


Abstract
A report that effects of butyrate on some cells may be mediated by activation of AMP-activated protein kinase (AMPK) prompted this study which examines if other AMPK activators can induce differentiation and inhibit proliferation of colon cancer cells in a manner similar to butyrate. Using induction of alkaline phosphatase as a marker, it was observed that compound C, an AMPK inhibitor, is able to reduce the differentiating effect of butyrate on SW1116 and Caco-2 colon cancer cells. Metformin was observed to be less effective than butyrate in the induction of alkaline phosphatase but was more effective as a growth inhibitor. Phenformin was found to be a more potent growth inhibitor than metformin and both compounds cause acidification of the medium when incubated with colon cancer cells. Combined incubation of 2-deoxyglucose with either of the biguanides prevented the acidification of the medium but enhanced the growth inhibitory effects.
Differentiation of colon cancer cells in culture is favored by incubation with butyrate. This simple four-carbon fatty acid has a variety of effects on cancer cells but most attention has been directed to its action as an inhibitor of histone deacetylase activity. A report that butyrate may activate AMP-activated protein kinase (AMPK) in Caco-2 colon cancer cells (1) prompted the current investigation of whether AMPK activation by other agents will promote differentiation and growth inhibition in colon cancer cells. One such possible agent is metformin, a compound widely used in the treatment of type 2 diabetes. Metformin appears to be an indirect activator of AMPK (2, 3). Lactic acidosis is a possible side-effect of treatment with metformin and it was cases of lactic acidosis that led to the withdrawal of the related biguanide, phenformin. Like metformin, phenformin has been reported to activate AMPK (4) and to inhibit proliferation of some tumor cells (5). This report addresses the potential growth-inhibitory and -differentiating effects of metformin and phenformin in colon cancer cells. Possible effects mediated through activation of AMPK were tested by combined treatment with compound C, an inhibitor of AMPK (6).
Cancer cells tend to have increased glycolytic activity. Among the compounds that have been examined as inhibitors of glycolysis is 2-deoxyglucose. The combination of metformin and 2-deoxyglucose has been found to induce apoptosis in prostate cancer cells (2). This study aimed to examine this combination of drugs with colon cancer cells and to determine if additive effects would also be seen with the combination of phenformin and 2-deoxyglucose.
Materials and Methods
Cells and determination of cell proliferation. SW1116, HT29, and Caco-2 human colon cancer cells were obtained from American Type Culture Collection, Rockville, MD, USA, and were incubated at 37°C in RPMI-1640 medium with 5% fetal calf serum and 25 mM N-(2-hydroxyethyl)piperazine-N’-(2-ethanesulfonic acid) buffer. Of these cell lines, the HT29 cells exhibit the most rapid proliferation and the Caco-2 cells show the greatest tendency to differentiate including spontaneous differentiation when the cells are cultured for longer times than were examined in the present work. The incorporation of [3H]thymidine into DNA was measured after incubating cells for 2 h with 2 microcuries [3H]thymidine as previously described (7). Cell proliferation was also monitored using the WST-1 reagent according to the directions of the supplier, Roche Diagnostics, Mannheim, Germany. Incubations with the reagent were for 4 h and the difference in absorbance at 450 and 695 nm was recorded.

Effects of incubation of Caco-2 cells (A) and SW1116 cells (B) for 72 hours with 0.5 mM butyrate and 10 μM compound C on alkaline phosphatase activity. The data are expressed as the means and standard deviations for 3 determinations.
Reagents. Enzyme substrates, butyrate, compound C, 2-deoxyglucose, metformin and phenformin were obtained from Sigma-Aldrich, St. Louis, MO, USA. A769662 was obtained from LC Laboratories, Woburn, MA, USA.
Enzyme assays. A total of 1.5×106 cells were incubated with 10 ml RPMI-1640 medium with 25 mM HEPES and 5% fetal calf serum. The medium was changed after 24 h and unless stated otherwise the cells were incubated for 72 h before harvesting. The cells were washed with phosphate-buffered saline and extracted with 0.5% NP40, 0.25 M NaCl, 5 mM EDTA and 50 mM Tris pH 8.0. The protein concentration of the extract was determined using the BCA Protein Assay Reagent from Pierce, Rockford, IL, USA. Enzymes were assayed at 37°C as previously described (8). Alkaline phosphatase was assayed using para-nitrophenyl phosphate as substrate. Formation of product was monitored by the change in absorbance at 410 nm.
pH determination. At the end of the incubation, the medium was aspirated and centrifuged to remove suspended cells. pH was determined with an electrode and was found to correlate well with changes in the light absorbance at 560 nm reflecting changes in the pH indicator, phenol red.

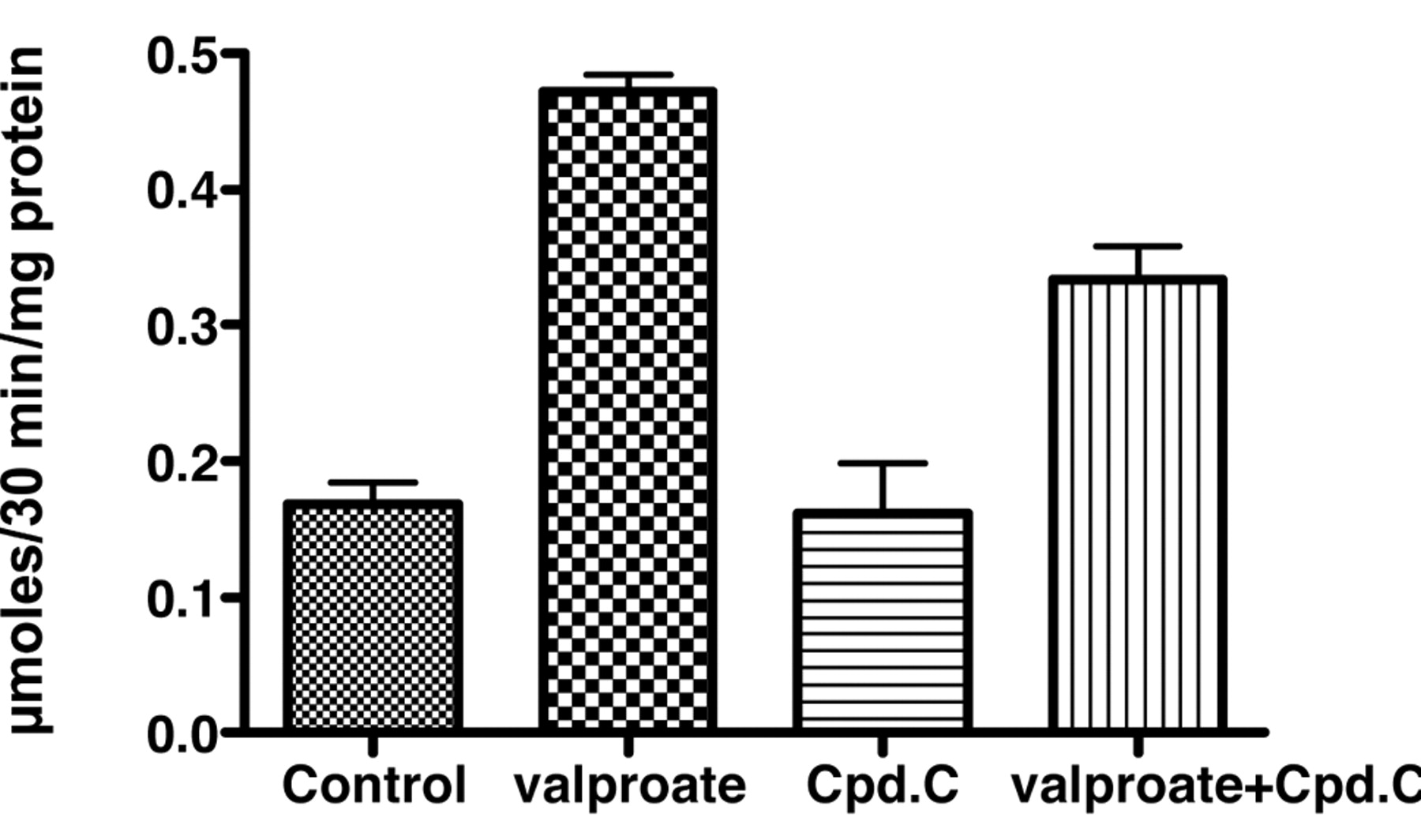
Effects of incubation of Caco-2 cells for 72 hours with 1.0 mM valproate and 10 μM compound C on alkaline phosphatase activity. The data are expressed as the means and standard deviations for 3 determinations.
Statistical evaluation. Statistical significance of the results was determined by a two-tailed Student's t-test or by Dunnett's test for multiple comparisons using the Instat program from GraphPad Software, Inc., La Jolla, CA, USA. A probability of less than 5% was considered significant.
Results
Incubation of Caco-2 cells with 10 μM compound C for 72 h was accompanied by alkaline phosphatase activity that was somewhat higher than in controls, but in combination with 0.5 mM butyrate there was less induction of alkaline phosphatase than with butyrate alone (Figure 1A). In SW1116 cells, 10 μM compound C essentially blocked the induction of alkaline phosphatase activity by butyrate (Figure 1B). A decrease in the induction of alkaline phosphatase activity in Caco-2 cells was also observed after coincubation with compound C and another histone deacetylase inhibitor, valproate (Figure 2).
Substantial induction of alkaline phosphatase activity comparable to that caused by butyrate was not seen with compounds reported to be activators of AMPK. The data in Figure 3 illustrate this point for the compound A769662, which did not have a significant effect as a single agent and did not significantly affect the response to butyrate. In studies with metformin (1 mM) and phenformin (0.1 mM), a consistent significant effect on alkaline phosphatase activity was not observed after 72 h incubations with Caco-2 cells (data not shown). Decreases in protein yield were seen, suggesting an inhibition of growth and changes in the color of the phenol red pH indicator suggested an acidification of the medium. A decrease in pH was further documented after incubation of 3 colon cancer cell lines with phenformin for 72 hours (Table I). The acidification of the medium was countered by coincubating with phenformin and either compound C or 2-deoxyglucose. The observed effect on the color of the medium was monitored by measuring the absorbance of the medium at 560 nm (Table II). The readings correlated with the pH measurements using a pH meter and again indicated that the acidification of the medium caused by incubating the cells with phenformin was blocked more effectively by 2-deoxyglucose than by compound C.

Effects of incubation of Caco-2 cells for 72 hours with 0.5 mM butyrate and 100 μM A769662 on alkaline phosphatase activity. The data are expressed as the means and standard deviations for 3 determinations.
A significant increase in alkaline phosphatase activity in Caco-2 cells was not seen after incubation with either metformin or phenformin. However, in SW1116 cells, small but significant increases were observed. This is illustrated for phenformin in Figure 4. The increase in enzyme activity was blocked when phenformin was coincubated with compound C.
Although phenformin and 2-deoxyglucose as single agents did not have significant effects on alkaline phosphatase activity in Caco-2 cells, the combination did result in a significant increase in alkaline phosphatase activity (Figure 5A). At the same time there was a significant decrease in the protein yield when the Caco-2 cells were incubated with a combination of phenformin and 2-deoxyglucose (Figure 5B). This suggested that the combination reduced cell growth and this conclusion was supported by data for the incorporation of [3H]thymidine into DNA (Figure 6). The data in Figure 7 suggested that a combination of metformin and 2-deoxyglucose was more potent for the inhibition of [3H]thymidine incorporation into DNA of SW1116 cells than either compound as a single agent. Using reduction of a tetrazolium salt as an alternative measure of cell proliferation, the data in Figure 8 indicate that at a concentration of 1 mM, phenformin was a more active inhibitor of growth than metformin in the HT29 and Caco-2 human colon cancer cell lines. In these two cell lines, the effect of the biguanides was enhanced by co-incubation with 2-deoxyglucose. The assay procedure indicated an increase of tetrazolium salt reduction when SW1116 cells were incubated with either metformin or phenformin, but this effect was blocked when there was a combined incubation with 2-deoxyglucose.
Effects of phenformin, compound C and 2-deoxyglucose on the pH of the medium after incubation with colon cancer cells for 72 hours.
Effects of phenformin, compound C and 2-deoxyglucose on the absorbance of the medium at 560 nm after incubation with colon cancer cells for 72 hours.
Discussion
Butyrate is a short-chain fatty acid that inhibits most types of histone deacetylase activity and causes differentiation of several types of cancer cell. Its differentiating action on colon cancer cells can be monitored by the induction of alkaline phosphatase activity. There is evidence to suggest that some actions of butyrate may be mediated through activation of AMPK (1, 9). Peng et al. found that treatment of Caco-2 colon cancer cells with butyrate resulted in an activation of AMPK and that compound C, an AMPK inhibitor, could block some of the effects of butyrate on Caco-2 cells (1). This study investigated whether compound C can block the action of butyrate on the induction of alkaline phosphatase and whether some activators of AMPK can mimic the action of butyrate in colon cancer cells. It was found that induction of alkaline phosphatase by butyrate was inhibited by compound C more effectively in SW1116 cells than Caco-2 cells. On the other hand, activators of AMPK including A769662, metformin and phenformin, had at best only weak activity for the induction of alkaline phosphatase. The greatest response was seen with SW1116 cells incubated with phenformin and the effect was blocked by co-incubation with compound C. Together the data suggest that activation of AMPK may be a factor in the induction of alkaline phosphatase by butyrate but the response of the enzyme activity to butyrate is greater than that seen with some more established activators of AMPK.

Effects of incubation of SW116 cells for 72 hours with 0.1 mM phenformin and 10 μM compound C on alkaline phosphatase activity. The data are expressed as the means and standard deviations for 3 determinations.

Effects of incubation of Caco-2 cells for 72 hours with 0.1 mM phenformin and 1.0 mM 2-deoxyglucose on (A) alkaline phosphatase activity and (B) protein yield. The data are expressed as the means and standard deviations for 3 determinations.
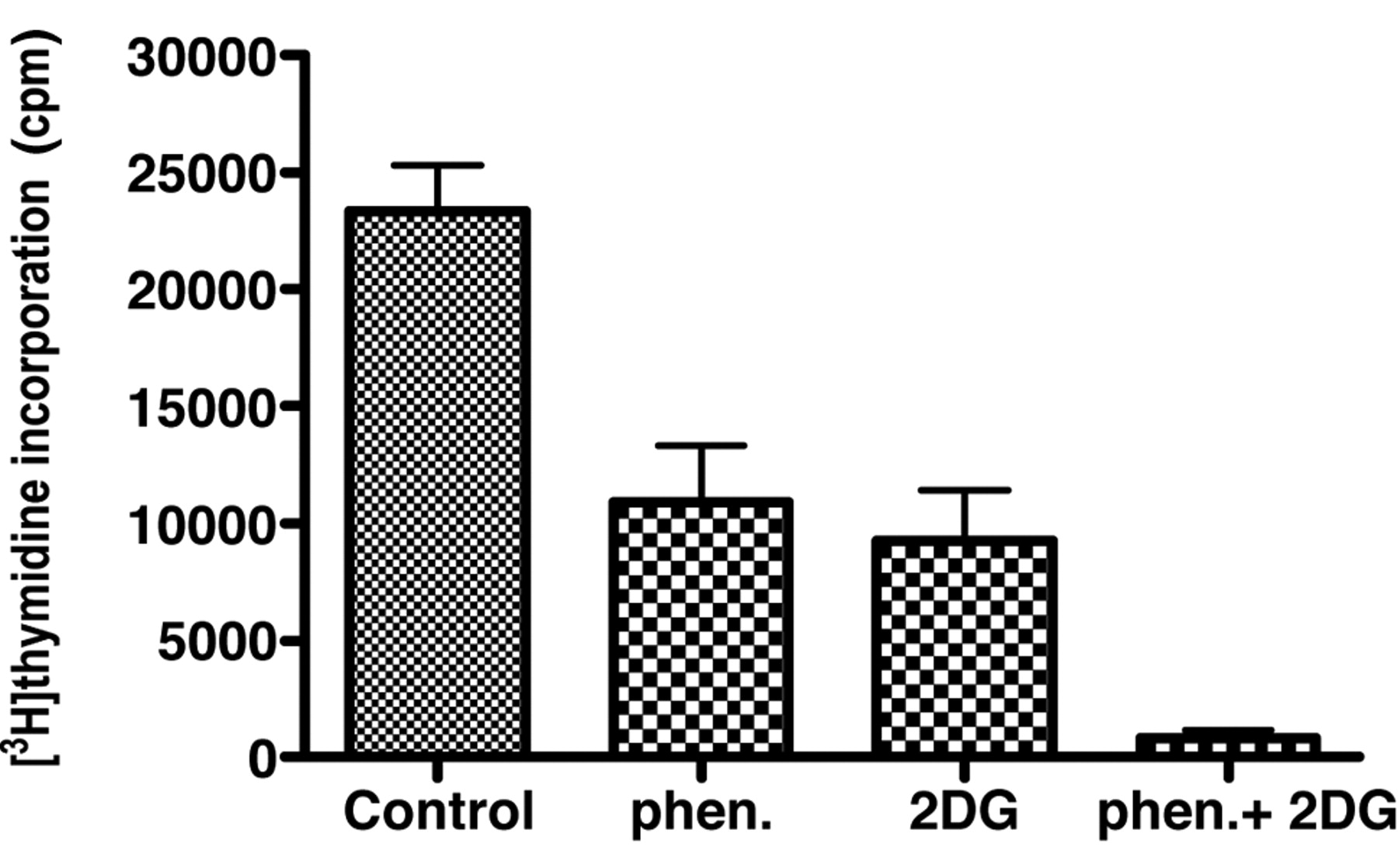
Effects of incubation of Caco-2 cells for 72 hours with 0.1 mM phenformin and 1.0 mM 2-deoxyglucose on the incorporation of [3H]thymidine into DNA. The data are expressed as the means and standard deviations for 6 determinations.

Effects of incubation of SW1116 cells for 24 hours with 1.0 mM metformin and 10.0 mM 2-deoxyglucose on the incorporation of [3H]thymidine into DNA. The data are expressed as means and standard deviations for 6 determinations.
In the course of these studies with metformin and phenformin, the acidification of the medium when these biguanides were incubated with colon cancer cells was apparent from the change in color of the phenol red indicator. Since cases of lactic acidosis had resulted in the withdrawal of phenformin as a treatment for type 2 diabetes and the possibility of lactic acidosis has been of some concern in the use of metformin (10-12), compounds that might block acidification of the cells were sought. One such agent was compound C on the basis of the previously described studies and another was 2-deoxyglucose, which is an inhibitor of glycolysis and is known to be taken up well by many cancer cells. Both these molecules diminished the acidification of the medium, and in the three colon cancer cell lines that were examined, the most consistently effective compound blocking acidification was 2-deoxyglucose. A further advantage of the combination treatment with phenformin and 2-deoxyglucose was an enhancement of the inhibition of cellular proliferation as judged by [3H]thymidine incorporation into DNA and an assay measuring tetrazolium salt reduction.


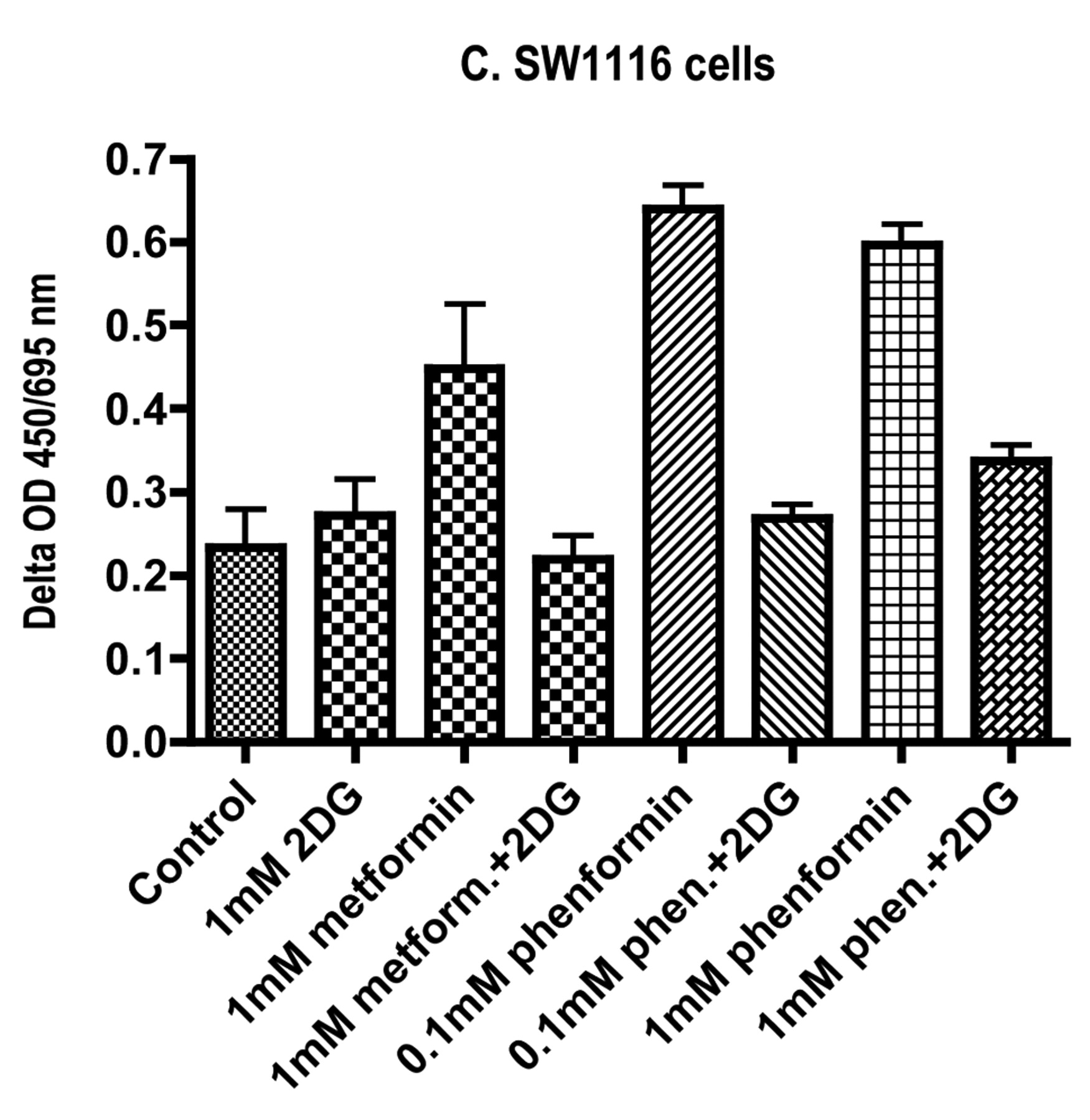
Effects of incubation of (A) HT29 cells, (B) Caco-2 cells and (C) SW1116 cells for 72 hours with metformin, phenformin and 2-deoxyglucose (2DG) on tetrazolium salt reduction measured by the difference in light absorbance at 450 and 695 nm. The data are expressed as the means and standard deviations for 8 determinations.
The widespread use of tetrazolium salt reduction assays as measures of cell viability or proliferation is largely because they are convenient and lend themselves to multiple determinations using 96-well plates. The results are often interpreted in terms of mitochondrial activity but there is evidence to suggest that the activities of cell surface NADH oxidoreductases may be important determinants (13). Inconsistency with other measures of cell viability and proliferation has been noted particularly when studying redox active reagents (14, 15). In the studies reported here, results were more consistent with data from protein assays and thymidine incorporation for the Caco-2 and HT29 cells than for the SW1116 cells. In the case of the SW1116 cells, the data suggested that incubation with metformin or phenformin as single agents was associated with increased values while the values for protein synthesis and thymidine incorporation indicated growth-inhibitory effects. The factors underlying this discrepancy remain to be established. However, all the assays indicated that the co-incubation of the biguanides with 2-deoxyglucose enhanced the growth inhibition in Caco-2, HT29 and SW1116 cells relative to values obtained with single agents.
Studies on biguanides as potential anticancer agents have focused largely on metformin (3, 16-22). Phenformin is a drug that was abandoned in clinical usage over 30 years ago. It has been previously reported to have growth-inhibitory properties when incubated with cancer cells (5). Growth inhibition in colon cancer cells has been described for metformin (23) and the combination of metformin and 2-deoxyglucose induced p53-dependent apoptosis in prostate cancer cells (2). Phenformin is inhibitory for growth at lower concentrations than metformin and, in combination with 2-deoxyglucose, it may be possible to avoid the lactic acidosis that was problematic for treatment of diabetes. Perhaps phenformin should be re-examined as a potential agent in cancer therapy.
Acknowledgements
This research was supported by a grant from the Alma Toorock Memorial for Cancer Research.
- Received December 31, 2010.
- Revision received January 23, 2011.
- Accepted January 25, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Phenformin Induces Caspase-dependent Apoptosis of FaDu Head and Neck Squamous Cell Carcinoma Cells
- Effects of Biguanides on Growth and Glycolysis of Bladder and Colon Cancer Cells
- Inhibition of Growth by Combined Treatment with Inhibitors of Lactate Dehydrogenase and either Phenformin or Inhibitors of 6-Phosphofructo-2-kinase/Fructose-2,6-bisphosphatase 3
- Inhibition of Growth of Bladder Cancer Cells by 3-(3-Pyridinyl)-1-(4-pyridinyl)-2-propen-1-one in Combination with Other Compounds Affecting Glucose Metabolism
- Molecular Pathways: Preclinical Models and Clinical Trials with Metformin in Breast Cancer
- A Cohort Study of Metformin Exposure and Survival in Patients with Stage I-III Colorectal Cancer
- Regulation of the Proliferation of Colon Cancer Cells by Compounds that Affect Glycolysis, Including 3-Bromopyruvate, 2-Deoxyglucose and Biguanides