


Abstract
Quercetin, a principal flavanoid compound in onions, has been shown to possess a wide spectrum of pharmacological properties, including anticancer activities. Our earlier study showed that quercetin induced cytotoxic effects on SAS human oral cancer cells. In this study, we found that quercetin significantly reduced wound closure of SAS cells in culture plates after 12- and 24-h treatments. Results indicated that quercetin inhibited the expression and activity of matrix metalloproteinase (MMP)-2 and MMP-9, as measured by western blotting and gelatin zymography. The results from western blotting also showed that quercetin reduced the protein levels of MMP-2, -7, -9 and -10, vascular endothelial growth factor (VEGF), nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) p65, inductible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), urokinase-type plasminogen activator (uPA), phosphatidylinositide-3 kinases (PI3K), nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha (IKBα), IKB-α/β, phosphorylated nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor kinase, alpha/beta (p-IKKα/β), focal adhesion kinase (FAK), son of sevenless homolog-1 (SOS1), growth factor receptor-bound protein-2 (GRB2), mitogen-activated protein kinase kinase kinase-3 (MEKK3), MEKK7, extracellular-signal-regulated kinase 1/2 (ERK1/2), p-ERK1/2, c-Jun N-terminal kinase 1/2 (JNK1/2), p38, p-p38, Jun proto-oncogene (c-JUN) and p-c-JUN but it did not affect Ras homolog gene family, member A (RhoA), Protein kinase C (PKC) and rat sarcoma viral oncogene homolog (RAS) in SAS cells. Confocal laser microscopy also showed that quercetin promoted the expressions of RhoA and Rho-associated, coiled-coil containing protein kinase-1 (ROCK1), but inhibited the expression of NF-κB p65 in SAS cells. It is concluded from these data that inhibition of migration and invasion of SAS cells by quercetin is associated with the down-regulation of PKC and RhoA by blocking MAPK and PI3K/AKT signaling pathways and NF-κB and uPA, resulting in inhibition of MMP-2 and MMP-9 signaling.
In Taiwan, approximately 10 individuals per 100,000 die annually from oral cancer, which it is the fourth most common type of cancer and the sixth cause of death in the entire population, based on the 2010 report from the Department of Health, R.O.C. (Taiwan) (1). Factors related to oral cancer include cigarette smoking, chewing of betel quid, and alcohol consumption (2). Betel quid chewing is one of the major factors in Taiwan (3). Current treatments (surgery, radiotherapy and chemotherapy) for oral cancer are still unsatisfactory (4); furthermore, oral cancer also metastasizes and causes cancer in other organs or tissues of patients (5).
Metastasis involves cytoskeletal proteins, and changed cell adhesion ability may influence cell motility, including invasion and migration of tumors cells (6, 7). It is well-documented that when cells metastasize, matrix metalloproteinases (MMPs) and the urokinase plasminogen activator (uPA), associated with cell invasion and migration, are overexpressed (8, 9). Therefore, it was suggested that use of an agent to inhibit MMP expression or MMP enzyme activity (9-11) and uPA (12-14) may prevent cancer metastasis.
Quercetin (3,3’,4’,5,7-pentahydroxyflavone), a bioactive plant flavonoid, has been shown to induce cytotoxic effects such as anti-proliferation, induction of apoptosis and cell-cycle arrest in many human cancer cell lines (15-19). Numerous lines of evidence have also shown that quercetin induces cytotoxic effects on human cancer cells via cellular signal transduction pathways including change of the ratio of pro-apoptotic Bcl-2-associated X protein (BAX) and anti-apoptotic B-cell lymphoma-2 (BCL2) proteins, and mitogen-activated protein kinases (MAPKs) and protein kinase B (AKT) levels (19, 20). It was reported that quercetin is a potential inhibitor of phosphatidylinositide-3 kinases (PI3K), an enzyme involved in the pivotal cell survival pathway (21, 22). Although many studies have shown the cytotoxic effects and the possible mechanisms of quercetin action, whether and how quercetin inhibits the migration and invasion of oral cancer cells is still unclear. In the current study, we investigated the molecular signaling pathways of quercetin-induced inhibition of migration and invasion in human oral cancer SAS cells.
Materials and Methods
Materials and reagents. Quercetin, dimethyl sulfoxide (DMSO), propidium iodide (PI), ribonuclease-A, trypan blue, and Tris-HCl were obtained from Sigma-Aldrich Corp. (St. Louis, MO, USA). Dulbecco's modified Eagle's medium (DMEM), L-glutamine, fetal bovine serum (FBS), penicillin-streptomycin, and trypsin-EDTA were obtained from Life Technologies (Carlsbad, CA, USA). Primary antibodies for MMP-9 (cat. AB19016) and FAK (cat. 05-537) were purchased from Merck Millipore (Billerica, MA, USA), and antibodies against MMP-2, MMP-7, growth factor receptor-bound protein 2 (GRB2), cyclooxygenase-2 (COX-2), extracellular-signal-regulated kinase 1/2 (ERK1/2), Ras homolog gene family, member A (RhoA), phosphatidylinositide 3-kinases (PI3K), phosphorylated extracellular signal-regulated kinase (p-ERK), protein kinase C (PKC), rat sarcoma viral oncogene homolog (RAS), son of sevenless homolog-1 (SOS1), inductible nitric oxide synthase (iNOS) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and secondary antibodies, were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA) and diluted in PBS Tween-20 before use.
SAS cell culture and treatment. The SAS cell line (human oral squamous cell carcinoma) was kindly provided by Dr. Pei-Jung Lu (Graduate Institute of Clinical Medicine, National Cheng Kung University, Tainan, Taiwan). Cells were immediately plated onto 75 cm2 tissue culture flasks with DMEM with 2 mM L-glutamine adjusted to contain 10% FBS, 100 Units/ml penicillin and 100 μg/ml streptomycin. Then cells were maintained in 5% CO2 at 37°C until reaching approximately 50-70% confluence for further treatment. Cells at a density of 2×105 cells/well were maintained in a 12-well plate for 24 h then were treated with 25, 50, 100, 150, 200 and 400 μM of quercetin for 24 h. Cells were then harvested and were stained with PI (4 μg/ml) then analyzed by flow cytometry (FACS calibur flow cytometer; BD Biosciences, San Jose, CA, USA) as previously described (23, 24).

Quercetin affected the percentage of viable SAS cells in vitro. Cells were placed in RPMI-1640 medium + 10% Fetal bovine serum (FBS) with 25, 50, 100, 150, 200 and 400 μM of quercetin for 24 h. The cells were collected and were analyzed for viability by flow cytometry, as described in Materials and Methods. Each point is the mean±S.D. of three experiments.
Wound-closure migration assay. Cells were allowed to form a confluent monolayer in 6-well plates at 5×105 cells/well and then cells were wounded with a 200-μl pipette tip, as described previously (12, 25). After being washed twice with PBS, all cells in the dish were treated with or without quercetin at a final concentration of 0, 25 or 50 μM and then were incubated in fresh DMEM with 1% FBS for 24 h. Photographs were taken using a phase-contrast microscope. The wound closure was monitored for 24 h. The cell-free area of each treatment in the dish was observed and measured by an inverted microscope (OlympusIX71, Tokyo, Japan) as described previously (23). Cell migration was calculated as the percentage of the remaining cell-free area compared with the area of the initial wound. Each test was performed triplicates.
In vitro migration assay of SAS cells. Cell migration was assessed using chemotactic directional migration by using a 24-well Transwell insert (25, 26). About 30 μg type-I collagen (Merck Millipore) was used to coat the 8 μm pore filters (Cat. PIEP12R48; Merck Millipore) for 1 h then cells at a density of 104 cells/0.4 ml in DMEM were placed in the upper chamber and treated with 0.5% DMSO (as a control) or with quercetin (25 and 50 μM) then allowed to migrate for 24/48 h. A cotton swab was used to remove non-migrated cells in the upper chamber and the filter from each treatment was individually stained with 2% crystal violet. Migrated cells adherent to the underside of the filter were stained and photographed and then were counted under a light microscope at ×200. Each treatment including a control was assayed twice and three independent experiments were conducted as previously (25, 26).

Quercetin inhibited the migration of SAS cells. Cells were exposed to different concentrations (0, 25 and 50 μM) of quercetin for 0, 12 and 24 h then the migration distance of cells was evaluated by a wound-healing assay at 0, 12, and 24 h time points, as described in Materials and Methods.
In vitro invasion assay of SAS cells. Cell invasion assay was performed by using Matrigel-coated transwell cell culture chambers (8 μm pore size) (12, 27). Cells were cultured for 24 h in serum-free-DMEM then were collected and re-suspended in serum-free medium. The membrane at the bottom of the Transwell chamber was coated with 50 μl Matrigel (BD Biosciences, San Jose, CA, USA) and air-dried in a laminar hood overnight. Cells at a density of 5×104 cells/well were placed in the upper chamber of the transwell insert and treated with 0.5% DMSO (as a control) or quercetin (25 and 50 μM). Medium containing 10% FBS was placed in the lower chamber and cells were then incubated at 37°C in 5% CO2 atmosphere for 24 or 48 h. Non-penetrated cells were removed (which were maintained) in the upper chamber by using a cotton swab and the invasive cells in the lower surface of the filter which penetrated through the Matrigel were fixed with 4% formaldehyde in PBS and stained with 2% crystal violet in 2% ethanol. Cells were then counted and photographed under a light microscope at ×200 as described previously (12, 27).
Gelatin zymography for determination of MMP-2/-9 activity. The activity of MMP-2 and -9 in SAS cells was determined by gelatin zymography. In brief, cells (2×106 cells/well) were plated in 6-well tissue culture plates and were incubated in serum-free DMEM in the presence of 10, 15, 20, 25 and 50 μM quercetin for 24 h. The conditioned medium from each sample was collected by centrifugation then was separated by electrophoresis on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) containing 0.1% gelatin. After electrophoresis, the gel were soaked in 2.5% Triton X-100 in dH2O twice for a total of 60 min at 25°C, then were incubated for 18 h in substrate buffer (50 mM Tris HCl, 5 mM CaCl2, 0.02% NaN3 and 1% Triton X-100, pH 8.0) at 37°C (19). Bands corresponding to activity of MMP-2 and -9 were visualized by negative staining using 0.3% Coomassie blue in 50% methanol and 10% acetic acid as described previously (12, 26).
Western blotting analysis. SAS cells (1×106 cells/well) were placed in 6-well plates and then incubated with quercetin (25 and 50 μM) for 24 and 48 h at 80% culture confluence were harvested and re-suspended in PRO-PREP TM protein extraction solution (iNtRON Biotechnology, Seongnam-si, Gyeonggi-do, Korea). The homogenate from each sample was centrifuged at 13,000 ×g for 10 min at 4°C to remove cell debris and collect the supernatant. Bio-Rad protein assay kit (Hercules, CA, USA) was used to measure the total protein from each sample of supernatant. Proteins from each sample (40 μg) were separated by 10% SDS-PAGE and were transferred to an Immobilon-P transfer membrane (cat. IPVH00010; Merck Millipore) as described previously (25, 28, 29). Primary antibodies were used and then the membrane was washed for secondary antibody staining. The bound antibodies from each sample were visualized using Immobilon Western Chemiluminescent HRP Substrate (cat. WBKLS0500; Merck Millipore) and X-ray film (GE Healthcare, Piscataway, NJ, USA).
Confocal laser scanning microscopy showed protein translocation in SAS cells. Cells (5×104 cells/well) were placed on 4-well chamber slides, and quercetin at a final concentration of 25 μM was added to cells for 24 h. At the end of incubation, cells on the slides were fixed in 4% formaldehyde in PBS for 15 min and then were permeabilized with 0.3% Triton-X 100 in PBS for 1 h with blocking of non-specific binding sites using 2% Bovine serum albumin (BSA) (30). The fixed cells were stained by primary antibodies to RhoA, ROCK1 and NF-κB (1:100 dilution) (green fluorescence) overnight then were washed twice with PBS. All samples were the stained with secondary antibody [Fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG at 1:100 dilution], followed by DNA staining with mitotracker (Invitrogen, Carlsbad, CA, USA) (red fluorescence) as described previously (29, 30). All samples on 4-well chambers were examined and photomicrographed by using a Leica TCS SP2 Confocal Spectral Microscope (Leica Microsystems, HD, DE).

Quercetin-affected cell migration and invasion of SAS cells. Cells were treated with 0, 25 and 50 μM of quercetin for 24 and 48 h. Cell migration was examined in a Boyden chamber and transwell with polycarbonate filters (A); Cell invasion was examined in a Boyden chamber (C); polycarbonate filters (pore size, 8 μm) were pre-coated with matrigel. The migration (B) and invasion (D) ability of SAS cells were quantified by counting the number of cells that invaded the underside of the porous polycarbonate membrane under microscopy and represent the average of three experiments. ***p<0.001 as compared with the untreated control.
Statistical analysis. Data are presented as means±S.D. Differences of the variables between quercetin–treated and untreated (control) groups were analyzed by Student's t-test. Differences were considered significant when p<0.05.
Results
Effects of quercetin on the percentage of viable SAS cells in vitro. After SAS cells were treated with different concentrations of quercetin for 24 h, cells from each treatment were collected individually for PI staining in order to measure the percentage of viable cell. As shown in Figure 1, there were fewer viable cells (p<0.05), as concentration of quercetin increased at 24 h (Figure 1) compared to the control group. Quercetin induced concentration-dependent responses at both examined time points.
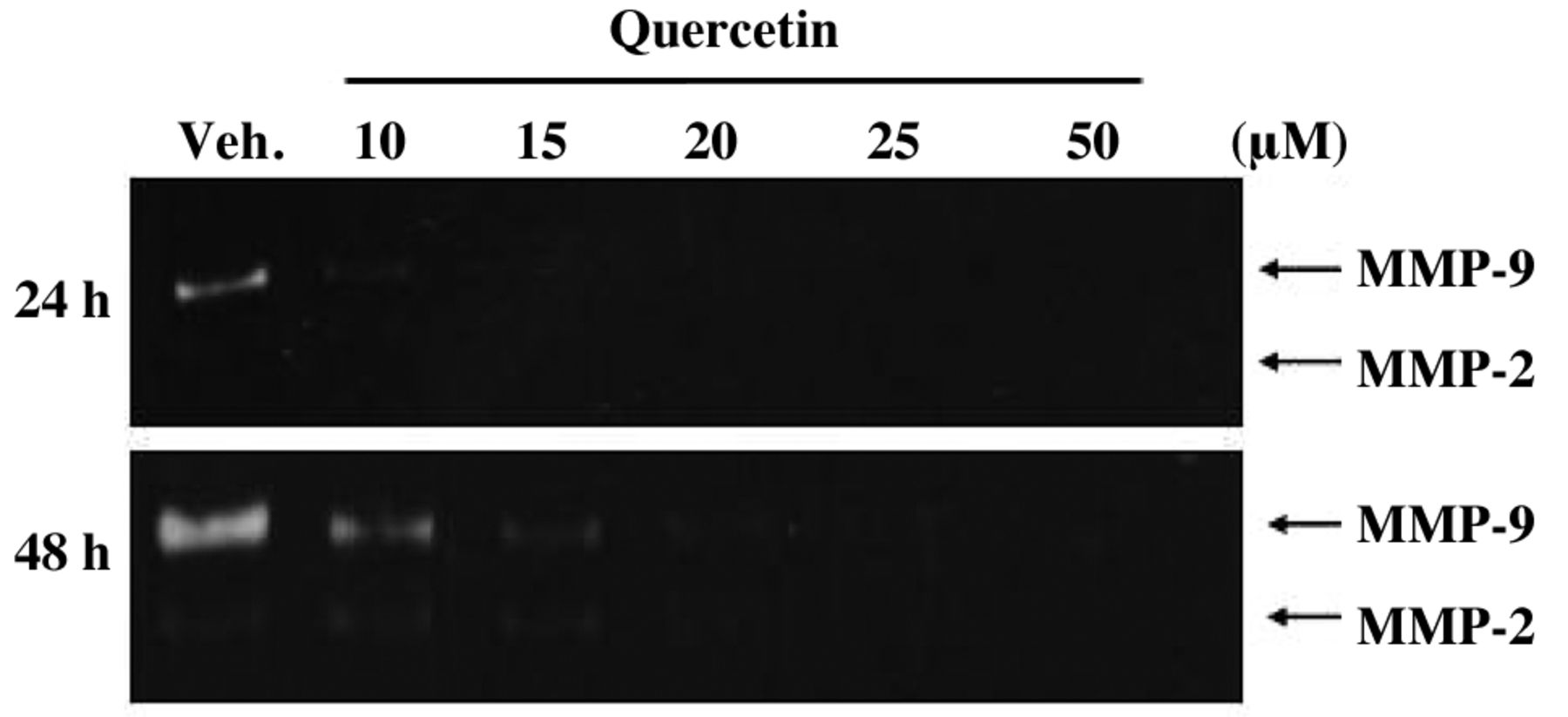
Quercetin inhibited as matrix metallopeptidase-2 (MMP-2) and matrix metallopeptidase-9 (MMP-9) activities in SAS cells. Cells were incubated with 0, 10, 15, 20, 25 and 50 μM of quercetin for 24 and 48 h. Cells were harvested and proteins separated by gelatin zymography, as described in Materials and Methods. The ratio of MMP-2 and MMP-9 activities was quantified. ***p<0.001 as compared with the untreated control.

Quercetin affected on the levels of proteins associated with migration and invasion in SAS cells. Cells were treated with 0, 25 and 50 μM of quercetin for 24 and 48 h and then harvested. The total proteins from each treatment were collected and the proteins levels (A: matrix metalloproteinase (MMP)-2, -7, -9 and -10 and vascular endothelial growth factor (VEGF); B: nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) p65, inductible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2) and urokinase-type plasminogen activator (uPA); C: phosphatidylinositide 3-kinases (PI3K), nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha (IKBα), IKB-α/β, phosphorylated nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor kinase, alpha/beta (p-IKKα/β); D: focal adhesion kinase (FAK), son of sevenless homolog-1 (SOS1), Ras homolog gene family, member A (RhoA), growth factor receptor-bound protein-2 (GRB2), protein kinase C (PKC), rat sarcoma viral oncogene homolog (RAS), mitogen-activated protein kinase kinase kinase-3 (MEKK3) and MEKK7; E: extracellular-signal-regulated kinase 1/2 (ERK1/2), phosphorylated-extracellular-signal-regulated kinase 1/2 (p-ERK1/2), c-Jun N-terminal kinase 1/2 (JNK1/2), p38, p-p38, Jun proto-oncogene (c-JUN) and phosphorylated-Jun proto-oncogene (p-c-JUN), were investigated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting, as described in Materials and Methods.

Quercetin affected Ras homolog gene family, member A (RhoA), Rho-associated, coiled-coil containing protein kinase-1 (ROCK1) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) p65 expression in SAS cells. Cells were incubated with quercetin at 25 μM for 24 h. Cells were the stained with RhoA (A), ROCK1 (B) and NF-κB p65 (C) FITC-labeled secondary antibodies (green fluorescence) and the proteins were detected by a confocal laser microscopy system. Scale bar 20 μm.

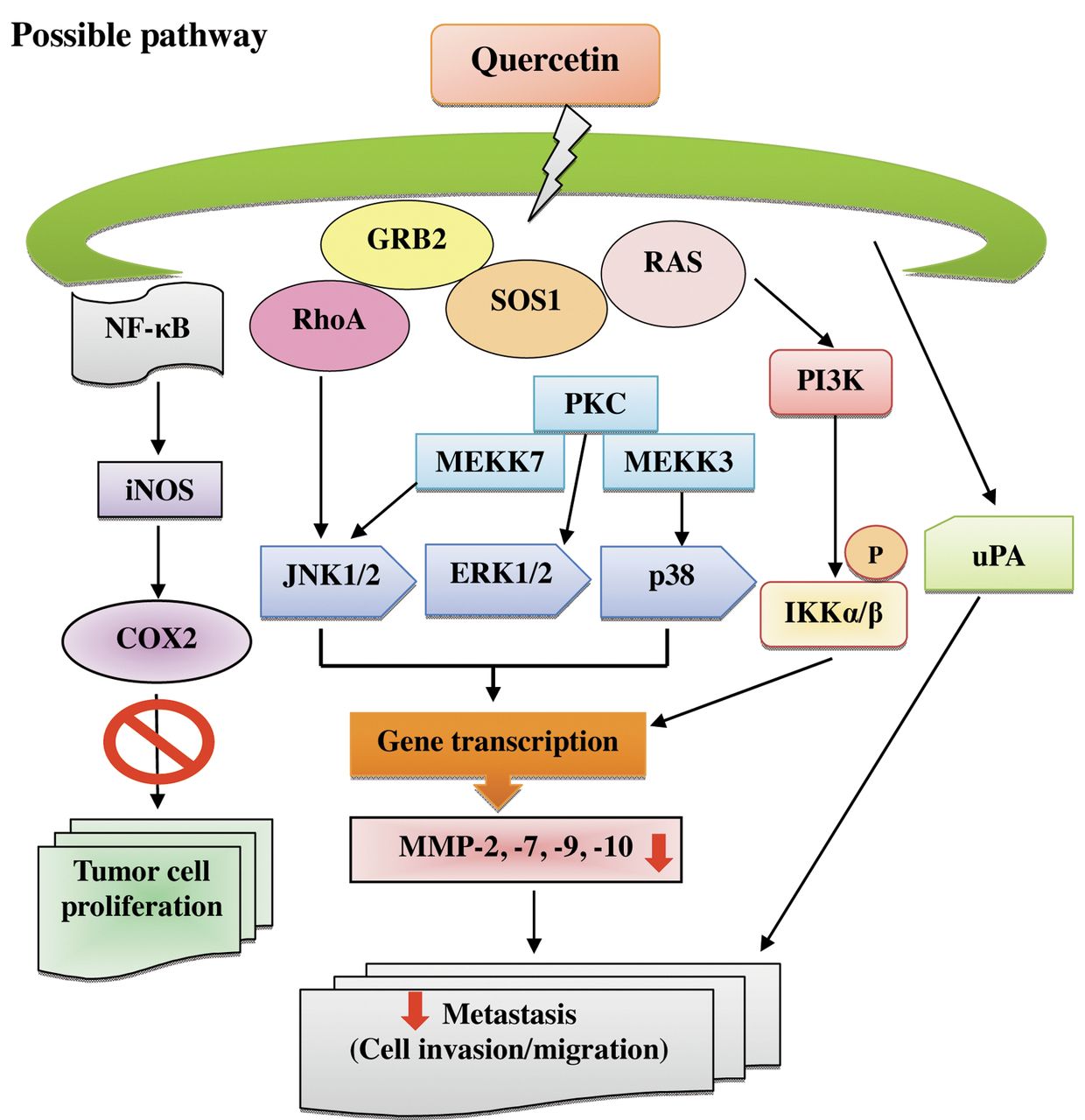
The proposed schematic presentation of the mechanism of quercetin-inhibited migration and invasion of SAS human oral cancer cells. (matrix metalloproteinase (MMP)-2, -7, -9 and -10, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) p65, inductible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), urokinase-type plasminogen activator (uPA), phosphatidylinositide 3-kinases (PI3K), phosphorylated nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor kinase, alpha/beta (p-IKKα/β), son of sevenless homolog-1 (SOS1), growth factor receptor-bound protein-2 (GRB2), mitogen-activated protein kinase kinase kinase-3 (MEKK3), MEKK7, extracellular-signal-regulated kinase 1/2 (ERK1/2), c-Jun N-terminal kinase 1/2 (JNK1/2), p38, Ras homolog gene family, member A (RhoA), protein kinase C (PKC) and rat sarcoma viral oncogene homolog (RAS).
Effects of quercetin on in vitro wound closure by SAS cells. We investigated the effects of quercetin on the migration of SAS cells by means of a wound closure assay. Figure 2 indicates that the relative wound closure was higher in control cells when compared with quercetin-treated cells. The inhibition was observed when cells were incubated with quercetin at 25 and 50 μM for a 12 and 24 h treatment, respectively. These effects demonstrated that quercetin inhibited cell migration in a dose- and time-dependent manner (Figure 2).
Effects of quercetin on migration and invasion of SAS cells. It is well-known that multiple steps and associated factors are involved in cancer cell migration and invasion (6, 7). Thus, we further investigated whether quercetin inhibits migration and invasion of SAS cells in vitro. The results from the migration assay showed that quercetin had a significant inhibitory effect on cell migration at concentrations between 25-50 μM (Figure 3A). Data in Figure 3B indicate that the inhibition was at least 42% and 44% at 24 and 48 h, when cells were treated with quercetin for 24 and 48 h, respectively. The invasion assay indicates that quercetin had a significant inhibitory effect on invasion (the penetration of the EHS-coated filter by SAS cells) of SAS cells, cell in the upper chamber to the lower chamber in the absence of quercetin (control group) were high (Figure 3C). The percentage inhibition at 25 and 50 μM was at least 62% and 55%, respectively (Figure 3D), when cells were incubated with quercetin for 24 and 48 h, respectively.
Effects of quercetin on MMP-2 and MMP-9 activities in SAS cells. Evidence has shown that MMP-2 and -9 play and important roles in invasion of human cancer cells (9-11). We examined whether quercetin could inhibit the secretion of MMP-2 and -9 in SAS cells by using gelatin zymography. After exposure to quercetin (0, 10, 15, 20, 25 and 50 μM) for 24 and 48 h, cells were harvested and the results of zymography are shown in Figure 4, which indicates that quercetin inhibited MMP-9 and MMP-2 activities of SAS cells.
Effects of quercetin on levels of protein expression associated with migration and invasion in SAS cells. For investigating whether or not quercetin inhibits the migration and invasion of SAS cells via the inhibition of associated proteins, the levels of proteins from each treatment were investigated by western blotting and results are presented in Figure 5. The results in Figure 5 show that quercetin reduced the protein levels of MMP-2, -7, -9 and -10 and VEGF (Figure 5A), NF-κB p65, iNOS, COX-2 and uPA (Figure 5B), PI3K, IKB-α, IKB-α/β, p-IKKα/β (Figure 3C), FAK, SOS1, GRB2, MEKK3 and MEKK7 (Figure D), ERK1/2, p-ERK1/2, JNK1/2, p38, p-p38, c-JUN and p-c-JUN (Figure 5E), but did not affected RhoA, PKC and RAS (Figure 5D) in SAS cells. It was reported that the ERK signaling pathway up-regulates the expression of MMPs (31, 32). We, thus, tested the effects of quercetin on the ERK signaling pathway. The results indicate that quercetin inhibited the phosphorylation of ERK1/2 (Figure 5E) in SAS cancer cells at 48 h, indicating that quercetin can inhibit the ERK signaling pathway.
Quercetin affected RhoA, ROCK1 and NF-κB expression in SAS cells. Based on the results from western blotting, revealing that quercetin affected the protein levels of RhoA, and NF-κB (Figure 5) in SAS cells, confocal laser microscopy examination indicated that quercetin promoted the expression of RhoA, ROCK1 and NF-κB (Figure 6) in SAS cells.
Discussion
It is well-documented that tumor metastasis occurs through multiple steps such as vessel formation and angiogenesis, cell attachment and adhesion, invasion, migration and cell proliferation, and that it is regulated through complex mechanisms (33). The migration and invasion of cancer cells involve the expression of MMPs (8, 9). The high incidence of lymph node spread and distant metastases associated with oral cancer make for poor prognosis of this disease (34). We previously elucidated the effect of quercetin on oral cancer cell death through cell-cycle arrest and induction of apoptosis. In this study, we investigated the effects of quercetin on the migration and invasion of SAS oral cancer cells (Figure 2 and 3) in vitro by using a Boyden chamber assay to quantify for the migratory potential of SAS cells.
The inhibitory effect of quercetin on SAS human oral cancer cells was concentration-dependent. Furthermore, we found that quercetin inhibited the migration and invasion of SAS cells through inhibition of expression and activity of MMP-2 and -9. The inhibition of MMP expression or enzyme activity can be used as early targets for preventing cancer metastasis (9, 11, 35). MMP-2 and -9 have been shown to be involved in the invasive metastatic potential of tumor cells (36, 37).
In the present study, we found that quercetin inhibited the invasion and migration of SAS cells, which may account for its inhibitory effect on tumor metastasis. Quercetin inhibited the protein expression and activities of MMP-2 and -9 (Figure 4 and 5A), inhibited the protein expression of ERK1/2 (Figure 5E), which is involved in promoting tumor invasion and metastasis, and inhibited the protein expression of JNK (Figure 5E). Therefore, we found that quercetin alone suppressed the proliferation and migration and invasion of SAS cells in vitro.
In this study, our results showed that quercetin reduced the protein levels of PI3K, NF-κB, MMP-2 and MMP-9. AKT is an important downstream target of PI3K for regulating cell proliferation and is involved in cell-cycle regulation and apoptosis (38, 39), and cell invasion (37, 40). Therefore, the P13K/AKT and MAPK pathways could play significant roles as potential targets for oral cancer treatment. Herein, based on those observations, we suggest that the regulation of NF-κB, downstream of PI3K and MAPK (ERK1/2, p38 and JNK) pathways, might be involved in quercetin suppression of MMP-2 and -9 expression and activity for the inhibition of SAS cell invasion and migration. Other investigators have shown that quercetin significantly reduces the rat sarcoma viral oncogene homolog (RAS) and v-raf-1 murine leukemia viral oncogene homolog-1 (RAF1) expression, thereby reducing the cell-cycle progression in PC-3 human prostate cancer cells (41). RAS regulates several cell-cycle proteins; it inactivates the Rb protein through the activation of G1/Cyclin-dependent kinases (CDKs) (42). However, the present study did not show that quercetin inhibited RAS expression in SAS cells. It was reported that quercetin inhibited the migration and invasion of PC-3 through downregulating uPA, uPAR, epidermal growth factor (EGF) and EGF-R mRNA expressions (41). uPA is a serine protease that is involved in cancer progression, especially invasion and metastasis (41). We found that quercetin inhibited the migration and invasion of SAS cells through the down-regulation of NF-κB, causing the inhibition of the protein expression and activity of MMP-2 and -9. Apparently, quercetin-inhibited migration and invasion of cancer cells may have cell specificity. Further investigations are needed to determine this.
In conclusion, this study has shown the quercetin, a plant compound, caused the inhibition of invasion and migration of SAS human oral cancer cells. Quercetin inhibited the activity of MMP-2 and -9 in a concentration-dependent manner. The possible signal pathways for quercetin inhibition of migration and invasion in SAS cells may be via inhibition of MMP-2 and MMP-9 via down-regulation of PKC, blocking of MAPK and PI3K signaling pathways and both COX-2 and NF-κB (Figure 7).
Acknowledgements
This study was supported by a research grant (DOH101-TD-C-111-005) from Taiwan Department of Health, China Medical University Hospital Cancer Research Center of Excellence.
Footnotes
-
↵* These Authors contributed equally to this study.
- Received February 20, 2013.
- Revision received April 1, 2013.
- Accepted April 2, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Herb-paths, a network and statistical model to explore health-beneficial effects of herbs and herbal constituents
- A Tricin Derivative from Deschampsia antarctica Desv. Inhibits Colorectal Carcinoma Growth and Liver Metastasis through the Induction of a Specific Immune Response
- 3,4,7-O-trimethylquercetin Inhibits Invasion and Migration of Ovarian Cancer Cells
- Cantharidin Impairs Cell Migration and Invasion of Human Lung Cancer NCI-H460 Cells via UPA and MAPK Signaling Pathways
- Citric Acid Induces Cell-cycle Arrest and Apoptosis of Human Immortalized Keratinocyte Cell Line (HaCaT) via Caspase- and Mitochondrial-dependent Signaling Pathways