


Abstract
We investigated the effects of sodium meta-arsenite (NaAsO2) on human cancer cells (MDA-MB-231, MCF-7 and U-87 MG), dental papilla tissue stem cells (DPSCs) and somatic cells [MRC-5 fetal fibroblasts and adult muscle cells (MCs)] by examining telomeric properties, endogenous reverse transcriptase (RT) activity and the expression of tumorigenesis-linked genes. Half maximal inhibitory concentration (IC50) values were higher in DPSCs and MCs, possessing longer telomere lengths when compared to cancer cells. Levels of telomerase and RT activity, and the expression of protein 53 (p53), B-cell lymphoma 2 (BCL2), nuclear factor kappa-light-chain-enhancer of activated B-cells (NFκB), transforming growth factor beta (TGFβ) and vascular endothelial growth factor (VEGF) were significantly lower in cancer cells following sodium meta-arsenite treatment, whereas the effect was absent or marginally detected in DPSCs and somatic cells. Collectively, sodium meta-arsenite effectively induced cellular cytotoxicity by inhibiting telomerase and RT activity, and down-regulating transcript levels in cancer cells with shorter telomere lengths, whereas more tolerance was evident in DPSCs and somatic cells possessing longer telomere lengths.
Arsenic has been considered as a carcinogen that causes increased incidence of cancer in skin, lung, liver, bladder and kidney in the human population (1-3). An inorganic and trivalent form of arsenic (As3+), such as arsenite, exerts via potential cytotoxic effects through a direct affinity to sulfur groups (4, 5) and considerable accumulation of reactive oxygen species (ROS) that can lead to DNA damage in mammalian cells (6). The induction of chromosomal abnormalities has also been related to its ability to disrupt mitotic progression during cell division (7).
In contrast to its carcinogenic affects, arsenic has also been considered as an effective and successful chemotherapeutic drug for the treatment of acute promyelocytic leukemia (APL) (4, 5). Although the exact mechanism of the anti-tumor effects of arsenic remain poorly understood, proposed actions of arsenic trioxide (As2O3) involve the induction of apoptosis by activating caspases and the generation of ROS (8-10), alteration of growth by disrupting the cell cycle (9, 11), and the down-regulation of angiogenic growth factors (12). In addition, several investigations have also shown that arsenic potently modifies the telomere length (13), reduces the telomerase activity by inhibiting the transcription of the reverse transcriptase subunit of the human telomerase gene (hTERT) (10, 14), and binds telomeric proteins (15). More recently, it has been reported that sodium meta-arsenite, an orally bioavailable trivalent arsenical compound, specifically binds to telomere repeats and induces erosion of telomere repeats and growth inhibition of human prostate cancer cells (16). Moreover, a study involving both arsenic trioxide and sodium meta-arsenite resulted in growth inhibition, apoptosis induction and down-regulation of hTERT expression in leukemia cell lines, but did not demonstrate any major differences between the compounds (17).
In human, telomeres play a critical role in chromosome stability from destruction and fusion by capping of their own DNA strand ends (18). Cells with shorten telomere repeats are induced to cellular senescence by losing the ability of cell division and reach crisis status by continual erosion of telomere repeats (18, 19). However, transformed cells possessing short telomere repeats escape from crisis status due to a high level of telomerase activity which is capable of maintaining or extending the telomeric repeats (20). Most cancer cells with short telomere length normally exhibit unlimited proliferation, resulting from a high level of telomerase activity (19). These distinguishing properties of telomere and telomerase activity displayed in cancer cells may have potential as targets for anticancer treatment.
In addition, endogenous non-telomerase reverse transcriptase (RT) activity has been linked to proliferation, tumor progression and differentiation of cancer cells (21-23). Even though the functions and mechanisms of RT activity in tumorigenesis are still unclear, it has been reported that inhibition of non-telomeric RT activity suppressed the proliferation and tumor progression of cancer cells, and predicted epigenetic regulation of cell differentiation and proliferation (23). These observations propose endogenous RT activity as a viable target in cancer therapy. Moreover, it has been shown that arsenic compounds modulate the activity of some cell cycle proteins and genes, including protein 53 (p53), B-cell lymphoma 2 (BCL2) and nuclear factor kappa-light-chain-enhancer of activated B-cells (NFκB) in cancer cells (8, 9, 24, 25). Furthermore, signaling proteins and growth factors such as transforming growth factor-beta (TGFβ) and vascular endothelial growth factor (VEGF) have also been implicated in promoting the tumor microenvironment (12, 26).
To date, limited information is available on the cytotoxicity of sodium meta-arsenite towards breast cancer and glioblastoma-astrocytoma cell lines, specifically on telomere length, telomerase activity, endogenous RT activity and the expression profile of selected genes linked to tumorigenesis. Moreover, fewer attempts have been made to assess the effects of sodium meta-arsenite exposure on adult stem cells and somatic cells. Therefore, the present study investigated the half maximal inhibitory concentration (IC50) of sodium meta-arsenite in cancer cells, adult stem cell and somatic cell lines, and correlation and comparison between was performed their IC50 values and telomere lengths. We further examined telomerase and RT activity, as well as alterations of selected gene expression patterns in these cells following treatment with sodium meta-arsenite under IC50 values.
Materials and Methods
All chemicals used in the present study were purchased from the Sigma Chemical Company (St. Louis, MO, USA) and media from Gibco (Invitrogen, Burlington, ON, Canada), unless otherwise specified. For all the media, the pH was adjusted to 7.4 and the osmolality to 280 mOsm/kg.
Preparation and culture of cells. MDA-MB-231 and MCF-7 human breast cancer, U-87 MG human glioblastoma-astrocytoma and MRC-5 normal human fetal lung fibroblasts were purchased from the American Type Culture Collection (Manassas, VA, USA). Previously isolated and characterized mesenchymal stem cells derived from dental papilla tissue (DPSCs) and adult muscle cell lines were used for this experiment (27). Briefly, immature third molars were collected from five patients (16-17 years of age, female) and muscle tissues were obtained from four patients (16-17 years of age, female) during orthognathic surgery at Gyeongsang National University Hospital under approved guidelines set by the GNUH IRB-2009-34, after obtaining informed consent from their patients. The finely minced dental papilla and muscle tissues were digested with phosphate buffered saline (PBS) containing 1 mg/ml collagenase type I at 37°C with gentle agitation for 30 min. After centrifugation, the suspension was passed through 100 μm and 50 μm nylon cell strainers to harvest single-cell suspension. The cells were maintained in complete medium which consisted of advanced Dulbecco's modified Eagle's medium (A-DMEM) supplemented with 10% fetal bovine serum (FBS) and 1.0% penicillin-streptomycin (10,000 IU and 10,000 μg/ml, respectively) at 37.5°C in a humidified atmosphere of 5% CO2 in air and then sub-cultured when they had grown to approximately 70-80% confluence. To determine the mesenchymal-specific marker expression, the papilla cells at passage 3 were identified with cell-surface antigen markers by flow cytometry and differentiation capacity, as previously described (27). Stock solution of sodium meta-arsenite (NaAsO2, KOMINOX, provided by KOMIPHARM, Korea) was dissolved in PBS at 50 μM, and stored at −20°C. The stock solutions were freshly prepared daily by diluting with complete medium.
Cytotoxicity test by 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay. MTT assay was used to determine the cytotoxicity of sodium meta-arsenite for each cell line following a previously described protocol (28) with minor modifications. Briefly, 1×104 cells per well were seeded into 96-well plates in complete medium at 37.5°C in a humidified atmosphere of 5% CO2 in air. Following overnight incubation, media were carefully removed, and replaced with complete media containing sodium meta-arsenite at 0, 0.5, 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 μM concentrations, and cells were continuously exposed for 48 h. MTT assay was performed by adding 20 μl of 5 mg/ml MTT to each well, followed by incubation at 37°C. After 3.5 h incubation, the treated wells were covered with tin-foil and agitated on an orbital shaker for 15 min. The resulting purple formazan was measured with an ELISA reader (Bio-Tek, Seatle, WA, USA) at 550-610 nm wavelength and with Gen5 software (Bio-Tek). Growth inhibition was assessed as half maximal inhibitory concentration (IC50) compared with vehicle-treated controls.
Analysis of telomere length by non-radioactive chemiluminescent assay. Telomere length was determined by a non-radioactive chemiluminescent assay with the TeloTAGGG telomere restriction fragment (TRF) length assay kit (Roche, Indianapolis, IN, USA) according to the manufacturer's instructions. Briefly, genomic DNA of the cells was extracted using the DNeasy Blood and Tissue kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. A total of 1 μg genomic DNA was digested with a mixture of HinfI and RsaI restriction enzymes at 37°C. Digested DNA was then separated in 0.8% agarose gel, followed by depurination, denaturation, neutralization, and was then Southern-transferred onto a positively charged nylon membrane (Roche). The membrane was hybridized with a digoxigenin (DIG) labeled telomere probe (Roche) at 42°C for 3 h. The hybridized membrane was washed in a high stringency buffer and incubated with anti-DIG-AP solution. After the final wash, AP substrate was applied and exposed on X-ray film for 20 min at 25°C. The exposed X-ray film was scanned, and the mean length of telomere in the acquired images was calculated using a Gelviewer image processing software (Innogene, Korea).
Analysis of telomerase activity by real-time relative-quantitative telomerase repeat amplification protocol (RQ-TRAP). The real time RQ-TRAP assay was modified from a conventional TRAP assay for its use on the LightCycler 3.0 (Roche) as previously described by Jeon et al. (29). Briefly, cells were either frozen at −80°C for future analysis or lysed in 0.5% (v/v) 3-[(3-cholamidopropyl) dimethylammonio]-1-propanesulfonic acid (CHAPS) lysis buffer (pH 7.5) supplemented with 10 mM Tris-HCl, 1 mM MgCl2, 1 mM EGTA, 0.1 mM benzamidine, 5 mM 2-mercaptoethanol and 10% glycerol at a density of 250 cells/μl for 30 min on ice. Lysed samples were centrifuged at 12,000 ×g at 4°C for 20 min to remove cell debris. The concentration of protein was measured by a spectrophotometer (Mecasys, Korea) and 5 μg of total protein was analyzed by RQ-TRAP assay. Each run included measurements of telomerase-positive control 293T (Chemicon, Lake Placid, NY, USA) samples and telomerase-negative control 293T samples inactivated by incubation for 10 min at 85°C. The RQ-TRAP was optimized using the PCR reagent LightCycler FastStart DNA Master SYBR Green 1 (Roche), according to the manufacturer's instructions, containing 2.5 mM MgCl2, 0.02 μg of primer TS (5’-AAT CCG TCG GAG CAG AGT T-3’), 0.04 μg of primer ACX (5’-GCG CGG CTT ACC CTT ACC CTT ACC CTA ACC-3’). The assay was set up for 30 min incubation at 30°C, followed by 10 min incubation at 94°C, and 40 cycles of PCR at 94°C for 30 s and 60°C for 90 s. All samples were quantified using the LightCycler Quantification Software's (Roche) second derivative method of crossing point (Cp) determination, and relative telomerase activity (RTA) was calculated to ratio based on the level of telomerase activity in the MRC-5 fibroblasts.
Analysis of RT activity. Analysis of RT activity with cell lysates containing the RT enzyme by PCR-based assay was carried out as previously described by Mangiacasale et al. (21) with minor modifications. Briefly, the total protein of cells was extracted with CHAPS lysis buffer as explained above. RT reactions contained 10 μg of total protein, 10 ng of purified bacteriophage MS2 RNA (Roche), 2 μl of reverse transcription buffer (Qiagen), 1 mM of four nucleotide triphosphate mix (Invitrogen), 2 U of RNaseOUT (Invitrogen), and 30 pmol of MS2 reverse primer (see below) in a final volume of 20 μl. Reaction mixtures were incubated at 42°C for 1 h, followed by 5 min at 85°C to inactivate the RNase. For removing the spared MS2 RNA after cDNA synthesis, 2 U of Escherichia coli RNase H (Invitrogen) was added to each sample and the mixture was further incubated at 37°C for 20 min in a thermal cycler (Eppendorf, Hamberg, Germany). Positive control reactions were added with 1 U of Omniscript reverse transcriptase (Qiagen) and negative control reactions were added with diethylpyrocarbonate (DEPC) water. After cDNA synthesis, 4 μl of each reaction was amplified with 10 pmol each of forward (5’-ACC TCC TCT CTG GCT ACC GA-3’) and reverse (5’-ACA GGC AGC CCG ATC TAT TT-3’) primers using PCR pre-mix kit (Intron, Korea), adjusted to a total volume of 20 μl. PCR conditions were set for 10 min incubation at 94°C, and 35 cycles of PCR at 94°C for 30 s and 60°C for 90 s. A total of three RT reactions were used for each RNA sample. The PCR amplification product was a 297-bp DNA fragment spanning positions 2225-2521 at the 5’ end of the MS2 RNA. The PCR products were confirmed through 1.0% agarose gel electrophoresis. The relative quantification of transcripts were calculated with the second derivative method of crossing point (Cp) determination using the LightCycler Quantification Software's (Roche) based on the level of DPSCs.
Quantitative analysis of transcripts by real-time RT-PCR. Total RNA was extracted using the QIAshredder column and RNeasy Micro Kit (Qiagen). Homogenization, isolation, precipitation and purification of RNA were performed according to the manufacturer's procedures with an extra step of DNase I treatment for the removal of DNA contamination. The concentration of total RNA was determined by a spectrophotometer (Mecasys). A total of 1 μg RNA was converted to the first-strand cDNA with Omniscript RT Kit (Qiagen). Each of cDNA reactions contained 2 μl of 10 μM Oligi-dT12-18 primer (Invitrogen), 1 μl of 10 U/μl RNase Inhibitor (Invitrogen), 2 μl RT buffer, 2 μl dNTP, and 1 μl Omniscript (Qiagen), adjusted to a total volume of 20 μl. The cDNA samples were then incubated in a PTC-200 Peltier thermal cycler (MJ research Ramsey, MN, USA) at 42°C for 1 h, followed by 5 min at 95°C to inactivate the enzyme. A total of three reverse transcription reactions were used for each RNA sample. The real-time RT-PCR was carried out using LightCycler 4.0 and the LightCycler Faststart DNA Master SYBR Green I (Roche) according to the manufacturer's protocols. Each reaction mix contained 2 μl of the cDNA reaction, 2 μl of the FastStart DNA Master SYBR Green I reaction mix, 3 mM MgCl2, and 2 μl of each of the forward and reverse primer (0.1 μg/μl), adjusted to a total volume of 20 μl. The amplification protocol consisted of an initial denaturation step for 10 min at 95°C followed by 40 cycles of denaturation for 15 s at 95°C, annealing for 6 s 57-60°C and extension for 16 s at 72°C. Reference genes (glyceraldehyde 3-phosphate dehydrogenase, GAPDH; peptidylprolyl isomerase A, PPIA) and tumorigenesis-related genes (B-cell lymphoma, BCL-2; protein 53, p53; nuclear factor kappa-light-chain-enhancer of activated B-cells, NFκB; transforming growth factor-beta, TGFβ and vascular endothelial growth factor, VEGF) were analyzed for their abundance levels by quantitative real-time RT-PCR. Primer sequences, annealing temperatures and the size of amplified products are shown in Table I. The relative quantification of transcripts were calculated with the second derivative method of Cp determination using the LightCycler Quantification Software's (Roche) based on the level of GAPDH in each cDNA samples.
Statistical analysis. Differences among the treatments were analyzed by using one-way analysis of variance (ANOVA) by SPSS 15.0 (Chicago, IL, USA). Data are expressed as the mean±SE. Comparisons of mean values among the treatments were analyzed using a Turkey's multiple comparisons test. A 5% probability (p<0.05) was used as the level of significance.
Results
Determination of IC50 values in cancer cells, DPSCs and somatic cells. The cytotoxicity of sodium meta-arsenite was determined using IC50 values by MTT assay in MDA-MB-231, U-87 MG, MCF-7, DPSCs, MRC-5 and MCs, and the results are shown in Figure 1A and B. Sodium meta-arsenite treatment dose-dependently inhibited the survival of cancer cells, DPSCs and somatic cells. Cell survival showed that all the tested cancer cell lines respond similarly to sodium meta-arsenite. The mean IC50 value was 2.9±0.35, 3.2±1.32, 3.5±1.17, 29.8±2.01, 5.9±1.93 and 15.5±1.01 μM for MDA-MB-231, U-87 MG, MCF-7, DPSCs, MRC-5 and MCs, respectively. The IC50 values for DPSCs and MCs were significantly (p<0.05) higher when compared to those of MDA-MB-231, U-87 MG, MCF-7 and MRC-5 cells.
Primer sequences, size of PCR products and annealing temperature used for RT-PCR and real time RT-PCR.
Telomere length in cancer cells, DPSCs and somatic cells. Telomere length was analyzed by non-radioactive chemiluminescent assay and the results are presented in Figure 2A and B. The mean telomere length in the spot which shows the highest density was 4.1±0.91, 5.5±1.18, 3.9±1.14, 20.2±3.11, 14.3±2.33 and 15.3±3.23 kbp in the MDA-MB-231, MCF-7, U-87 MG, DPSCs, MRC-5 and MCs, respectively (Figure 2B). The telomere length in the three types of cancer cell lines was significantly (p<0.05) shorter than those of DPSCs and somatic cell lines. Amongst all cell lines, DPSCs exhibited the longest telomere length. We further examined the correlation coefficient analysis between IC50 values and telomere length in each cell line, and the results are shown in Figure 3. A significant correlation coefficient (r2=0.8691) was observed in each cell lines, and the IC50 values increased in proportion to the telomere length of each cell line.
Furthermore, telomere lengths were 3.9±1.28, 5.6±1.01, 3.8±1.91, 19.3±2.93, 7.4±5.09 and 13.6±3.37 kbp in the MDA-MB-231, MCF-7, U-87 MG, DPSCs, MRC-5 and MCs, respectively, when treated with 3 μM sodium meta-arsenite for 48 h. The observed telomere lengths were not significantly (p>0.05) different from those of the untreated contorls, except in the case of sodium meta-arsenite treated MRC-5 cells, which had significantly (p<0.05) shorter telomere length compared to untreated MRC-5 cells (Figure 2B). Moreover, we observed the inhibition of cell growth in MDA-MB-231, and MCF-7 cancer cell lines treated with 3 μM sodium meta-arsenite (Figure 4). Morphological alternations were also evident, with cells appearing as flat or elongated, and eventually retracted and detached from the substratum after treatment with 3 μM sodium meta-arsenite. Based on these observations, the telomerase and RT activity, and gene expression were further investigated in each cell line after treating with 3 μM sodium meta-arsenite.
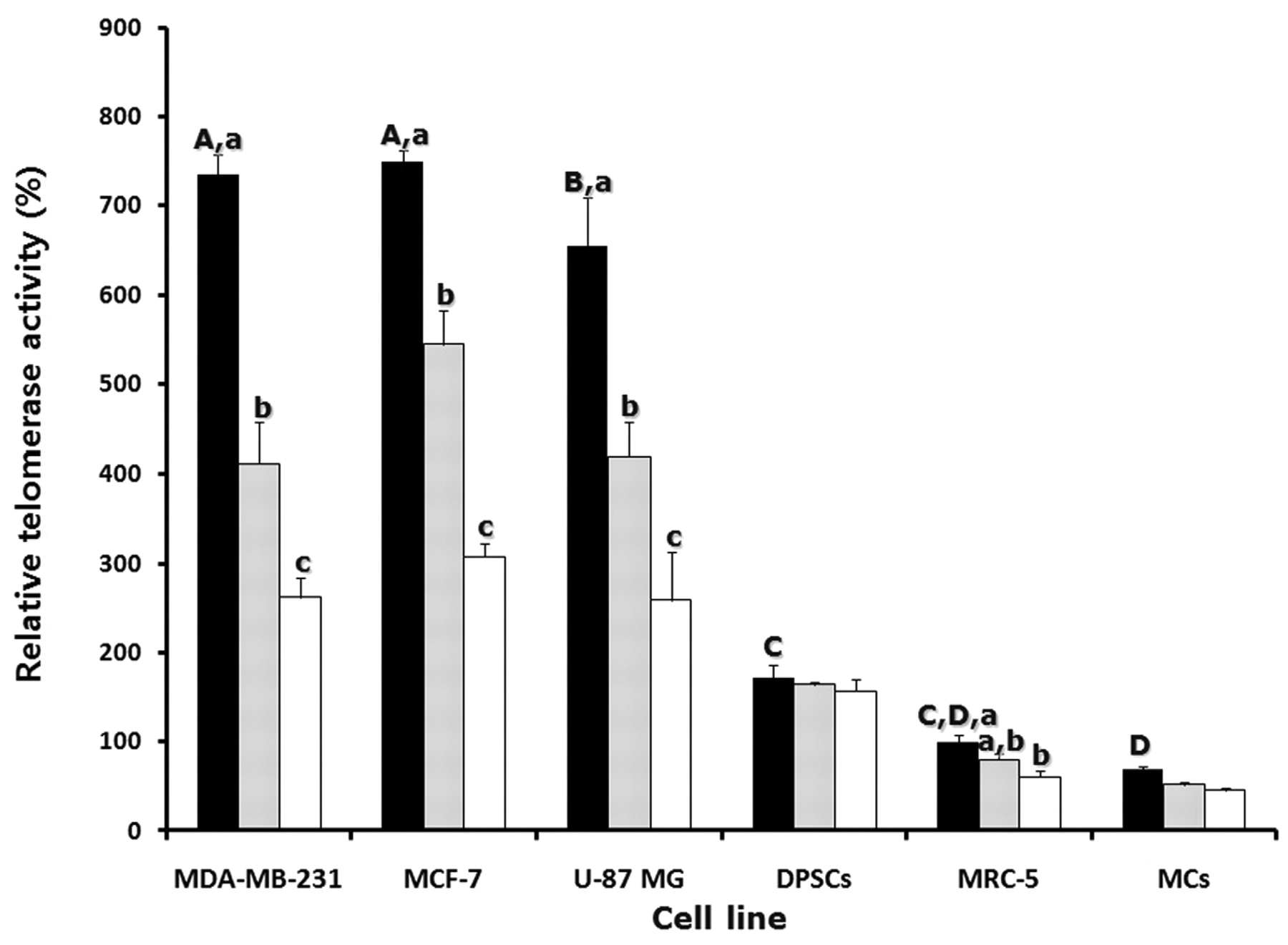
Telomerase activity in cancer cells, DPSCs and somatic cells. Telomerase activity was analyzed by RQ-TRAP assay in MDA-MB-231, MCF-7, U-87 MG, DPSCs, MRC-5 and MCs untreated or treated with 3 μM sodium meta-arsenite for 24 and 48 h, and the results are depicted in Figure 5. The level of telomerase activity in MRC-5 cells was considered as 100% for comparison with other cell lines. The level of telomerase activity was 735±22.2, 749±13.9, 654±53.7, 171±13.7 and 59±2.7% in untreated MDA-MB-231, MCF-7, U-87 MG, DPSCs and MCs, respectively. The level of telomerase activity in the three types of cancer cell lines was significantly (p<0.05) higher than in somatic cell lines, reaching levels over 600%, when compared to MRC-5 cells. However, the level of telomerase activity was significantly (p<0.05) decreased in MDA-MB-231, MCF-7 and U-87 MG cells treated with 3 μM sodium meta-arsenite (411±23.5, 544±14.1 and 419±45.0%, respectively) at 24 h when compared to those of the untreated counterparts. Moreover, the level of telomerase activity was further reduced significantly (p<0.05) to half the levels in treated MDA-MB-231, MCF-7, U-87 MG and MRC-5 cells (261±22.2, 307±13.9, 258±53.7 and 60±7.7%, respectively) at 48 h, in comparison to untreated cells. However, there were no significant (p>0.05) differences in the telomerase activity between treated and untreated DPSCs and MCs.

Analysis of IC50 values by MTT assay. A: Inhibition curves by MTT assay in each cell line as indicated after sodium meta-arsenite (NaAsO2) treatment. Dotted lines, logarithm trend line of inhibition curves. B: Mean±SE (n=3) of IC50 values in the MDA-MB-231, MCF-7, U-87MG, dental papilla stem cells (DPSCs), MRC-5 fetal lung fibroblasts and adult muscle cells (MCs) treated with sodium meta-arsenite. a, b, c and d, significant (p<0.05) differences among cells.
RT activity in cancer cells, DPSCs and somatic cells. To analyze relative RT activity, PCR-based RT assay was performed with MS2 RNA and cell lysates extracted from MDA-MB-231, MCF-7, U-87 MG, DPSCs, MRC-5 and MCs. Relative RT activity in DPSCs was considered as 100% for comparison with other cell lines. Relative RT activity was 128±20.1, 112±12.6 and 129±17.5% in untreated MDA-MB-231, MCF-7 and U-87 MG, respectively (Figure 6A). However, relative RT activity was 36±21.1, 48±12.6, 45±10.5 and 70±5.5% in MDA-MB-231, MCF-7, U-87 MG and DPSCs cells treated with 3 μM sodium meta-arsenite for 48 h, respectively. Sodium meta-arsenite treatment significantly (p<0.05) reduced the RT activity in these cell lines when compared to their untreated counterparts. However, RT activity was not detected in somatic cell lines, MRC-5 and MCs (Figure 6B).
Expression of genes in cancer cells, DPSCs and somatic cells. Quantitative expression of tumorigenesis-linked transcripts (BCL2, NFκB, p53, TGFβ and VEGF) in MDA-MB-231, MCF-7, U-87 MG, DPSCs, MRC-5 and MCs cells treated with 3 μM sodium meta-arsenite treated for 48 h is shown in Figure 7C. GAPDH and PPIA were analyzed as reference genes for the accurate quantification of transcripts among different samples. RT-PCR expressions of GAPDH and PPIA were similar in all the cells examined (Figure 7A). For the real-time RT-PCR analysis, GAPDH was employed as internal control as the levels were not significantly (p>0.05) different between untreated and sodium meta-arsenite treated cells (Figure 7B). Transcript levels of BCL2 were significantly (p<0.05) higher in MDA-MB-231, MCF-7 and U-87 MG cells than in those of others cell lines. However, following sodium meta-arsenite treatment, BCL2 levels significantly (p<0.05) decreased in MDA-MB-231, MCF-7, U-87 MG and MRC-5 cells when compared to those of their untreated counterparts. p53 was diversely expressed in each cell line with significantly (p<0.05) higher levels noticed in DPSCs and MRC-5 cells followed by MCF-7 and MCs, than those of MDA-MB-231 and U-87 MG cell lines. Except for DPSCs and MCs, p53, expression levels were significantly (p<0.05) reduced in sodium meta-arsenite-treated MDA-MB-231, MCF-7, U-87 MG, and MRC-5 cell lines. Levels of NFκB were significantly (p<0.05) higher in MCF-7 cells than other cell lines. But after sodium meta-arsenite treatment, the expression significantly (p<0.05) decreased in MDA-MB-231, MCF-7, U-87 MG, and MRC-5 cells. Transcript levels of TGFβ and VEGF were found to be significantly (p<0.05) higher in MDA-MB-231, MCF-7 and U-87 MG cells when compared to those of other cell lines. As observed with other genes, the levels of TGFβ and VEGF expressions significantly (p<0.05) decreased in sodium meta-arsenite-treated MDA-MB-231, MCF-7, U-87 MG and MRC-5 cells when compared to untreated cells. To a large extent, treatment of 3 μM sodium metaarsenite for 48 h reduced the transcript levels of tumorigenesis-linked genes in the cancer cell lines with low IC50 values more than those of DPSCs and somatic cell lines with high IC50 values.
Discussion
The efficacy of sodium meta-arsenite in anticancer therapy prompted this study to examine its mechanism of action in cancer cell lines (MDA-MB-231, U-87 MG and MCF-7), adult stem cells (DPSCs) and somatic cell lines (MRC-5 and MCs). Our data indicate that the cancer cell lines exhibit lower IC50 values for sodium meta-arsenite in comparison to DPSCs and somatic cells. Differential cytotoxic effects of sodium meta-arsenite were correlated by the fact that cancer cells with shorter telomeres were more sensitive to the compound than those of stem and somatic cells with relatively longer telomeres. Moreover, the treatment of sodium meta-arsenite at a concentration of cancer cell IC50 value significantly reduced the telomerase and RT activity, and down-regulated transcript levels of genes linked to tumorigenesis in the cancer cell lines, without exerting any deleterious effects on DPSCs and somatic cells.
Previous studies have shown the growth inhibitory effects of arsenic compounds on human keratinocytes and leukemia cells, cord blood cells, and prostate cancer cells (8, 13, 16, 30). Supporting these observations, we also found that the treatment of sodium meta-arsenite led to the effective inhibition of the growth phase of cancer cell lines and a dramatic reduction of cell survival with increasing concentration as revealed by MTT assay. Treatment with arsenic compound was shown to increase the G2/M and S phases of the cell cycle, with reduced viability of human HT1197 cells in a dose-dependent manner, implying that the drug was involved in incomplete DNA synthesis (31). In another study, MCF-7 cancer cells treated with sodium meta-arsenite had increased G2/M phase of the cell cycle by arrest of DNA synthesis with an enhanced apoptosis rate (8). Moreover, both arsenic trioxide and sodium meta-arsenite resulted in growth inhibition, apoptosis induction and down-regulation of hTERT expression in leukemia cell lines, without exhibiting any major differences between the compounds (17). These results suggested that arsenite induced cytotoxicity by interfering with the progression of the cell cycle and by inducing DNA fragmentation and apoptosis.
In a recent investigation, it was demonstrated that sodium meta-arsenite led to translocation of the telomerase catalytic subunit into the cytoplasm by binding sodium meta-arsenite to telomeric repeats at a ratio of one molecule per three TTAGGG repeats, and cytotoxicity was linked to rapid telomere erosion through direct telomere binding in prostate cancer cells (16). An earlier report from the same group also showed a potent cytotoxic activity of sodium meta-arsenite by inducing telomere attrition, senescence and chromosomal instability in solid human tumor cell lines with short telomeres (32). Moreover, the IC50 values of sodium meta-arsenite were lower in the cells with shorter telomeres than in those with longer telomere length (16), indicating a strong association between telomeres and cytotoxicity of sodium meta-arsenite. Our study also provides the evidence that the sodium metaarsenite cytotoxicity correlats with the shorter telomere repeats in human breast cancer and glioblastoma-astrocytoma cell lines. Furthermore, DPSCs and MCs with longer telomere lengths had higher IC50 values relative to the cancer cell lines used here. Interestingly, even though telomere length in the MRC-5 fetal lung fibroblasts was greater than those of the cancer cell lines, its IC50 value was low in our results. Here, there may be a possibility that the difference between the susceptibility to sodium meta-arsenite of two somatic cell lines resulted from the cell origin or the alterations occurring during routine culture and maintenance. Furthermore, unlike in an earlier report (16), conclusive evidence on the exclusion of ROS as a causative factor of telomere erosion was not ascertained in the present study. However, all this evidence indicates that cancer cells with shorter telomeres are more susceptible to sodium meta-arsenite than cells with longer telomeres.

Analysis of telomere length by non-radioactive chemiluminescent assay. A: Representative image showing telomere length in the control (lane 1-6) and sodium meta-arsenite-treated cell lines at IC50 values for 48 h (lane 7-12). Lane 1 and 7, U-87 MG; lane 2 and 8, MCF-7; lane 3 and 9, MDA-MB-231; lane 4 and 10, MRC-5; lane 5 and 11, DPSCs; lane 6 and 12, MCs. B: Mean±SE (n=3) telomere length in the control (black bars) and cell lines treated with 3 μM sodium meta-arsenite for 48 h (white bars). a, b, c and d, significant (p<0.05) differences among cell lines. *, Significant (p<0.05) difference between the control and sodium meta-arsenite-treated cells.
Although most cancer cells exhibit shorter telomere lengths, the telomerase activity is up-regulated or re-activated to maintain the telomere length at immortal status without undergoing cellular senescence or crisis, thus overcoming the limited capacity for proliferation (20). We also observed higher levels of telomerase activity in the cancer cell lines with unlimited proliferative capacity, compared to those of DPSCs and somatic cell lines. It is well known that telomerase activity is mainly regulated by the expression of telomerase reverse transcriptase (TERT) catalytic subunit, RNA-dependent polymerases, and telomerase RNA component (TERC) (18) and among these, the expression of TERT is considered as a critical factor. Most cancer cells exhibit the up-regulated telomerase activity by overexpression of TERT through demethylation of its promoter and increasing copy number of TERT loci (33). Several reports have shown that the treatment with arsenic compounds at higher concentrations induces the down-regulation of telomerase activity by inhibiting TERT mRNA and/or protein in human cancer cell lines (10, 14, 16, 34). Our results clearly demonstrated the down-regulation of telomerase activity in sodium meta-arsenite treated cancer cell lines, and we suppose that the reduced proliferative capacity of cancer cells may be related to down-regulation of telomerase activity. Interestingly, except for a noticeable decrease in the telomerase activity of the MRC-5 fibroblast cell line, treatment with sodium meta-arsenite did not exert any inhibitory effects on DPSCs and MCs. Even though low concentrations of arsenic compounds induce telomerase expression, and maintain telomere lengths and cellular growth in normal and cancer cells (13, 30), exposure to increased concentrations inhibits telomerase activity and enhances apoptosis rates as a result of an increase in genomic instability and chromosomal abnormalities (13, 14, 16, 34). Therefore, down-regulation of telomerase activity by sodium meta-arsenite may be considered as a critical event that decides the survival and proliferation of cancer cells. Our results further emphasize the evaluation of telomerase activity as a reliable biomarker for assessing the antitumor activity of sodium meta-arsenite.
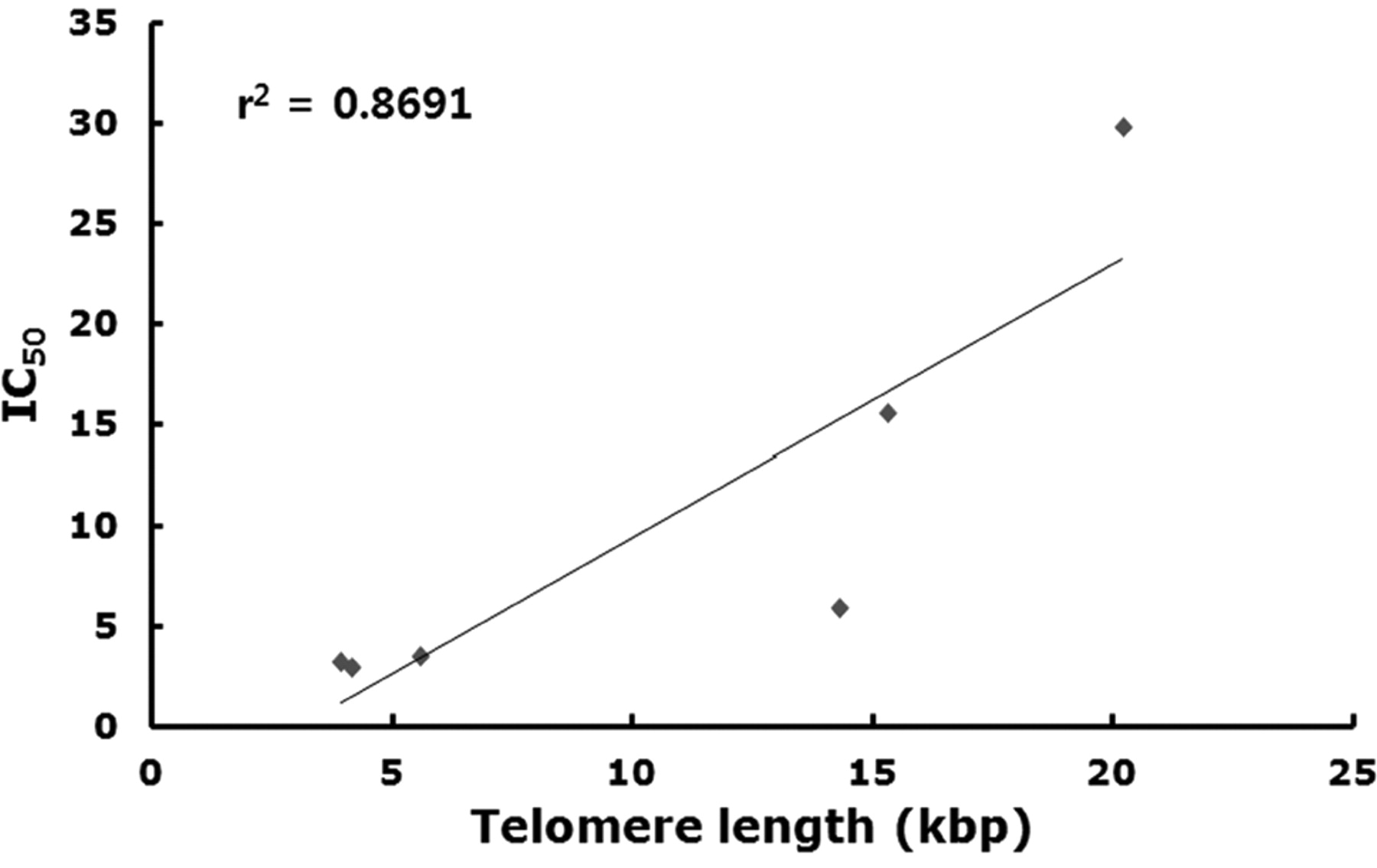
A. Correlation coefficient analysis of IC50 values and telomere length in each cell line. Increased susceptibility of cells to sodium meta-arsenite corresponded with lower telomere length.
Earlier studies provide the insight that the up-regulation of telomerase activity can be influenced by an increased copy number of TERT to more than two copies per cell on the chromosome of most cancer and immortalized cell lines (33, 35, 36). The exact causes for the increased copy number of TERT are still unclear. However, we speculate that the role of endogenous RT activity in increasing the copy number of TERT in the cancer cell lines by incorporation of TERT, intermediates into different locations of DNA. This is partly supported by a fact showing the up-regulation of endogenous non-telomeric RT activity by encoded retrotransposons in the early pre-implantation mouse embryos and cancer cells, but its inhibition by different methods reduced progression and proliferation, and increased cellular differentiation in a variety of cancer cell lines (21, 22, 23, 37). Moreover, morphological changes and differentiation by altered cellular shape and increased adhesion were evident in cancer cells with pharmacological inhibition of RT activity (23). In support of these observations, treatment with 3 μM sodium meta-arsenite in our study also induced morphological alterations along with reduced RT activity in the cancer cell lines. Even though the biological characterization and exact mechanisms of endogenous RT activity are not fully understood, based on our findings, we suppose that RT activity is influenced by genetic or epigenetic modifications and may have an important role in cellular proliferation and differentiation in cancer cells.

Effect of sodium meta-arsenite on cell morphology and growth. Appearance of MDA-MB-231 cells at 12 h of seeding (A), after 48 h of seeding (B) and after treatment with sodium meta-arsenite for 48 h (C) under phase contrast microscopy. Nuclei of MCF-7 cells (D and E) and DPSCs (F and G) stained with 4’,6-diamidino-2-phenylindole (DAPI) under fluorescence microscopy. MCF-7 and DPSCs after 48 h of seeding (D and F, respectively). Morphological alterations of cells, such as flattening or elongation with detachment from substratum, were observed in MDA-MB-231 and MCF-7 cells treated with 3 μM sodium meta-arsenite (C and E, respectively). In contrast, DPSCs retained their normal shape with no noticeable detachment after sodium meta-arsenite treatment (G). Scale bar=100 μm.
The expression profile of selected genes related to cancer and cell growth was analyzed in the cancer cells, DPSCs, and somatic cell lines. BCL2 gene generally regulates the release of apoptotic factors from mitochondria during apoptosis and its overexpression is involved in resistance to conventional cancer treatment (38, 39). Supporting this, up-regulation of BCL2 gene has been reported in the chronic lymphocytic leukemia (40). We also found higher levels of BCL2 gene in the cancer cell lines, but sodium meta-arsenite treatment down-regulated its expression in these cell lines as observed in earlier investigation using arsenic compound (24). However, no changes were noticed in DPSCs and MCs. In contrast to these observations, BCL2 levels were increased following sodium meta-arsenite treatment in the human malignant melanoma A375 cell line (9). The p53 tumor suppressor gene is inactivated or down-regulated in most human cancer cells, and is functionally involved in the regulation of the cell cycle and apoptosis in response to diverse cellular stresses (41). A previous report has demonstrated the increased levels of p53 protein in A375 human melanoma cells after sodium meta-arsenite treatment in a concentration- and exposure time-dependent manner (9). In contrast, a reduced expression of p53 protein was observed in response to different exposure time or the dose of arsenite drug in human keratinocytes (25, 42). In our study, a lower expression of p53 was observed in the cancer cells compared to DPSCs and somatic cells under control conditions. However, sodium meta-arsenite treatment down-regulated the p53 expression in the cancer cell lines and MRC-5 fibroblasts without affecting its abundance in DPSCs and MCs. Interestingly, the down-regulation of both BCL2 and p53 expression in the cancer cell lines could be associated with the reduced cell survival after sodium meta-arsenite treatment. Furthermore, these results emphasize the presence and correct functioning of BCL2 and p53 in DPSCs and MCs even after sodium meta-arsenite exposure. The findings collectively demonstrated that the arsenic-induced effects on the expressions of BCL2 and p53 both at mRNA and protein levels are associated with divergent results, and these differential responses could be related to the cell specificity.

Analysis of telomerase activity by real-time telomeric repeats amplification protocol (RQ-TRAP) with LightCycler in the control (black bars) and cell lines treated with 3 μM sodium meta-arsenite for 24 h (grey bars) and 48 h (white bars). The relative telomerase activity in MRC-5 was considered as 100% for comparison with other cell lines. A, B, C and D, Significant (p<0.05) differences among cell lines. a, b and c, Significant (p<0.05) difference between the control and sodium meta-arsenite-treated cell line. Values are mean±SEM (n=3).
NFκB regulates the transcription of numerous genes that are linked to tumorigenesis and the expression of these genes may be suppressed through inhibition of NFκB activation (25). Furthermore, it has been reported that NFκB regulates the expression of p53 as well as VEGF, a growth factor necessary for the growth of new blood vessels, and also an abundant cytokine in the tumor microenvironment (25, 43). Moreover, different activation pathways of NFκB may cause the expression of proteins that promote or inhibit apoptosis (43). In addition, TGFβ is a protein that controls proliferation and cellular differentiation in normal cells, but has been reported to be increased for the tumorigenicity of glioma-initiating cells by autocrine signaling pathway (26). We recorded the transcripts of NFκB, TGFβ and VEGF to be of higher levels in the cancer cell lines, but their expressions were down-regulated in the sodium meta-arsenite-treated cancer cell lines and MRC-5 cells. However, the levels were not altered in DPSCs and MCs. We have considered that the down-regulated transcript level of genes may be due to cellular apoptosis by cytotoxicity of sodium meta-arsenite in the cancer cell lines and MRC-5 (low IC50 values), but not in DPSCs and MCs (high IC50 values). Furthermore, there are possibilities that the activation of key cytotoxic factors in sodium meta-arsenite-treated cancer cells reduce the expression of growth factors and cytokines, resulting in the generation of more cellular mediators and the formation of a pro-apoptotic microenvironment.
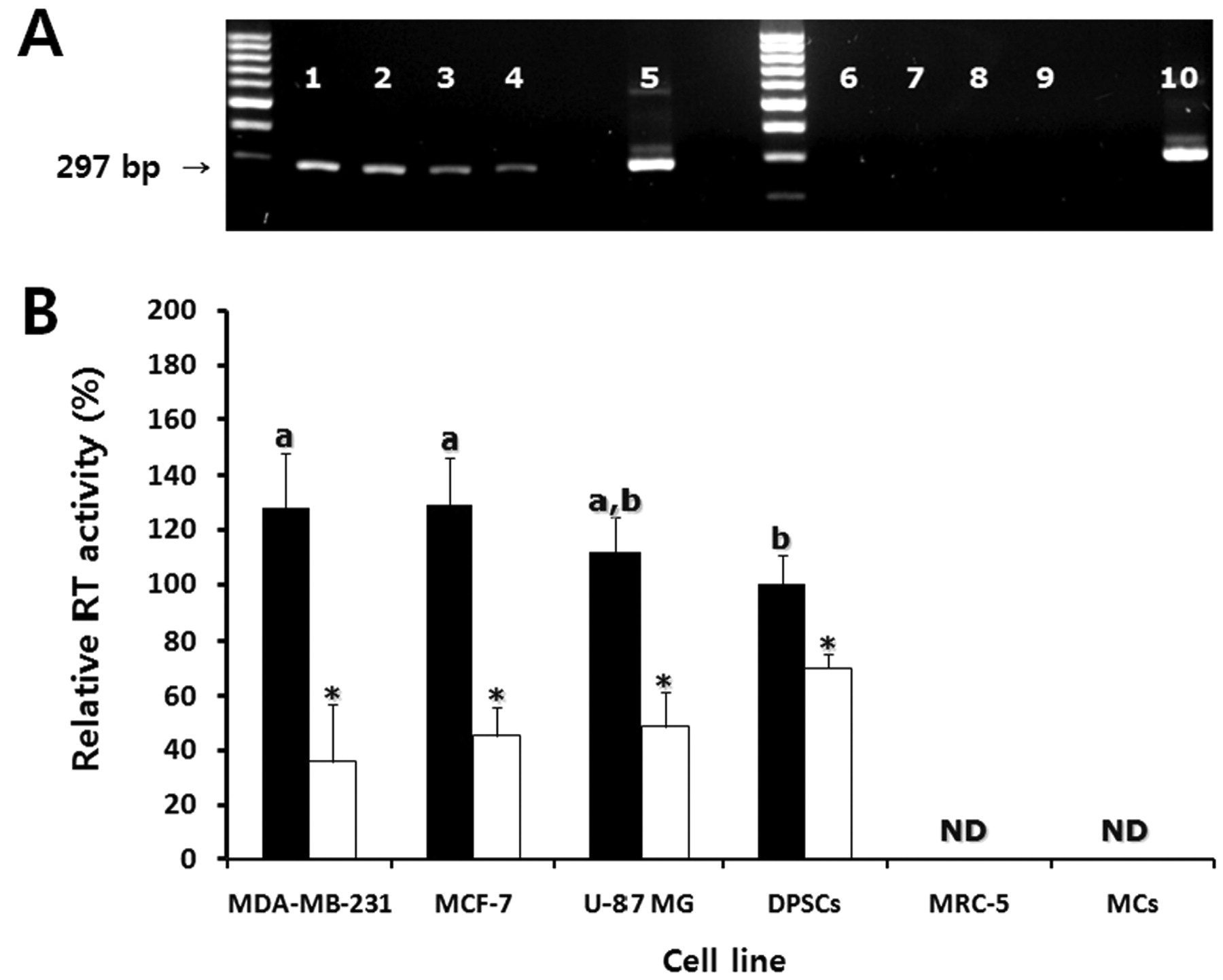
Analysis of reverse transcriptase (RT) activity by PCR-based assay. A: Representative gel image of RT activity by using MS2 RNA and cell lysate in the control cell lines. Lane 1, MCF-7; lane 2, MDA-MB-231; lane 3, U-87 MG; lane 4, DPSCs; lane 5 and lane 10, positive control using commercial RT enzyme; lane 6, MRC-5; lane 7, MCs; lane 8, negative control reacted with water instead of cell lysates; lane 9, negative control reacted with water instead of MS2 RNA. B: Mean±SEM (n=3) of relative RT activity by quantitative real-time RT-PCR using LightCycler in the control (black bars) and cell lines treated with 3 μM sodium meta-arsenite for 48 h as indicated (white bars). The RT activity in DPSCs was considered as 100% for comparison with other cell lines. a and b, Significant (p<0.05) differences among cell lines. *, Significant (p<0.05) difference between the control and sodium meta-arsenite-treated cell line. ND: Not detected.
In conclusion, sodium meta-arsenite may be a potentially useful chemotherapeutic drug for human breast cancer and glioblastoma-astrocytoma cells by exerting its effects on the telomeric properties and gene expression profile. Our observations clearly show that these cancer cells, with shorter telomere lengths, have the low IC50 values rendering them more sensitive to sodium meta-arsenite treatment when compared to DPSCs and somatic cells. Furthermore, 48 h exposure to sodium meta-arsenite reduced the telomerase and RT activity, and down-regulated the transcript level of genes associated with tumorigenesis in the cancer cell lines without causing any repressive effects on DPSCs and MCs. However, further studies are warranted to investigate the effects after prolonged exposure of sodium metaarsenite on the different types of cancer cell lines, including somatic cells with shorter telomere lengths.

Gene expression analysis by RT-PCR and real time RT-PCR. A: Expression of two reference genes (GAPDH and PPIA) in the cell lines as indicated. B: Expression of GAPDH by real-time RT-PCR using LightCycler in the control (black bars) and cell lines treated with 3 μM sodium meta-arsenite (white bars). C: Mean±SE (n=3) of relative transcript levels by real-time RT-PCR in the control (black bars) and cell lines treated with 3 μM sodium meta-arsenite for 48 h (white bars). The transcript levels in MDA-MB-231 cells were considered as 100% for comparison with other cell lines. a, b, c, d and e, significant (p<0.05) differences among cell lines. *, Significant (p<0.05) difference between the control and sodium meta-arsenite-treated cell lines.
Acknowledgements
This study was supported by grants from BioGreen 21, 20070301034041 and 200908FHT010204005, Rural Development Administration, Republic of Korea.
Footnotes
-
Declaration of Interest
GJ Rho was partially supported by KOMIPHARM, South Korea. The authors alone are responsible for the content and writing of this article.
- Received September 29, 2011.
- Revision received November 2, 2011.
- Accepted November 3, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}