


Abstract
Background: Prostate cancer is characterized by a low rate of glycolysis and glucose uptake. We hypothesize that fatty acid is dominant over glucose in uptake by prostate cells. Materials and Methods: One benign (RWPE1) and two malignant (LNCaP and PC3) prostate cell lines were assayed for their in vitro uptake of radiolabeled glucose analogs 3H-fluoro-2-deoxyglucose and 18F-fluoro-2-deoxyglucose, and the long-chain fatty acid 3H-palmitic acid. Fractional uptake was standardized to viable cell numbers. Results: Uptake of palmitate in all 3 prostate cell lines was significantly higher than that of glucose at all incubation times (p<0.01). But in malignant cell lines, neither glucose nor palmitate uptake was quantitatively higher than that in the benign cell line. The uptake of fatty acid by prostate cells is a dynamic, active process mediated by the membrane receptors. Conclusion: Prostate cells are characterized by a dominant uptake of fatty acid over glucose, suggesting that future development of new diagnostic and therapeutic approaches in prostatic cancer should focus on fatty acid substrate. Fatty acid imaging may be useful in detection of recurrence and metastasis, but not in differentiating malignant from benign prostate tissue.
Prostate cancer remains a major public health concern. It is the most prevalent cancer in human beings and the second leading cause of male cancer death in the U.S. (1). Initial diagnosis of prostate cancer is relatively straightforward, being based on prostate specific antigen (PSA) screening and confirmatory biopsy, but accurate staging and detection of recurrent and metastatic disease remains clinically challenging.
Metabolic changes during malignant transformation have been noted for many years. Warburg first reported that cancer cells preferentially rely on glycolysis as a major bioenergetic source even in the presence of oxygen and produce high levels of lactate and pyruvate (2). Increased glucose consumption is a basic characteristic of malignant cells and is linked to energy production from glycolysis. Amplified glucose usage by malignant cells therefore forms the theoretic basis for cancer detection through uptake of the glucose analog, fluorine-18-labeled 2-deoxy-2-fluoro-D-glucose (18F-FDG) which can be imaged by positron-emission tomography (PET). Today, FDG-PET is widely used for diagnosis, staging, restaging and monitoring therapy of numerous types of malignant diseases.
Unlike most malignancies, prostate cancer is characterized by a slow glycolysis and low FDG avidity on PET imaging (3, 4). Other positron-emitting radiotracers, such as 18F-choline, 11C-choline, 11C-acetate and 18F-fluorodihydrotestosterone have recently been suggested as putative imaging agents for the detection of prostate tumors (5-9). Clinical evaluation of these radiotracers is presently underway and the role of PET scaning in prostate cancer is still evolving (10-12). Cell proliferation in malignant tumors is associated with increased energy requirements. If glucose consumption is not elevated in prostate cancer, alternative metabolic pathways must provide bioenergy for abnormal cell proliferation and growth. We therefore hypothesize that increased fatty acid metabolism characterizes prostate cancer, which depends on fatty acid oxidation, rather than glucose metabolism, as its dominant energy source. To test this hypothesis, as the first step, we investigated the uptake of radiolabeled glucose and fatty acid in various benign and malignant prostate cell lines.
Materials and Methods
Cell culture. Human prostate cancer cell lines LNCaP, PC3 and normal human prostate cell line RWPE1 were purchased from the American Type Culture Collection (Manassas, VA, USA). LNCaP and PC3 cells were grown in RPMI-1640 cell culture medium supplemented with 10% fetal bovine serum, L-glutamine (2 nM), penicillin (100 IU/ml) and streptomycin (100 μg/ml) in 95% air and 5% CO2. RWPE1 cells were grown in keratinocyte serum-free medium supplemented with 5 ng/ml human recombinant EGF, 50 mg/ml bovine pituitary extract and penicillin/streptomycin in the same atmosphere as above.
Incubation study. 3H-Palmitic acid and 3H 2-deoxy-D-glucose were purchased from Sigma-Aldrich, St. Louis, MO, USA. For LNCaP and PC3 cells, culture medium was changed to glucose-free RPMI-1640 one day prior to the uptake experiments. Medium for RWPE1 cells contained no glucose. Medium containing labeled substrate was prepared with glucose-free RPMI-1640 medium for LNCaP/PC3 cells and keratinocyte serum-free medium for RWPE1 cells, containing either 37 kBq/ml of 3H-palmitic acid (specific activity 1.15 TBq/mmol), or 37 kBq/ml 3H 2-deoxy-D-glucose (specific activity 370 GBq/mmol). Original flasks of each cell line were split into palmitate and glucose groups consisting of 2-3 flasks per group. A total 10 ml of labeled medium were added to each of the plamitate and glucose flasks, while plain medium without labeled substrate was added to an additional flask which was used for cell counting and viability measurements. For subsequent experiments, cells were incubated for periods of 30, 60 and 120 minutes. Following incubations, the cells of the flasks were washed with ice-cold phosphate-buffered saline (PBS) and then treated with trypsin-EDTA at 37°C for 5 minutes. Dispersed cells were transferred to 15 ml centrifuge tubes and washed 3 times with ice-cold PBS. A 0.2 ml aliquot of the last PBS supernatant was saved for radioactivity counting to confirm complete removal of extracellular isotope. After the last wash, 1 ml of 3 N NaOH and 1% sodium dodecyl sulfate was added to lyse cell precipitates and 0.2 ml aliquots of the cell lysates were mixed with 5 ml Ecolume scintillation cocktail (MP Biomedicals, Irvine, CA, USA) for β-ray scintillation counting (LS-5000; Beckman, Fullerton, CA, USA). Additionally, aliquots of labeled substrate were pipetted and measured in triplicate for calculating the activity actually added to each flask.
Co-incubation study. 18F-FDG was obtained from Eastern Isotopes (East Orange, NJ, USA). The three cell lines were prepared as described in the section above. Both 370 kBq of 18F-FDG and 370 kBq of 3H-palmitate were added to medium, while plain medium was added to the cell counting flask. After 60 minute incubation, the cells were washed and lysed as above. Radioactivity of aliquots of the cell lysates was determined initially by gamma scintillation counting (using a Packard Cobra Quantum, Downers Grove, IL, USA). One day thereafter, following 18F decay, aliquots of the same samples were counted by beta scintillation counting. Activity of the labeled media was simultaneously assayed for gamma and beta scintillation counting.
Inhibition of uptake of palmitic acid by HgCl2. All three cell lines were assayed as in the studies above following incubation for 0.5 hour and 1 hour with medium containing 3H-palmitate with and without HgCl2 added to a final concentration of 2.5 mM. Details of cell preparation, incubation, washing and lysis as described above.
Cell count and viability. Cell counting and viability determinations were carried out on the non-labeled cell counting sample. Following incubation, the cells were washed and treated with trypsin-EDTA at 37°C for 5 minutes. Culture medium was added to the cells and 0.4% trypan blue solution (Sigma-Aldrich) was added to the cell suspension at a ratio of 1:1. Cells were counted using a hemacytometer; unstained cells were considered viable based on the assumption that viable cells are impermeable to the dye (13).
Data analysis. Each of the experiments was repeated twice except for the HgCl2 inhibition study. The measured radioactivity was standardized to viable cell numbers (106 cells) and expressed as a percentage of total radioactivity added to the medium: % Uptake (per 106 cells)=[(radioactivity in cell compartment)/(total radioactivity added)] ×100. Uptake values for each combination of cell type, substrate, and incubation time were averaged over the sum total number of observations which consisted of the 2-3 flasks per experiment, and the 2 repeat experiments for each combination. Statistical analysis was performed by t-test.
Results
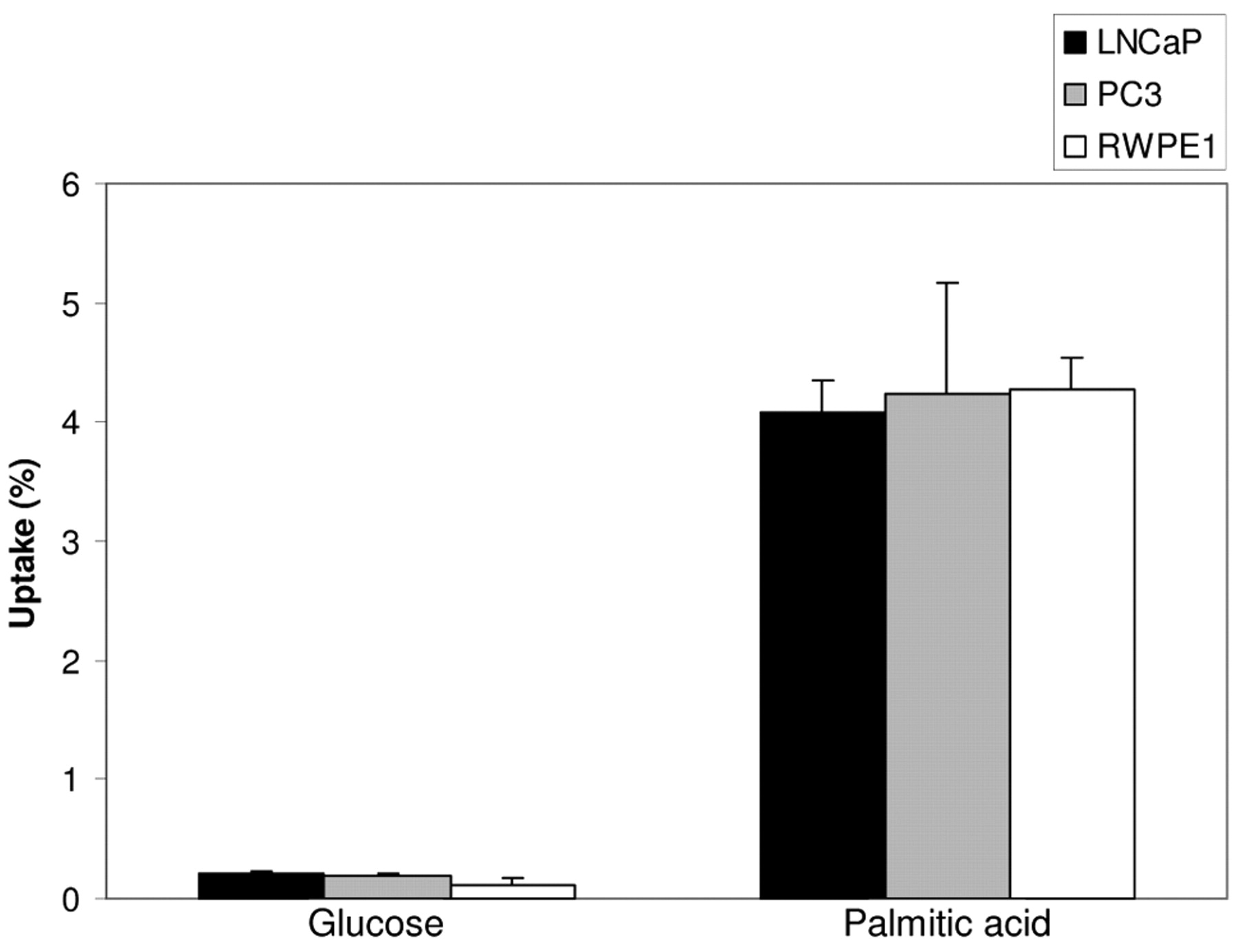
Uptake of 3H-palmitic acid and 3H-glucose in prostate cells. Standard RPMI-1640 medium contains 0.2% glucose. In our experiments, 24 hours prior to the incubation of the cells with radiotracer, the medium of LNCaP and PC3 cells was changed to glucose-free RPMI-1640, which eliminated any possible competitive effect of pre-existing glucose on the uptake of added 3H-glucose. As shown in Figure 1, in all cell lines, uptake of palmitate was at least 10-fold higher than that of glucose. Uptake of palmitate versus glucose following 1-hour incubation was (% uptake per 106 cells): 4.09±0.36 vs. 0.20±0.028 in LNCaP, 4.24±1.31 vs. 0.195±0.007 in PC3 and 4.27±0.37 vs. 0.12±0.084 in RWPE1, with all p-values <0.01. The mean uptake of palmitate and glucose differed significantly in all three cell lines for all incubation times (p<0.01). Upon co-incubation of 18F-FDG and 3H-palmitic acid, under identical cell conditions, uptake of palmitic acid was approximately 20 times higher than that of glucose (Figure 2).
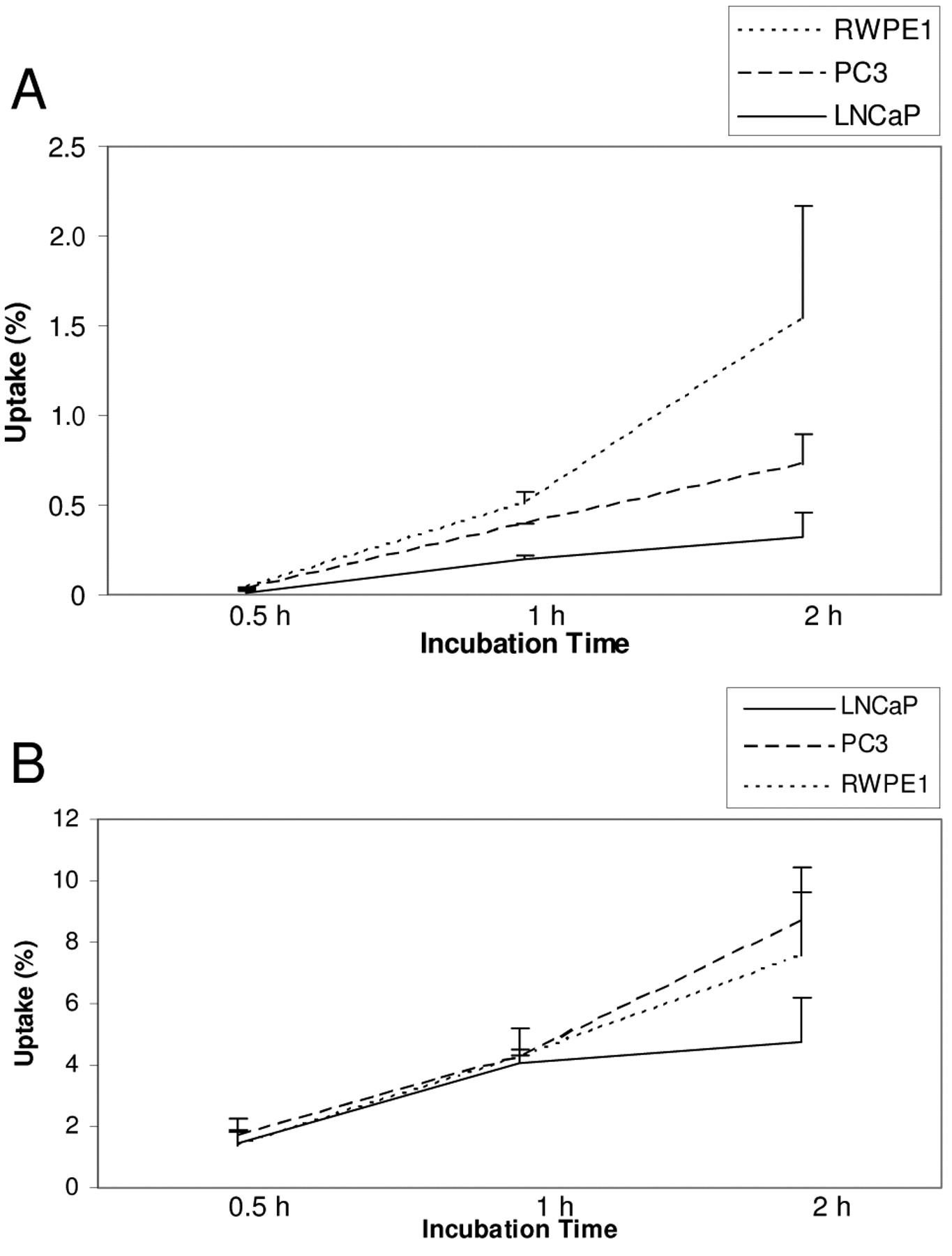
Time course of uptake. Because the significant sample processing required prior to counting of radioactivity precluded measuring multiple samples with variable incubation times in one large experiment, characterization of the time course of uptake was based on data aggregated from three subsequent experiments with variable incubation periods, performed under otherwise identical conditions. As shown in Figure 3, uptake of both glucose and palmitic acid was a time dependent, dynamic process. For all three cell lines, uptake of both glucose and palmitate increased between measurements at 0.5, 1 and 2 hours, but the difference was statistically significant only between 0.5-hour and 1-hour (p-values <0.02). Between the first and second hour, there was a slower, but not statistically significant (p-values >0.05) increase in uptake. It was unclear if saturation was achieved at 2 hours since we did not take any later measurements.
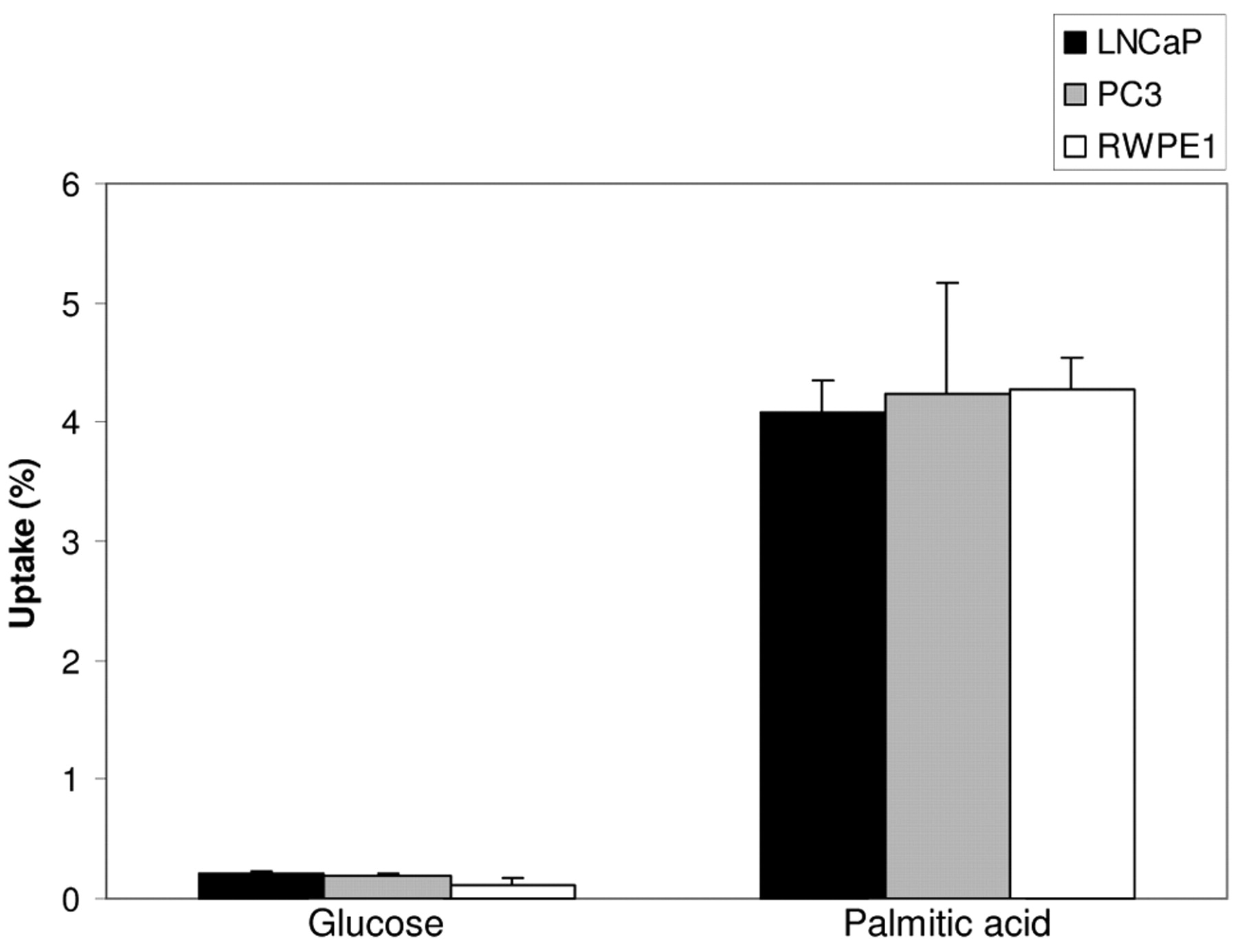
Comparison of uptake between different cells. Contrary to our hypothesis, the two malignant cell lines LNCaP and PC3 did not demonstrate increased uptake of either glucose or palmitic acid compared to normal human prostate cell line RWPE1 (Figure 4). At 0.5 hour and 1 hour post-incubation, the uptake of palmitic acid was similar among all three cell lines; the same was true with respect to glucose uptake. At 2 hours, there was a tendency towards increased uptake of both palmitic acid and glucose in PC3 compared to LNCaP cells, but this difference was not statistically significant. In addition, at all time points, the uptakes of both palmitic acid and glucose were similar in malignant PC3 and normal RWPE1 cells.

Uptake of 3H-glucose and 3H-palmitic acid by prostatic cells incubated in separate flasks after 0.5-hour (A), 1-hour (B) and 2-hour (C) incubations expressed as the percentage uptake of labeled substrate per 106 cells. The differences between the mean uptake of glucose and palmitate were statistically significant (p<0.01) in all three cell lines for all three incubations.

The mean uptake of 18F-FDG and 3H-palmitic acid by prostate cells co-incubated for1 hour in the same flask expressed as the percentage uptake of labeled substrate per 106 cells. The three prostate cell lines demonstrated 7-12% palmitate uptake compared to only minimal (<0.4%) uptake for glucose.

Composite time course of uptake of glucose (A) and palmitate (B). The uptake of both substrates was dynamic. There was no saturation of uptake in 2 hours.
Comparison of substrate uptake among three prostate cell lines following 1-hour incubation. For both glucose and palmitate, there were no differences of the mean uptake among all prostate cells. LNCaP and PC3 cells did not demonstrate higher uptake of either glucose or palmitate compared to normal human prostate cells RWPE1.
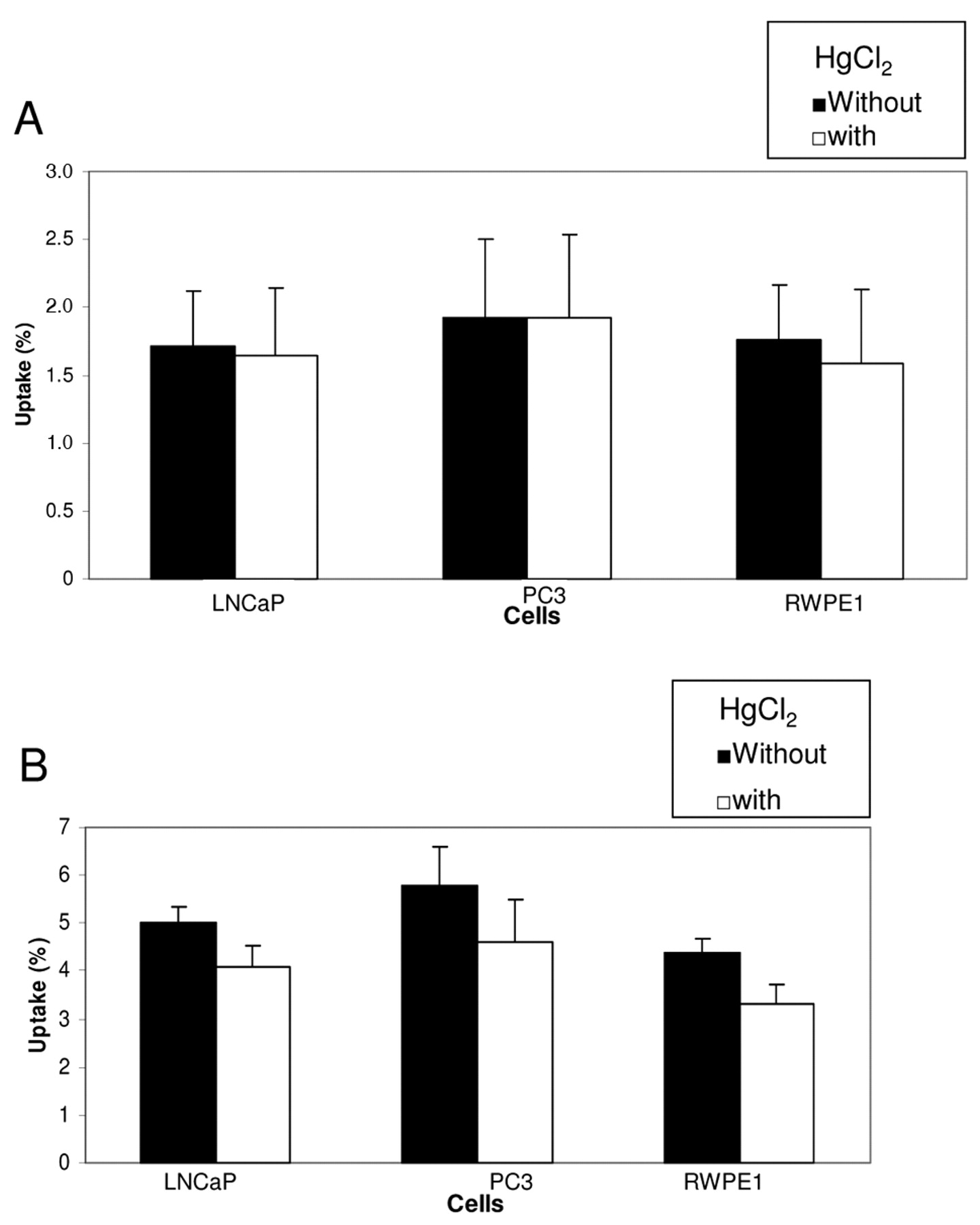
Inhibition of palmitic acid uptake by HgCl2. HgCl2 is a nonspecific protein modifier known to inhibit membrane receptors of the cells (14, 15). In all three cell lines, 2.5 mM HgCl2 inhibited palmitic acid uptake by <10% at 0.5 hour, but the inhibitory effect on uptake at 1 hour increased to between 18% and 25% (p<0.05) (Figure 5). These findings suggest involvement of membrane receptors, at least partially, in the uptake of palmitic acid.
Discussion
It is well established clinically that FDG accumulation is low in prostate tissue and correlates poorly with increasing grade or stage of prostate cancer. Furthermore, there is a significant overlap in FDG uptake between benign prostate hyperplasia and prostate cancer (2, 3). Laboratory studies have revealed that glucose transporter mRNA and protein are only very weakly expressed in human prostate cancer tissue, which is believed to account for its low FDG avidity (16). An in vitro study also suggested that glucose might not be required for androgen-dependent prostate cancer cells because LNCaP cells grow at control rates even in medium containing only 0.05 g/l glucose (17).
Early studies found that human prostatic fluid is rich in citric acid and contains very high levels of total lipid as compared to glucose (18). In addition, the lipid content of the prostate is age dependent, and higher in older patients (19). It has recently been suggested that prostate cancer is associated with fatty acid metabolism, although most of this evidence is from epidemiological studies relating diet, obesity and serum fatty acid levels to the risk of prostate cancer (20-23). In fact, much of the available data is inconsistent and occasionally controversial (24, 25). There is very sparse literature available regarding prostatic metabolism of fatty acid. Freeman et al. (26, 27) recently reported that fatty acid concentrations in non-cancerous prostatic tissue might help to predict risk of biochemical recurrence following radical prostatectomy for clinically localized prostate cancer.

Inhibitory effect of HgCl2 on the uptake of palmitate. HgCl2 had minimal effect on the uptake at 0.5 hour (A), but caused between 18% and 25% of inhibition at 1 hour (B).
It is therefore intriguing to explore whether fatty acid is the dominant bioenergetic substrate within the prostate. It is known that fatty acid catabolism is an important source of energy. For some tissues such as myocardium, fatty acid oxidation is the major pathway of bioenergetic supply. To date, there has been no research report regarding the role of fatty acids as a bioenergetic source in the prostate. Although 14C-acetate is under investigation as a PET tracer for prostate cancer (28-30), it does not reflect fatty acid metabolism and is not a true fatty acid tracer because acetate is converted to membrane-impermeable acetyl-CoA, which enters the tricarboxylic acid (TCA) cycle as an intermediate. Acetyl-CoA can be generated from either glucose or fatty acid oxidation. Our study found there was a significantly higher uptake of fatty acid compared to glucose by the prostatic cells, suggesting that catabolism of fatty acids may be the dominant bioenergetic source in the prostate. There was difference in specific activities of 3H-glucose and 3H-palmitic acid employed in our experiments. Since the same amounts of total radioactivity were used in each flask of different groups, the numbers of molecules of added labeled glucose were 3 times more than those of palmitic acid, which theoretically favors the uptake of glucose. Our results demonstrated several-fold greater uptake of palmitic acid by the prostate cells compared to glucose even when more glucose molecules were added to the medium.
The oxidation of fatty acids produces acetyl-CoA. Acetyl-CoA is the link between the two main metabolic cycles of the mitochondrion – the citric and fatty acid oxidation cycles. This end-product of fatty acid oxidation condenses with oxaloacetate to form citrate which is used in the TCA cycle for efficient ATP production. It is known that increased citrate oxidation is a significant metabolic characteristic for the bioenergy requirement in prostate cancer (31, 32). In contrast to the high citrate levels associated with normal prostate tissue and benign hyperplasia, prostate cancer has a low level of citrate (33). The activity of mitochondrial m-aconitase, an enzyme for the first reaction before citrate oxidation, is significantly higher in prostate cancer compared to normal prostate, which drives the utilization of citrate as an energy source (34). Net citrate production requires the continuous availability of oxaloacetate and acetyl-CoA for continuous citrate synthesis. Acetyl-CoA is the only molecule consumed in the citrate cycle and its continuous availability is crucial for driving citrate oxidation. Because of the limited availability of acetyl-CoA from the low rate of glycolysis, fatty acid oxidation may instead represent its major source. Increased uptake of fatty acid is the first step for subsequent oxidation in prostate cancer cells. Intracellular metabolism is the most important regulator of its initial uptake rate, in other words, the uptake of a substrate by the cells is always proportional to its metabolism (35, 36). Therefore, although we did not directly measure fatty acid oxidation in this preliminary study, it is reasonable for us to assume that fatty acid oxidation in the prostate is the major energetic source.
It is interesting to note that the current study shows no difference in uptake of either palmitic acid or glucose between the two malignant human cell lines (PC3 and LNCaP) and a normal human prostate cell line (EWPE1). In general, malignant cells need more bioenergy for rapid cell proliferation compared to their normal counterparts. Since there is increased citrate oxidation in prostate cancer, one possible explanation for our finding is that normally dominant fatty acid oxidation provides enough acetyl-CoA for accelerated citrate oxidation in prostate cancer.
Increased evidence suggests that cellular uptake of fatty acids is via active transport, mediated by a number of different proteins (14, 37). HgCl2 is a nonspecific, membrane-permeable modifier of sulfhydryl groups on protein and is known to inhibit membrane cell receptors. Our results demonstrated approximately 25% inhibition of palmitic acid uptake by HgCl2 after an one-hour incubation, suggesting that the uptake is, at least in part, mediated by membrane receptors, which is consistent with previous reports (14, 37). Unfortunately, this type of assay does not permit discrimination between transport across the membrane and binding of the fatty acid to the membrane.
Conclusion
Normal and malignant prostate cells are characterized by dominant uptake of fatty acid over glucose. Subsequent intracellular oxidation of these fatty acids may be a major bioenergetic pathway in the prostate. Future development of new diagnostic and therapeutic approaches in prostatic cancer should focus on fatty acid substrate or metabolism. The uptake of fatty acid by the prostate cells is a dynamic, active process mediated by membrane receptors. Because the most significant uptake happens in the first one hour after the incubation of prostatic cells with fatty acid, a routine one-hour interval between the tracer administration and imaging is likely to be sufficient, if a fatty acid tracer is used for PET imaging.
The two malignant prostate cell lines we studied have neither increased fatty acid nor glucose metabolism compared to a benign cell line, consistent with a slow rate of metabolism and growth. Assuming the three cell lines studied were representative of benign and malignant tissue in general, fatty acid imaging would appear to be useful in detection of recurrence and metastasis, but not in differentiating malignant from benign prostate tissue.
Acknowledgements
This work was supported by a pilot grant from the Society of Nuclear Medicine.
Footnotes
- Received September 1, 2009.
- Revision received January 22, 2010.
- Accepted January 25, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- CPT1a regulates the delivery of extracellular fatty acids for cardiolipin turnover in prostate cancer cells
- Adipose triglyceride lipase is regulated by CAMKK2-AMPK signaling and drives advanced prostate cancer
- Imaging Fibroblast Activation Protein Alpha Improves Diagnosis of Metastatic Prostate Cancer with Positron Emission Tomography
- Exploitation of CD133 for the Targeted Imaging of Lethal Prostate Cancer
- Spatial modeling of prostate cancer metabolic gene expression reveals extensive heterogeneity and selective vulnerabilities
- Emergence of MicroRNAs as Key Players in Cancer Cell Metabolism
- Lipid Uptake Is an Androgen-Enhanced Lipid Supply Pathway Associated with Prostate Cancer Disease Progression and Bone Metastasis
- Accelerated lipid catabolism and autophagy are cancer survival mechanisms under inhibited glutaminolysis
- Suppressing fatty acid uptake has therapeutic effects in preclinical models of prostate cancer
- 3D Growth of Cancer Cells Elicits Sensitivity to Kinase Inhibitors but Not Lipid Metabolism Modifiers
- Endothelial cell metabolism in health and disease: impact of hypoxia
- Androgen control of lipid metabolism in prostate cancer: novel insights and future applications
- Conjugated Linoleate Reduces Prostate Cancer Viability Whereas the Effects of Oleate and Stearate Are Cell Line-dependent