


Abstract
Background/Aim: Although insulin-like growth factor-1 (IGF-1) is well-implicated in cancer biology, the potential roles of IGF-1 variants in different types of cancer are largely unknown. The aim of the present study was to in vitro characterize several human cancers for their IGF-1 variant expression patterns. Materials and Methods: Using a novel quantitative real-time polymerase chain reaction (qRT-PCR) assay, twelve human cancer cell lines were investigated for their endogenous expression levels of IGF-1 variants, including classes. Additionally, the hormonal regulation of IGF-1 transcripts was investigated in PC3 cells. Results: IGF-1Ea and Eb were found at higher levels in KLE and MEL28 cells, respectively. MCF7 had the lowest expression of Ec peptide and, along with MB231, lacked IGF-1Eb. In most cases, class 1 proved as the predominant origin. Estradiol (E2) or dexamethasone (Dexa) significantly modulated IGF-1Ea and IGF-1Eb and down-regulated the Ec peptide in PC3. Conclusion: Our results contribute to the notion of distinct roles of IGF-1 isoforms in human cancer depending on the type of malignancy.
The insulin-like growth factor-1 (IGF-1) is a potent mitogen of major importance in human physiology and development with numerous proliferative and anti-apoptotic effects addressed (1). The 100kb IGF-1 gene is located on the long arm of chromosome 12 (12q22-12q24) and it consists of 6 exons (2, 3). Combination of different transcription initiation sites, alternative splicing at both 5’ and 3’ ends and post-translational modifications, such as proteolytic processing and glycosylation events, give rise to multiple products (isoforms) (4). More specifically, the precursor IGF-1 polypeptides consist of the signal peptide, mature IGF-1 and the E peptides; mature IGF-1 is a single-chain 70-amino-acid (aa)-peptide consisting of exons 3 and 4, while at the carboxyl (C)-terminal, extension of the pro-IGF-1 peptides and combination of exons 5 and 6 results in distinct E peptides, named Ea, Eb and Ec. Thus, alternative splicing, containing only exon 5, results in the IGF-1Eb isoform, whereas containing exon 6 but excluding exon 5 results in IGF-1Ea (5). A third variant, the IGF-1Ec, is also generated by alternative splicing in the human IGF-1 gene and contains both exons 5 and 6 (6). At the N-terminal, usage of different promoters, located either in exon 1 or 2 produces class 1 or class 2 IGF-1 molecules, respectively. Pro-IGF-1 peptides consist of mature and E peptides (4) (Figure 1A).
Although over the last decade an increasing number of studies indicate a differential association of IGF-1 isoforms with several pathophysiological processes, such as myocardium remodeling, skeletal muscle regeneration and endometriosis (7-9), the potentially distinct molecular functions of IGF-1 E peptides, and especially of its different classes, are largely unknown. Interestingly, over the last years, focus has turned into clarifying the role of specific IGF-1 isoforms in cancer biology (9-12). The differential expression pattern of IGF-1 isoforms observed in in vitro models, indicate a unique and distinct role of pro-peptides in different cancer types including osteosarcoma, prostate, endometrial and cervical malignancies (10–12). However, less is known about the possibly specific roles of the IGF-1 isoforms or, particularly, of its different classes in the various types of cancer.
Against this background, the aim of the present study was to characterize several immortalized cell lines, the majority of which served as models of numerous human cancer types, regarding their expression pattern of IGF-1 isoforms at the mRNA level, including their classes, under control and/or hormonal conditions. Twelve cell lines were used simulating, among others, breast, hepatocellular, lung, melanoma and colon malignancies. The results presented herein indicate for the first time the distinct identification of IGF-1 transcripts in so many malignant tissue culture models, further supporting the notion for differential and unique roles of IGF-1 isoforms in human cancer biology, depending on the type of malignancy.
Sequences of oligonucleotides used for amplification and quantification of the distinct IGF-1 variants in the qRT-PCR assay and the resulting products' length.
Materials and Methods
Primer design. All primer sets designed de novo, for increased efficiency and specificity, to be complementary to the unique exons corresponding to each IGF-1 transcript (Figure 1A) and within the coding region using the sequencing provided by the National Center for Biotechnology Information (NCBI), while their specificity was checked using the NCBI Primer BLAST tool. Forward primers for pro- and class 1 or 2 IGF-1 peptides were common for all primer sets and located to exon 4 and 1 or 2-3 (junction) exons, respectively, while reverse primers for pro-IGF1-Ea and -Eb were located to exons 4-6 (junction) and 5, respectively. Amplification of the IGF-1Ec transcript corresponding to the last (C-terminal) 24 aa of bioactive protein (Ec peptide) was achieved using a primer's pair complementary to exon 5 (sense) and exon 6 (antisense) (Table I, Figure 1A). Efficiency of all primer's sets was validated before tested to cells using serial dilutions in cDNA obtained from human muscle tissue, which served as positive control. All sets were found to have acceptable efficiencies, between 90 and 120% (Figure 2A), and unique curve picks in predicted for their length and GC content melting temperature, as shown in melting curve analysis (Figure 3A-I) in the same cycling conditions. Both sequencing (not shown) and electrophoretic analysis of the real-time PCR products in agarose gel (e.g., Ec peptide is shown, in Figure 2B) further verified the specificity of each target transcript. Although all primer sets were designed to correspond to intron-exon spanning regions or to exon-exon junctions (Figure 1A and B), they were checked for their contaminating DNA amplification potential in a boosted PCR reaction of 45 cycles using 300 ng of genomic DNA as template. The resulting PCR product was further analyzed in agarose gel electrophoresis where no bands were observed (Figure 2C). Primers designed for amplification of genomic sequence of cyclophilin were used as positive control of the reaction.
Cell cultures and treatments. The following employed normal and cancerous human cell lines were obtained from the American Type Culture Collection (ATCC, Bathesda, MD, USA): normal lens epithelial (HLE-B3), androgen sensitive prostate adenocarcinoma (LnCaP), androgen negative prostate cancer (PC3), estrogen positive breast adenocarcinoma (MCF7), estrogen negative breast adenocarcinoma (MDA-MB-231), hepatocellular carcinoma (HuH7), normal kidney (HEK293), endometrial cancer (KLE), epithelial cervical cancer (HeLa), lung cancer (A549), melanoma (SK-MEL28), osteosarcoma (MG63) and colon cancer cells (DLD1). All cell lines were cultured in selective growth medium supplemented with 10-20% fetal bovine serum (FBS) and/or 2mM L-glutamine (Lonza, Basel, Switzerland) according to ATCC instructions and 1% penicilin/streptomycin (GIBCO, Life Technologies, Grand Island, NY, USA) and maintained in an tissue culture incubator at 37o C and 5% CO2. At control conditions, cells reached confluence of approximately 80% before harvesting. Adherent cells were washed twice with phosphate buffer salina (PBS) prior to trypsinization (GIBCO). Estradiol (E2), Dexamethasone (Dexa) and Growth Hormone (GH) (Sigma-Aldrich, St. Louis, MO) were selected as treatment factors and used in concentrations of 1, 10, 25 and 100 nM for 24 and 48 h in a dose- and time-response manner respectively.
RNA extraction and cDNA synthesis. Total RNA was extracted from cells using the TRI reagent RT-111 (RT-111; MRC Inc., Cincinnati, OH, USA) according to manufacturer's instructions. RNA concentration was photometricaly measured using the optical density absorbance at 260 nm in a nano-drop spectophotometer, whereas the absorbance ratio at 260/280 nm and the identification of 18s and 28s bands in 1% agarose gels after RNA electrophoresis were used to evaluate the quality of extracted RNAs (not shown). Following DNase treatment (Invitrogen, Life Technologies, Grand Island, NY, USA) one microgram (1 μg) of each RNA were processed to cDNA synthesis using 0.5 μg oligo DT primers (Fermentas, GmbH, Germany), 0.5 mM dNTPs (HT Biotechnology, Cambridge, UK), 200 U of M-MuLV reverse transcriptase (Finnzymes, Thermo Fisher Scientific, Pittsburgh PA, USA) and 4 U ribonuclease inhibitor (Qiagen, Valencia, CA, USA) according to manufacturer's instructions.

Schematic representation of IGF-1 alternative splicing peptides. Mature IGF-1 consists of exons 3 and 4. At the N-terminal the signal peptides consist of exon 1 or 2 spliced with part of exon 3 and give rise to class 1 or 2 molecules, respectively, whereas at the C-terminal combinations of exon 4 with exon 5 or 6 or both produce the E peptides named Ea, Eb and Ec. Primers used for amplification of alternative IGF-1 transcripts are marked as arrows and are located in their designed complementary exon or exon-exon junction. Forward (sense) primers are presented as right (clockwise) blue arrows, whereas reverse (antisense) primers as left (reverse clockwise) brown arrows (A). The intron-exon spanning or exon-exon junction design of primers and the length of intervening introns (B) eliminates the possibilities of contaminating DNA amplification. IGF-1, Insulin-like growth factor-1.
Quantitative reverse transcription real-time PCR. Equal amounts (20 ng) of the obtained cDNAs were further quantified for the expression of IGF-1 isoforms using 0.4 μM of each primer set described above and 1x Kapa SYBR FAST qPCR kit (KAPA Biosystems, Wilmington, MA, USA) in the same cycling conditions (95°C for 30 s, 95°C for 3 s, 63°C for 30 s, for 40 cycles) for all transcripts tested. The levels of the different variants of transcripts were assessed by calculating the threshold cycle (Ct) as the number of cycles at which the measured fluorescence exceeded the threshold for detection, which was automatically set by the Bio-Rad iQ5 Real-Time PCR Detection System (Hercules, CA, USA) software. Each sample was analyzed in triplicate and the resulting data were averaged. A melting curve (Tm) analysis was also generated by the system's software following the final cycle (40) for each experimental sample by continuous monitoring the SYBR green fluorescence throughout the temperature ramp from 65° to 95°C. QPCR products were verified after both melting curve analysis and agarose gel electophoresis prior to relative quantification. Relative differences between IGF-1 variant expressions after hormonal treatments and control conditions in PC3 cells were estimated as fold changes using the ddCT method. Differences in expression levels of IGF-1 isoforms within cell lines used were performed by comparing the mean±standard deviation (SD) of individual difference between threshold cycle values (dCT). In all cases β-actin (FWD: CCTCGCCTTTGCCGA, RVS: TGGTGCCTGGGGCG) served as an internal control.

Efficiencies and specificity of primers used for quantification of IGF-1 variants. Efficiency of primers' amplification used in the qRT-PCR assay was checked using serial dilutions of cDNA obtained from human muscle tissue and was calculated from the slopes of standard curves. All primer sets were found to have acceptable efficiencies between 90% and 120%, e.g., calculated efficiencies for Class 1 Ea and Eb correspond to percentages of 106 and 101 (SYBR and SYBR1), whereas for IGF-1Ea, IGF-1Eb and Ec peptide to 90, 119 and 100, respectively (SYBR2-4) (A). Following melting curve analysis, qRT-PCR products (e.g., Ec peptide) were further verified by gel electrophoresis, where only one band was observed corresponding to the predicted length (B). The possibility of potentially contaminating DNA amplification was checked in a boosted PCR of 45 cycles using the designed primers (e.g., IGF-1Ea, IGF-1Eb, Ec peptide) or the primers used for amplification of β-actin, which served as internal control for transcripts quantification and 300 ng of genomic DNA as a template. Products were analyzed electrophoreticaly, where no bands were observed, using as positive control cyclophilin. (C). qRT-PCR: Quantitative real-time-polymerase chain reaction.
Statistical analysis. Unpaired t-test and/or regression analysis were used for the fold changes or the dCTs values given and a p-value <0.05 was considered as significant.
Results
Identification of distinct IGF-1 isoform expression patterns unique for each cell line under control conditions. Cell lines were cultured under their individual suggested control conditions and examined for their mRNA levels of IGF-1 isoforms as described in Materials and Methods. The characterization of several human cancer cell lines concerning the expression pattern of IGF-1 Pro- and E-peptides is shown in Figure 4A. More specifically, the KLE and SK-MEL28 cell lines were found to have the maximum expression of pro-IGF1-Ea and -Eb peptides, respectively. Additionally, KLE and MEL28 cells, along with DLD1 and LnCap, were found to have elevated levels of both IGF-1Ea and IGF-1Eb isoforms, compared to others. The Ec peptide was also found at maximum levels in KLE cells, with LnCap following, whereas MCF7 cells presented the lowest expression compared to the rest of tested cell lines. All three isoforms were identified in both HLE-B3 and MG63 cells under control conditions, in contrast to previously reported results by our team under the particularly experimental conditions (10, 13), probably due to the increased sensitivity of our new assay. Very interestingly, the IGF-1Eb isoform was never identified in both estrogen receptor (ER)-positive or -negative breast cancer cell lines MCF7 and MDA-MB-231, respectively, under our experimental conditions.
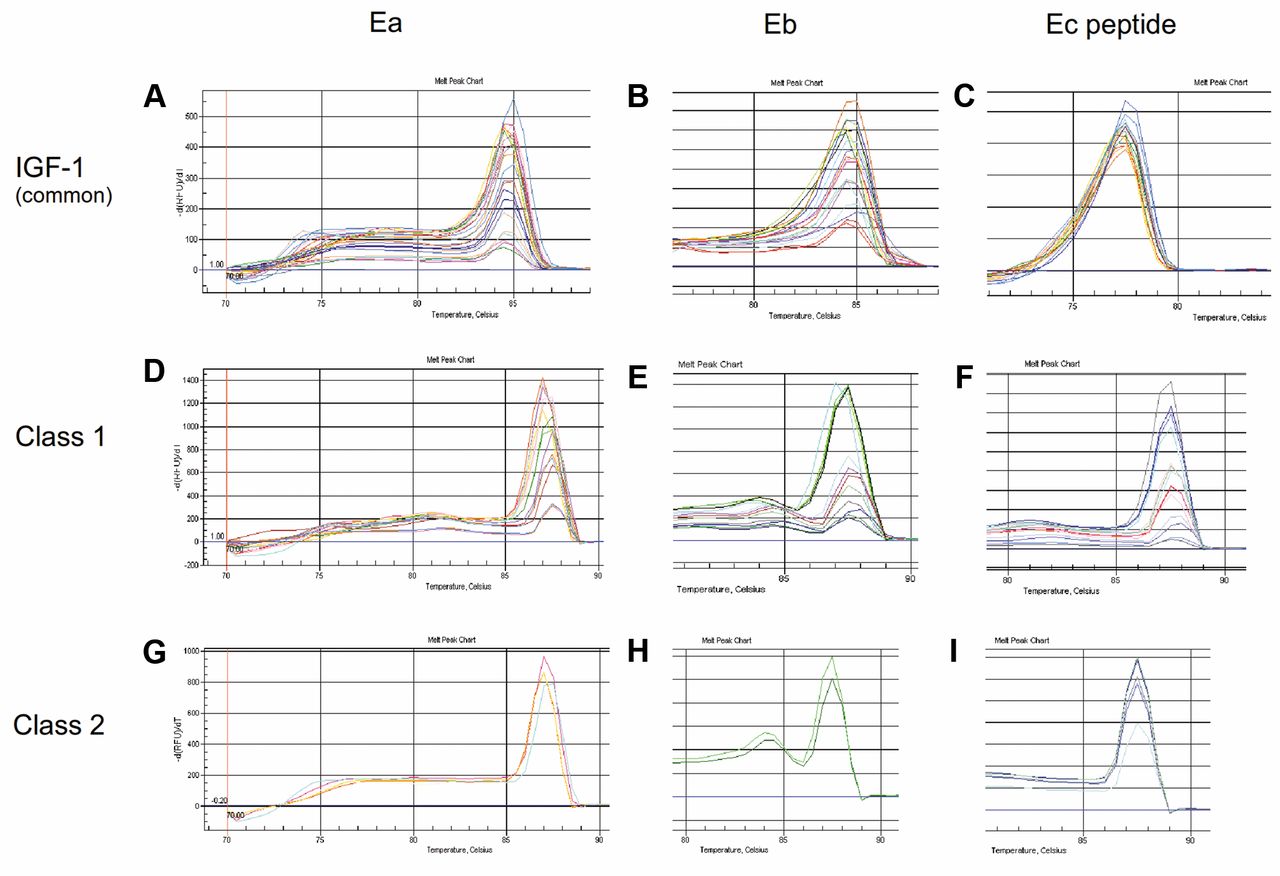
The melting curve analysis of the distinct IGF-1 transcripts. Following qRT-PCR's last cycle (40), a melting curve analysis was generated by the system's software measuring SYBR green fluorescence throughout a temperature ramp from 65°C to 95°C. Analysis showed a unique curve pick in specific for each transcript temperature, indicating the existence of a single product in each qRT-PCR reaction either for IGF-1Ea and IGF-Eb isofroms (A, B) or Ec peptide (C), as well as for class 1 Ea, Eb and Ec peptide (D-F) or class 2 Ea, Eb and Ec peptide molecules (G-I). qRT-PCR: Quantitative reverse transcription-polymerase chain reaction; IGF-1, insulin-like growth factor-1.
Identification of IGF-1 isoform origin; the prevalence of class-1 peptides. We next wondered about the transcription initiation sites of IGF-1 peptides located either on exon 1 or 2 using specific primers, as described, for class 1 or 2 molecules, respectively. The levels of class 1 or 2 originated peptides are shown in Figure 4B and C, although for some cell lines amplification of classes' transcripts was not possible, probably due to the extended length of some amplicons. Most isoforms are found to be mainly and/or exclusively transcribed under the control of exon1 promoter, especially those regarding the IGF1-Eb and Ec peptide. Interestingly, IGF-1Ea and IGF-1Eb isoforms were found to be strictly originated from exon 2 in HeLa and MG63 cells, respectively. Both promoters, located either to exon 1 or exon 2, were found to contribute equally to the transcription of all three isoforms in DLD-1 cells, whereas class 2 served as the predominant origin for all three peptides only in KLE cells.
E2 and dexa modulated pro-IGF1-Ea and Eb expression in PC3 cells. We have previously addressed the expression pattern of pro-IGF1-Ea, -Eb and -Ec in PC3 cells under control conditions (11). Since PC3 cells are androgen receptor-negative cells we tested the potentially hormonal regulation of IGF-1Ea, IGF-1Eb and Ec peptide expression using 10 or 100 nM E2 or Dexa for 24 or 48 h in PC3 cells. Indeed, the expression of IGF1-Ea and IGF-1Eb isoforms was significantly increased, namely Ea was up-regulated up to10-fold and Eb up to 4-fold, compared to control, by 10 nM E2 after a 24-h exposure (p=0.025 and p=0.034, respectively), whereas 100 nM of Dexa for 48 h significantly decreased the IGF-1Ea and Eb expression (p=0.031 and p=0.042, respectively). One hundred nM of E2 for 24 h or 10 nM of Dexa for 48 h slightly modulated Ea and Eb expression levels compared to control. Since the last 24 aa of the carboxyl (-C) terminal of IGF-1Ec isoform have been suggested to have distinct bioactive implications in prostate cancer (14), we analyzed its regulation at the mRNA level in response to several factors in PC3 cells (Figure 4B). The majority of these factors (E2 or Dexa at 10 or 100 nM for 24 or 48 h) decreased the Ec peptide in all time and dose conditions tested. A significant reduction was observed after 10 or 100 nM E2 for 24 h (p=0.048 and p=0.044, respectively) and 100 nM Dexa after 48 h of exposure (p=0.032). An increase of Ec peptide expression was noticed by 10 nM of synthetic GH for 48 h, however, not a significant one. For comparison reasons and using the fold change formula, in all cases the expression level of each IGF-1 isoform in PC3 cells under control conditions served as fold 1.

Levels of distinct IGF-1 transcripts expression. Because greater Ct values correspond to smaller expression, levels of relative expression were calculated after reversing the difference between the Ct of target gene and reference gene (β-actin) for each cell line (1/Dct). All samples were measured in triplicates and the values were averaged, whereas Ct values above 35 were excluded from analysis. The comparison of IGF-1 variants' expression between distinct cell lines revealed that the IGF-1Ea and Eb isoforms are expressed in greater levels in KLE, MEL28, DLD1 and LnCap cells. Additionally, the Ec peptide was found at maximum levels also in KLE and LnCap, whereas MCF7 cells showed the lowest expression. Interestingly, IGF-1Eb was completely lacking, under our experimental conditions, from both ER-positive or -negative MCF7 or MB231 breast cancer cells (A). For the majority of cell lines used, we were able to determine the classes origin of IGF-1 transcripts. Class 1 proved as the predominant origin of IGF-1 transcripts for most cell lines, especially for IGF-1Eb and Ec peptide variants (B), whereas, in some cases, exon 2 served as the only transcription initiation site (C). Ct, Threshold cycle; IGF-1, insulin-like growth factor-1.
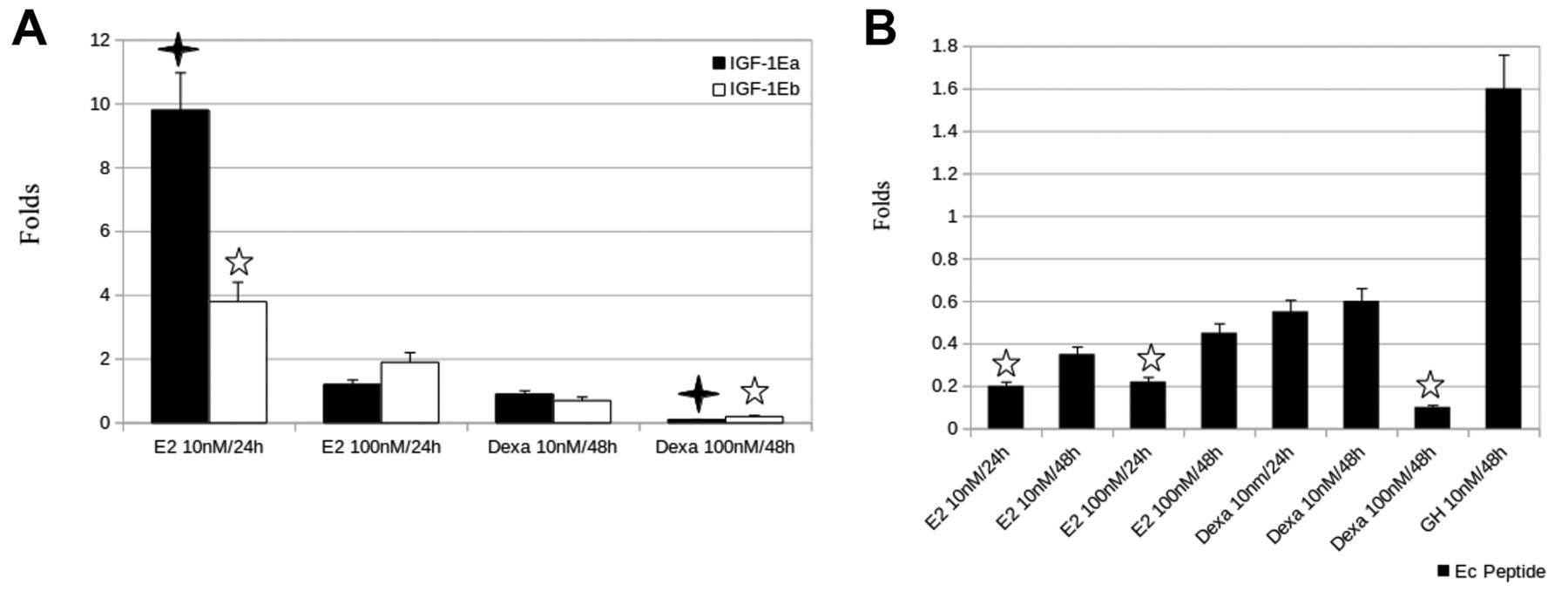
Hormonal regulation of IGF-1 transcripts in PC3 cells. Androgen receptor-negative prostate cancer PC3 cells were treated with 10 or 100 nM of E2, Dexa or GH for 24 or 48 h of exposure, before harvesting of cells and RNA extraction. Control cells treated with placebos for 24 or 48 h, as appropriately, and levels of each IGF-1 transcript following qRT-PCR analysis served as fold 1. Results are presented as fold changes and indicate a significant up-regulation of IGF-1Ea and IGF-1Eb isoforms by 10 nM E2 for 24 h (p=0.025 and p=0.034, respectively), whereas 100 nM of Dexa for 48 h significantly reduced both (p=0.031 and p=0.042, respectively), when compared to control (A). The Ec peptide was negatively regulated by the majority of treatment conditions used; 10 or 100 nM E2 for 24 h and 100 nM Dexa for 48 h significantly down-regulated the Ec peptide (p=0.048, p=0.044 and p=0.032, respectively). A small induction in Ec peptide expression was observed only after addition of 10 nM GH for 48 h (B). E2, Estradiol; Dexa, dexamethsone; GH, growth hormone; IGF-1, insulin-like growth factor-1; qRT-PCR: quantitative real-time-polymerase chain reaction.
Discussion
In this study we address, for the first time, the differential expression pattern of IGF-1 isoforms, including pro-, classes and E peptides, in a large number of human cancer cell lines under control and treatment conditions using a novel qPCR assay. The distinct IGF-1 transcript patterns presented herein raises the possibility for a unique role of each isoform, depending on cancer type.
It has been long enough since the scientific community started arguing about the existence and the biological activity of IGF-1 pro- and E peptides instead of the established mature IGF-1. Over the last years C-terminal E peptides have proved a major point of argument regarding their biological roles, whereas pro-IGF-1 peptides present a higher acceptance among scientists for their pathophysiological actions, probably due to the fact that they contain the mature IGF-1 sequence. Among others, our team has previously addressed unique roles for IGF-1 alternative transcripts, especially for the Ec peptide, which notably has been suggested to have IGF-1-independent actions (7-11). Unique roles have also been addressed, by independent teams, for IGF-1Ea, IGF-1Eb and IGF-1Ec isoforms in several pathophysiological conditions, including cancer (15-24). These results, along with the data presented herein, further support the notion for distinct roles of IGF-1 alternative transcripts and implicate IGF-1 isoforms in human cancer biology, in contrast to other studies suggesting a bioactivity only for the mature IGF-1 (25, 26).
The characterization of cells lines under control conditions for IGF-1 transcripts revealed some noticeable results. First, endometrial cancer KLE cells proved to have the highest levels of IGF-1Ea and Ec peptide among all cell lines tested. Although we have previously addressed that KLE cells express all three pro-peptides (9), the direct comparison with the other cancer cells could raise the biological significance of Ea and Ec peptide in endometrial malignancy. Second, the complete lack, in our hands, of IGF-1Eb peptide in breast cancer cell lines independently of estrogen status in contrast to all other cell lines where was indeed present, could reflect a unique and probably protective effect of this variant in breast cancer progression. Additionally, estrogen-sensitive MCF7 cells were found to express the minimum levels of Ec peptide. Except cancer type and estrogen status, many other factors could also contribute for the observed IGF-1 variant expression patterns, such as the existence of viral proteins (12).
Very interestingly and for certain cell lines, we were able to determine the transcription initiation site, located either on exon 1 or 2, for the expressed IGF-1 isoforms. The importance of this finding is fortified by the limited knowledge on the potentially distinct roles of classes. We address here that class 1 transcripts are predominant in the vast majority of the cell lines used. An explanation could be the finding that class 1 transcripts may have higher transcription efficiencies, whereas class 2 transcripts could be more stable (27). Additionally, and pursuant to our results, showing a prevalence of transcripts containing exon 1, class 2 peptides were suggested to have mainly endocrine actions, whereas class 1 mostly autocrine/paracrine ones (28). Class 2 transcripts have been found to be predominant only in KLE cells. Taken together, the KLE cell line seems to present a unique pattern of IGF-1 isoforms raising, thus, the need for further studies in order to determine the importance of IGF-1 alternative transcripts in endometrial cancer pathology.
Less is also known about the response of IGF-1 variants to hormonal treatments. For these experiments we used androgen receptor-negative prostate cancer cells (PC3), where the IGF-1 isoforms expression is well-addressed (11). We found a significant regulation (up or down) of pro peptides, Ea and Eb, depending on dose and time of E2 or Dexa used. Noticeable, the Ec peptide was down-regulated almost in all hormonal conditions tested, further supporting the notion for a distinct biological significance of this molecule in prostate cancer (11, 14). We found a small induction of Ec peptide expression only after GH treatment in PC3 cells, although in normal human tissues, such as the skeletal muscle, the up-regulation at the mRNA level of all IGF-1 isoforms following GH treatment was not evident (29).
One limitation of our study is the difficulty to compare the relative expression of IGF-1 isoforms between so many cell lines, using the fold change formula due to the differential molecular background represented by each one, as well as between the isoforms themselves due to the variant efficiency of the different pairs of primers used for each transcript (Figure 3A). For that reason we used the reversed Dct formula (1/Dct) as a more suitable approach. We also failed to efficiently amplify class 1 or 2 transcripts in some cell lines probably because the length of class 1 and 2 amplicons (Table I) were above the efficiency limits of the SYBR green assay, thus, a probes' assay design could probably be proved more suitable for the distinct identification of the IGF-1 classes. The disparities of the results presented herein, for some cell lines compared to previous published results by our team, could possibly be explained by the increased sensitivity of our new assay.
To conclude, this manuscript addresses the development of a specific, sensitive and cost effective tool for the identification of almost all IGF-1 alternative transcripts. The differential expression pattern in the cell lines used, which served as models of several human malignancies, further implicate the IGF-1 isoforms in human cancer biology. The unique pattern, which was revealed for some cancer models, such as endometrial and breast malignancies, highlight the importance of understanding the exact molecular function of the IGF-1 isoforms in these and other cancerous cases. In addition, our results may also contribute to the importance and notion for a unique and distinct role of the Ec peptide. Further studies at the mRNA and protein level, as well as determination of IGF-1 isoforms' bioactivities, are warranted to confirm our findings and define the role of these molecules in cancer.
- Received September 19, 2014.
- Revision received October 15, 2014.
- Accepted October 21, 2014.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Expression of IGF-IEc Isoform in Renal Cell Carcinoma Tissues
- IGF-IEc Expression Is Associated With Advanced Differentiated Thyroid Cancer
- Differential Expression of IGF-I Transcripts in Bladder Cancer
- The COOH-terminus of the IGF-1Ec Isoform Enhances the Proliferation and Migration of Human MCF-7 Breast Cancer Cells
- The Role of Insulin-like Growth Factor-1 Signaling Pathways in Uterine Leiomyoma