found in many cosmetic products such as anti-dandruff shampoos and hair tonics. Additionally, PO exhibits fungicidal and bactericidal effects on gram-positive as well as gram-negative bacteria. After penetrating the cell's membrane, iron(III)-chelation has been supposed to be the underlying mechanism by which PO inhibits iron-dependent mitochondrial enzymes (30, 31). Moreover, we have recently pointed-out PO's capacity to induce apoptosis in lymphoma and myeloma cells and reduce tumor growth in myeloma-bearing mice (13). The present data support our previous findings about PO's influence on Wnt signaling in the RCC experimental model where β-catenin levels decrease in all RCC cell lines upon treatment thus suggesting an effect on β-catenin itself or its upstream factors. Of note, PO displayed a significant induction of apoptosis in all tested RCC cells.
We showed that all three agents under investigation trigger a significant selective induction of apoptosis in RCC cell lines without exceedingly harming healthy PBLs, except for the case of EA. Moreover, β-catenin expression was altered in all tested cells. These experiments suggest that EA, CIC and PO selectively suppress RCC survival in part due to inhibition of Wnt/β-catenin signaling. Nevertheless, multiple effects due to an interaction of Wnt-antagonism and other associated signaling pathways are entirely conceivable and thus the elevated sensitivity of RCC cells vis-a-vis the agents employed may be attributed to their potential influence on both Wnt and other signaling cascades. In particular, the EA-mediated inhibition of NF-κB and MAPK signaling might, besides a destabilization of the LEF-1/β-catenin transcription complex, additionally contribute to its anti-proliferative properties. In the case of CIC and PO, the precise action mechanisms leading to cancer cell death and impaired Wnt expression, remain unclear. Therefore, further experimentation is required in order to delineate the exact pathways associated with the various patterns defining cellular survival.
Acknowledgements
The technical expertise of Petra Alpmann and Sabine Blaum-Feder is kindly acknowledged.
Footnotes
-
↵* These Authors contributed equally to this study.
- Received April 14, 2014.
- Revision received June 11, 2014.
- Accepted June 12, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Guanylate-binding Protein 2 Expression Is Associated With Poor Survival and Malignancy in Clear-cell Renal Cell Carcinoma
- In Vitro Apoptosis Induction by Fenofibrate in Lymphoma and Multiple Myeloma
- Griseofulvin Efficiently Induces Apoptosis in In Vitro Treatment of Lymphoma and Multiple Myeloma
- Clofibrate Demonstrates Efficacy in In Vitro Treatment of Lymphoma and Multiple Myeloma
- In Vitro Efficacy of Naftifine Against Lymphoma and Multiple Myeloma
- Flunarizine Exhibits In Vitro Efficacy Against Lymphoma and Multiple Myeloma Cells
- In Vitro Efficacy of Cinnarizine Against Lymphoma and Multiple Myeloma