


Abstract
Cancer metastasis is the major cause of cancer patient death. Melanoma is a highly important metastasis in human cancer. Cantharidin (CTD), identified as an active component of natural mylabris (Mylabris phalerata Pallas), induces apoptosis in many human cancer cells. In the present study, we investigated the anti-metastasis effects of CTD in human melanoma cancer A375.S2 cells. Flow cytometry was used to measure CTD-induced cytotoxic effects in A375.S2 cells. Wound healing assay indicated that CTD suppressed the migration of A375.S2 cells in a dose-dependent manner. The Matrigel Transwell Assay was used for cell migration and invasion examination and the results showed that CTD inhibited both. Gelatin zymography was used to investigate the activities of MMP-2/9 and the results indicated that CTD inhibited the enzymatic activities of MMP-2/9 in A375.S2 cells. The protein expression of A375.S2 cells following incubation with CTD was examined by western blotting and the results showed that CTD decreased the expression of ERK1/2, PI3K, FAK, MMP-2, -9, COX-2, NF-κB p65, TIMP 1, TIMP 2, VEFG, uPA, Rho A, GRB2, ROCK-1 and Ras, but increased the expressions of p38, JNK, p-c-jun and PKC. Based on those observations, we suggest that CTD may be used as a novel anti-cancer metastasis agent of human melanoma cancer in the future.
The degradation of the extracellular matrix (ECM), cell metastasis and proliferation are involved in neoplastic tumors invading normal tissues (1-3). Neoplastic metastasis has been recognized to be a major cause of cancer-mediated death, playing an important role in tumor progression and is the major cause of mortality in patients with cancer (4, 5). Tumor metastasis involves cell attachment (adhension), migration and invasion forming new tumor(s) in new sites. Cell adhesion molecules (integrin, cadherin and immunoglobulin superfamilies) have been studied in the context of tumor metastasis (6). Human melanoma, an aggressive skin cancer, has a low survival ratio (7, 8).
In physiological and pathological processes, matrix metalloproteinases (MMPs), a family of zinc-dependent proteinases, are involved in the degradation and remodeling of ECM that is associated with cell migration and invasion. Overexpression of MMPs is correlated with invasive and metastatic properties of cancer (9, 10) and, among them, MMP-2/9 can degrade matrix collagen and basement membrane (11, 12). It has been reported that MMP-9 can regulate the bioavailability of vascular endothelial growth factor (VEGF) through its receptor VEGFR2 (13). Furthermore, inhibition or activation of MMP-2/9 expression could be a preventive means towards cancer metastasis (14), rendering MMP-2/9 a good target for anticancer drug development.
Cantharidin (CTD) belongs to the terpenoids and is isolated from blister beetles and other insects. Numerous studies have shown that CTD induces apoptosis in many human cancer cells (15-20) and is an inhibitor of protein phosphatases 1 and 2a (21). Recently, in our laboratory, we found that CTD induced DNA damage and inhibited DNA repair gene expression (22). It also induced cell apoptosis (23) in human lung cancer H460 cells in vitro. Furthermore, we found that CTD inhibited cell migration and invasion of human bladder cancer cells through the inhibition of MMP2/9 in vitro (24). However, there is no available information to show that CTD inhibits migration and invasion of human melanoma cells. Therefore, the purpose of this study was to investigate the molecular mechanisms of CTD by which it suppresses the migration and invasion of human melanoma A375.S2 cells in vitro.
Materials and Methods
Chemicals and reagents. Cantharidin (CTD), dimethyl sulfoxide (DMSO) and propidium iodide (PI) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Minimum essential medium (MEM), fetal bovine serum (FBS), L-glutamine and penicillin-streptomycin were purchased from GIBCO®/Invitrogen Life Technologies (Carlsbad, CA, USA). Primary antibodies against ERK1/2, p38, JNK, p-c-jun, PI3K, GRB2, PKC, FAK, MMP-2, -9, COX-2, NF-κB p65, TIMP 1, TIMP 2, VEFG, uPA, Rho A, ROCK-1, RAS and anti-β-actin (a mouse monoclonal antibody) were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA). The enhanced chemiluminescence (ECL) detection system was obtained from Amersham Life Sciences, Inc. (Arlington Heights, IL, USA).
Cell culture. The A375.S2 human melanoma cancer cell line was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan). Cells were kept at 37°C, 5% CO2, in MEM medium supplemented with 10% heat-inactivated fetal calf serum, 2 mM glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin (25).
Cellural viability assays. A375.S2 cells were placed in a 12-well plate at a density of 1×105 cells/well and maintained for 24 h. Subsequently, the cells were treated with CTD at various concentrations (0, 1, 2, 3, 4 and 5 μM) for 24 h. At the end of the incubation period, cells were trypsinized and total cell number was measured by using flow cytometry (BD Biosciences, FACS Calibur, San Jose, CA, USA) as previously described (25, 26).
Wound healing assay. A375.S2 cells (1×105 cells/well) were maintained in petri dishes for 24 h. Once monolayers were established, a sterile yellow micropipette tip was used to create wounds in each plate. The unscraped cells in the plate were washed with PBS three times, dead cells were removed and fresh MEM medium supplemented with FBS containing 0 and 4 μM of CTD was added for 0, 12, 24 and 48 h. At the end of the incubation time, the wound healing area in each well was photographed with an inverted microscope as described previously (25, 26). The rate of wound closure was observed at the indicated time.
Cell migration and invasion assays. The cell migration assay and matrigel invasion system were used to measure cell migration and invasion of A375.S2 cells in vitro, as described previously (25, 26). For cell migration, transwell cell culture chambers (8 mm pore size; Millipore, Billerica, MA, USA) without coated matrigel were used. A375.S2 cells (1×105 cells/ml) with serum-free MEM medium were placed in the upper chamber of the transwell insert and various concentrations of CTD (0, 2 and 4 μM) were added on the top of the chamber. The 90% MEM medium containing 10% FBS was added to the lower chamber as chemoattractant. At the end of incubations, non-migrating cells were removed from the top chamber with a cotton swab. The cells on the lower surface of the chambers were fixed with 4% formaldehyde in PBS. Then, the chambers were rinsed with PBS and cells were stained with 2% crystal violet in 2% ethanol for 10 min. Subsequently, 10 fields were selected and photographed under a light microscope at 200×. For the cell invasion assay, the filter membrane was coated with Matrigel from a BioCoat Matrigel invasion kit and all experiments were performed as for the cell migration assay. All cells that invaded the underside of the filter were examined at 10 images, counted and photographed under a light microscope at ×200 as described previously (25, 26). Data are expressed as the percentage of invasive relative to migrating cells.
Gelatin zymography assay. The enzyme activities of MMP-2 and MMP-9 were analyzed by the gelatin zymography assay, as described previously (25, 26). A375.S2 cells (1×105 cells/well) were plated in 12-well tissue culture plates and allowed to adhere for 12 h in the presence of serum. Subsequently, the culture medium was replaced by serum-free MEM medium (1 ml per well) with different concentrations of CTD (0, 1, 2, 3 and 4 μM). After incubation for 24 h, the conditioned medium from each treatment was gathered and centrifuged for 10 min at 1,500 rpm to remove cell debris. The supernatant was separated by electrophoresis on 10% SDS-PAGE containing 0.2% gelatin. Then, the gel was soaked in 2.5% Triton-X-100 for 30 min twice to remove SDS and transferred to substrate buffer (50 mM Tris HCl, 5 mM CaCl2, 0.02% NaN3 and 1% triton X-100, pH 8.0) for 24 h, at 37°C, while shaking. Finally, the gel was stained with 0.1% Coomassie blue in 45% methanol and 10% acetic acid. The presence of clear bands (negative staining) indicated the presence of a protein with MMP-2 and MMP-9 gelatinolytic activity.
Western blotting. A375.S2 cells (1×105 cells/well) in a 12-well plate were incubated with 0 and 4 μM of CTD for 6, 12, 24 and 48 h. Cells were collected and washed before whole-cell extracts were lysed in lysis buffer [40 mM Tris-HCl (pH 7.4), 10 mM EDTA, 120 mM NaCl, 1 mM dithiothreitol, 0.1% Nonide P-40] and total protein was determined as described previously (25, 26). Whole cell lysates were clarified by centrifugation at 12,000 × g for 10 min at 4°C. The amounts of protein from each treatment were normalized by the Bradford method (25, 26). Equal amounts of total proteins (30 μg) were loaded onto 10% SDS-PAGE gels and subsequently transferred to PVDF membranes (Amersham Pharmacia Biotech, Piscataway, NJ, USA) by electro-blotting. Then the blots were probed overnight at 4°C by using primary antibodies against ERK1/2, p38, JNK, p-c-jun, PI3K, GRB2, PKC, FAK, MMP-2, -9, COX-2, NF-κB p65, TIMP 1, TIMP 2, VEFG, uPA, Rho A, ROCK-1 and Ras, while anti-β-actin (a mouse monoclonal antibody) was used as a loading control. The secondary antibody (anti-rabbit IgG) was added for 1 h for aiding detection by enhanced chemiluminescence (NEW Life Science Products, Inc, Boston, MA, USA), as described previously (25, 26).

CTD affects the percentage of viable A375.S2 human melanoma cancer cells. A375.S2 cells (1×105 cells/well) were incubated with 0, 1, 2, 3, 4 and 5 μM of CTD for 24 h and the percentage of viable cells was determined by flow cytometry, as described in Materials and Methods. *p<0.05, significant difference between CTD-treated groups and the control as analyzed by the Student's t-test.
Statistical analysis. All data were presented as the mean±standard deviation (S.D.) of at least 3 independent experiments. Statistically significant differences between CTD-treated and -untreated (control) groups were tested by the Student's t-test. A probability value less than 0.05 was considered statistically significant.
Results
CTD decreased the cell viability of A375.S2 cells. A375.S2 cells were treated with CTD (0, 1, 2, 3, 4 and 5 μM) for 24 h and the percentage of viable cells was evaluated. Figure 1 shows a significant reduction of living cells with CTD at 3-5 μM concentrations in A375.S2 cells. Thus, the 4 μM concentration of CTD was selected for further migration and invasion experiments.
CTD inhibited cell migration of A375.S2 as assessed by the wound healing assay. The capacity of cellular migration was evaluated using a scratch wound-healing assay. The results, shown in Figure 2 demonstrate that CTD-treated cells (4 μM) for 0, 12, 24 and 48 h close to the area of scratch in the plate decreased significantly in a time-dependent manner. These findings indicated that CTD inhibited cell migration of A375.S2 in vitro.
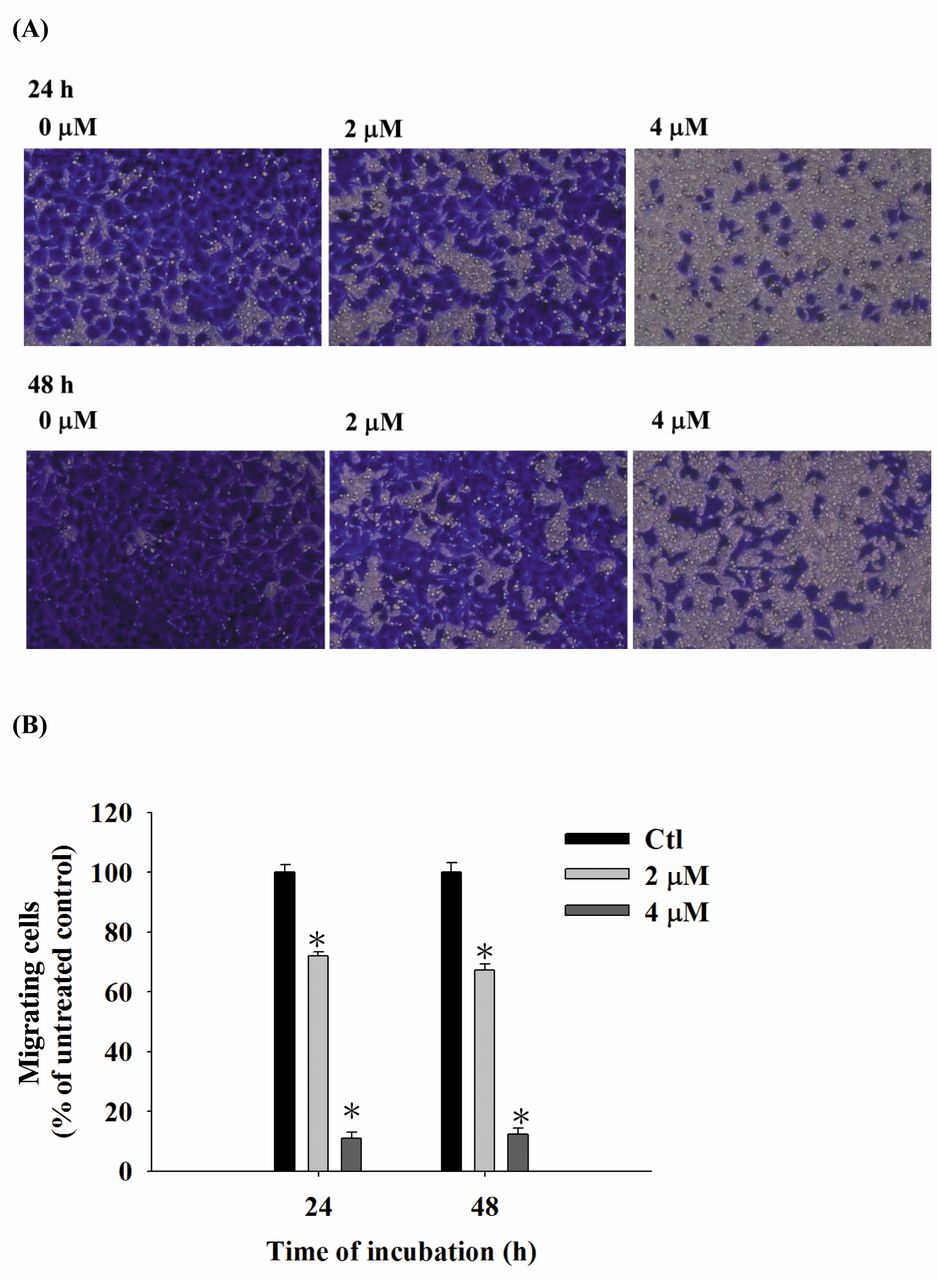
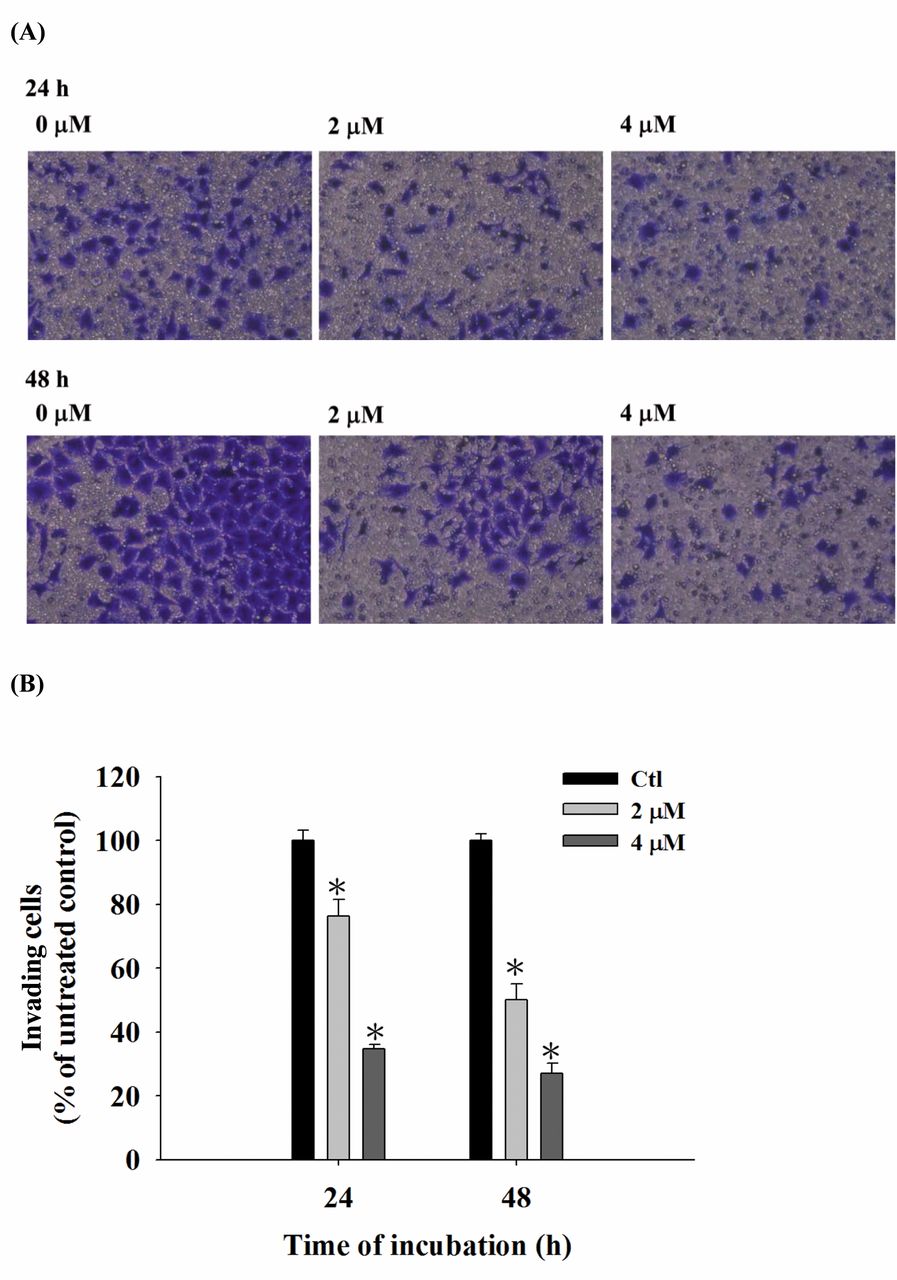
CTD inhibited both cell migration and invasion of A375.S2 cells. The cells were cultured for 24 and 48 h in the presence and absence of CTD (2 and 4 μM) and, subsequently examined, counted and photographed. The results are shown in Figure 3A and B. CTD significantly decreased cell migration by 20-80% and 26-80% at 2 and 4 μM, respectively, compared to control cells (Figure 3A and B). Results from Figure 4A and B also indicate that CTD significantly (p<0.05) inhibited the invasion ability by 35-72% and 51-28% at 2 and 4 μM, respectively, compared to control cells.

CTD inhibits wound closure in A375.S2 cells. A375.S2 cells (1×105 cells/well) were maintained in petri dishes, wounded and then treated with CTD (0 and 4 μM) for 0, 12, 24 and 48 h in 0.5% FBS-containing medium. Subsequently, phase-contrast pictures of the wounds at three different locations were taken as described in Materials and Methods.
CTD inhibited the activities of MMP-2 and -9 in A375.S2 cells. The potential effects of CTD treatment on MMP-2 and MMP-9 production by A375.S2 cells were studied by gelatin zymography to detect the gelatinolytic activity in conditioned media of A375.S2 cells treated by CTD. The results are shown in Figure 5. Treatment of A375.S2 cells with CTD had significant inhibitory effects on the levels of MMP-2 and MMP-9 activity (Figure 5). These findings suggest that inhibition of migration and invasion by CTD was caused by changes in gelatinase secretion or activation.

CTD suppressed the migration of A375.S2 cells in vitro. Cells (2×104 cells/well) were placed on a collagen-coated filter and incubated with 0, 2 and 4 μM of CTD for 24 and 48 h. Then, cells that penetrated through the lower surface of the filter, were collected, stained with crystal violet, photographed under a light microscope at ×200 (A) and counted (B). Results are from three independent experiments. *p<0.05, significant difference between CTD-treated groups and the control as analyzed by the Student's t-test.

CTD suppressed the invasion of A375.S2 cells in vitro. Cells (2×104 cells/well) were placed on slides coated with Matrigel and incubated with 0, 2 and 4 μM of CTD for 24 and 48 h. Cells that penetrated through the Matrigel to the lower surface of the filter were stained with crystal violet, photographed under a light microscope at x200 (A) and counted (B). Results are from three independent experiments. *p<0.05, significant difference between CTD-treated groups and the control as analyzed by the Student's t-test.
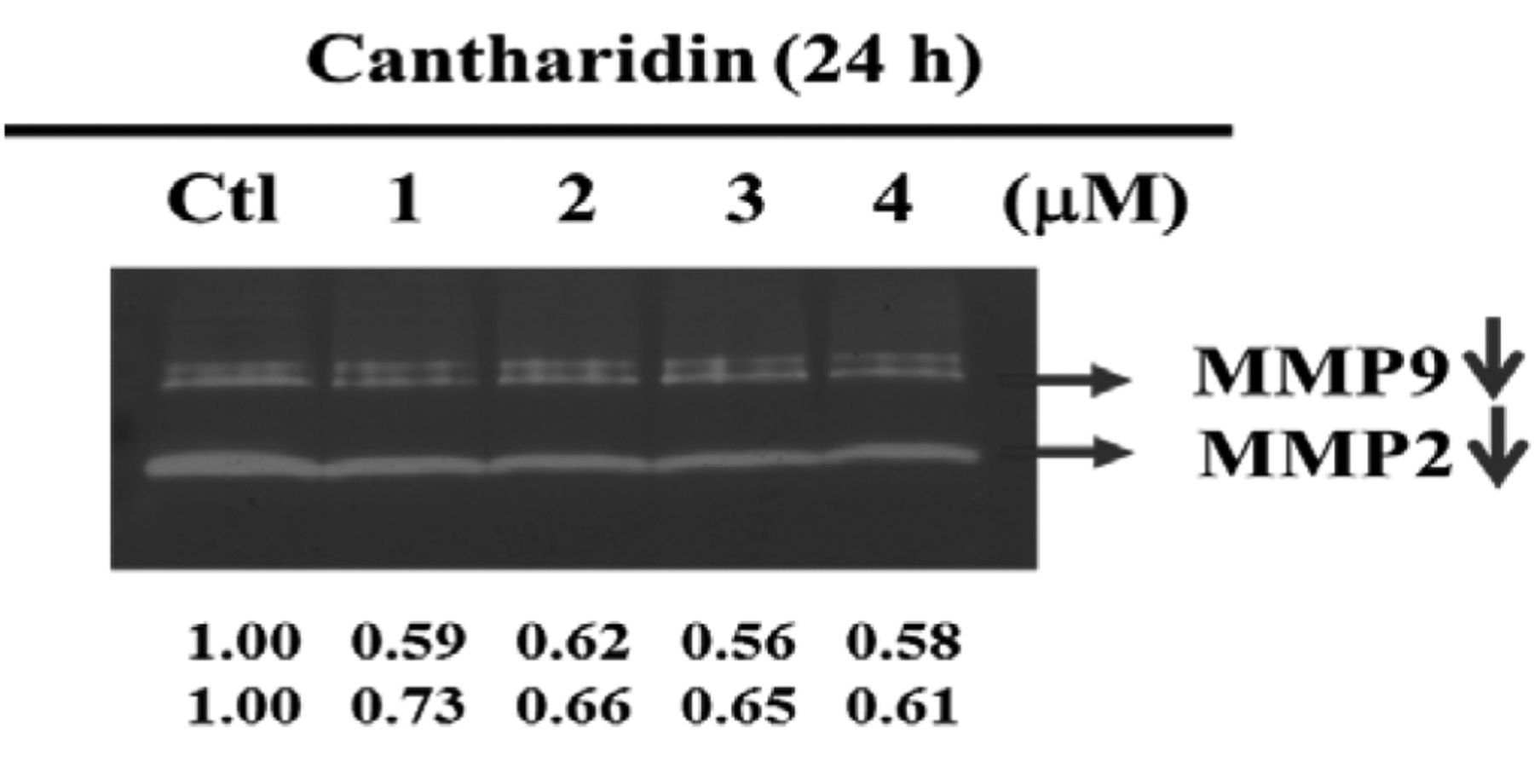
CTD affects the activities of MMP-2/9 in A375.S2 cells. Cells (1×105 cells/well) were incubated with 0, 1, 2, 3 and 4 μM of CTD for 24 h and their conditioned medium was collected for gelatin zymography as described in Materials and Methods. The different activity of MMP-2 and MMP-9 was determined by densitometry analysis and the results are expressed as percentage of the control (100%).
CTD alters levels of proteins associated with migration and invasion of A375.S2 cells. Western blotting was performed to examine the protein expression associated with migration and invasion in A375.S2 cells. It is well-documented that MMP-2 and -9 are potential targets for anti-metastatic drug function. To further explore the link among FAK and MMPs in A375.S2 cells after exposure to CTD, we examined MMP-2 and -9 and associated up-stream protein levels in A375.S2 cells. The results are shown in Figure 6 depicting that CTD significantly reduced the protein levels of ERK1/2 (A), PI3K, GRB2 and FAK (B), MMP-2, -9, COX-2 and NF-κB p65 (C), TIMP 1, TIMP 2, VEFG and uPA (D), Rho A, ROCK-1 and Ras (E) but increased the expressions of p38, JNK, p-c-jun (A) and PKC (B).
Discussion
Metastasis is the major cause of death for patients with cancer. Its diverse set of clinical features (27) that proceed through a complex multistep process involve cell adhesion, motility, proteolysis degradation of ECM, angiogenesis and invasion (28). Therefore, interruption of one or more of these steps in cancer development may indicate a potential approach for anti-metastatic therapy in a patient (29). A number of studies have demonstrated that CTD induces cell death, cell-cycle arrest and apoptosis in human cancer cells; however, no available information exists showing CTD's ability to inhibit migration and invasion of human melanoma cells. In this study, we investigated the effect of CTD on the migration and invasion of A375.S2 human melanoma cells and the results demonstrated that CTD can inhibit their metastatic process.
The scratch motility (wound healing) assay was used first to investigate the migration of A375.S2 cells and whether or not they were affected by CTD. The results (Figure 2) indicated that CTD significantly reduced the migration of A375.S2 cells. A transwell-assay was then used to examine cell migration and invasion of A375.S2 cells and whether or not they were affected by CTD. As observed in Figure 3A and B, CTD reduced the migration of A375.S2 cells, which comes in agreement with the results from the wound healing assay. Subsequently, the effect of CTD on A375.S2 melanoma cancer cell invasion was investigated by using a matrigel chamber invasion assay. It was found that CTD displayed an obvious inhibition of invasion capability in a concentration-dependent manner (Figure 4A and B).
It has been reported that the expression and activity of MMPs against matrix macromolecules is linked to the promotion of cell invasiveness and metastasis (30). Herein, CTD's anti-invasive action is also reflected by its suppressive effects on the activities of MMP-2 and MMP-9 (Figure 5), that are two major MMPs mediating the degradation of ECM. It is known that the breakdown of the basement membrane is a critical step for invasion and metastasis of cancer cells that proceeds via the activation of MMPs (31, 32). In this study, CTD treatment not only decreased the enzymatic activity, but also reduced the protein expression of MMP-2 and MMP-9. Based on these results, we may suggest that CTD's anti-invasive action is mediated partly via diminishing the ability of melanoma cancer cells to degrade the components of ECM by modulating MMP-2 and MMP-9 expression and activity in A375.S2 cells, supporting already published findings that inhibition of MMP expression or enzyme activity can be considered as early targets in the prevention of cancer metastasis cascade (33-35).
MMP-2/9 plays an important role in cancer invasion and metastasis and transcription of MMP-2/9 genes is regulated by upstream regulatory factors including NF-κB, c-Jun and AP-1 (36-38). Thus, we used western blotting to investigate the expression of upstream regulatory factors of MMP-2/9 in A375.S2 cells after exposure to CTD for various time periods. Firstly, we found that CTD significantly reduced the protein expression of NF-κB, p65 levels (Figure 6C) when compared to the control groups. Further, as depicted in Figure 6, CTD reduced the protein levels of PI3K, ERK1/2, Rock 1 and FAK in A375.S2 cells. It has been reported that the PI3K-Akt signaling pathway plays a significant role in MMPs concerning uPA gene regulation, cell survival and cell invasion (39, 40). Based on these observations, we found that CTD also affected the MAPK signaling pathway due to the reduction of protein expression, such as p38, JNK and ERK. Thus, CTD inhibited the migration and invasion of A375.S2 cells via suppression of the MAPK signaling pathway.
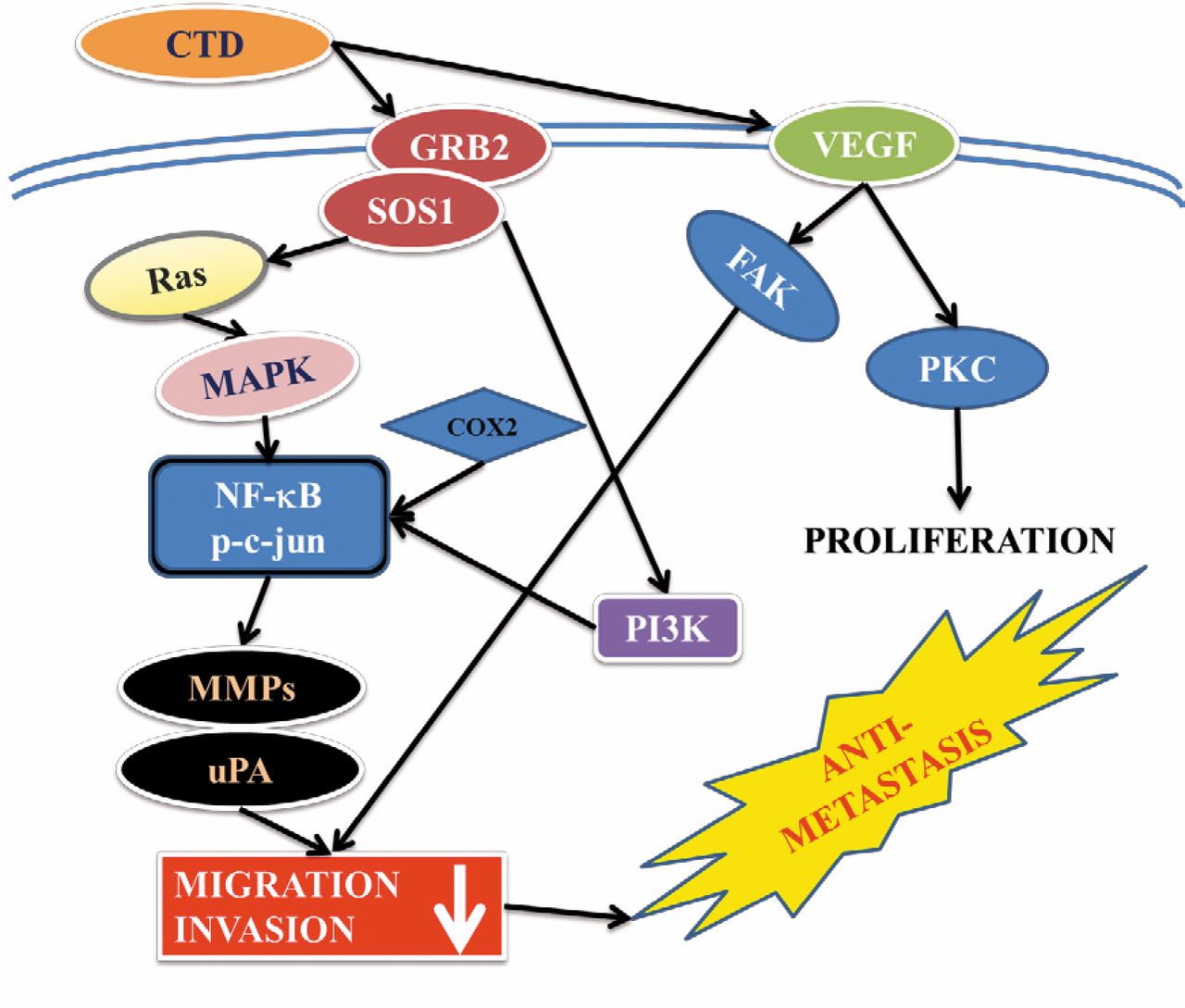
CTD inhibited the MAPK (p38, ERK and JNK) signaling pathway by reducing AKT and NF-κB leading to MMP-2/-9 down-regulation and uPA protein expression as summarized in Figure 7. These results show that CTD may be a potential candidate for developing preventive agents against human melanoma cancer metastasis. CTD should be further tested in vivo to justify its effectiveness in the prevention of invasion or migration in animal models of melanoma cancer.

CTD affects the levels of associated proteins in migration and invasion of A375.S2 cells. Cells (1×105 cells/well) were treated with CTD 4 μM for 0, 6, 12, 24 and 48 h and total protein was determined by SDS-PAGE gel electrophoresis, as described in the Materials and Methods section. The levels of ERK1/2, p38, JNK, p-c-jun (A), PI3K, GRB2, PKC and FAK (B), MMP-2, -9, COX-2 and NF-κB p65 (C), TIMP 1, TIMP 2, VEFG and uPA (D), Rho A, ROCK-1 and RAS (E) expression were estimated by western blotting as described in Materials and Methods.

The possible signaling pathways for the CTD-mediated inhibition of migration and invasion in A375.S2 human melanoma cancer cells.
Taken together, our results demonstrate that CTD mediated the inhibition of migration and invasion in melanoma cancer cells, through metastasis-associated proteases and their natural upstream-associated proteins. We attribute the mechanism underlying the above effects to the attenuation of the activity and expression of MMP-2 and MMP-9. Furthermore, CTD inhibited the MAPK (p38, ERK and JNK) signaling pathway via NF-κB leading to MMP-2/9 protein expression. This study suggests that CTD is a potential candidate for interventions against melanoma cancer metastases.
Acknowledgements
This study is supported in part by a research grant from China Medical University [CMU102-ASIA-20].
Footnotes
-
↵* These Authors contributed equally to this study.
- Received September 11, 2014.
- Revision received October 17, 2014.
- Accepted October 22, 2014.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Demethoxycurcumin Suppresses Proliferation, Migration, and Invasion of Human Brain Glioblastoma Multiforme GBM 8401 Cells via PI3K/Akt Pathway
- Tissue Rebuilding During Spontaneous Regression of Melanoma in the Melanoma-bearing Libechov Minipig
- Bisdemethoxycurcumin Suppresses Migration and Invasion of Human Cervical Cancer HeLa Cells via Inhibition of NF-ĸB, MMP-2 and -9 Pathways
- Demethoxycurcumin Suppresses Migration and Invasion of Human Cervical Cancer HeLa Cells via Inhibition of NF-{kappa}B Pathways
- Phenethyl Isothiocyanate (PEITC) and Benzyl Isothiocyanate (BITC) Inhibit Human Melanoma A375.S2 Cell Migration and Invasion by Affecting MAPK Signaling Pathway In Vitro