


Abstract
Aim: To enhance the anticancer activity of vinorelbine, cisplatin and ionizing radiation (IR) combination against non-small cell lung cancer (NSCLC) cells by co-administration of sodium valproate (VPA), a histone deacetylase inhibitor, and to elucidate molecular events underpinning treatment efficacy. Materials and Methods: The NSCLC A549 cell line was treated with cisplatin (0.2 μg/ml), vinorelbine (2 nM), VPA (1 mM) and IR (2.5 Gy) alone, or in combination. Cell proliferation, cell-cycle distribution, apoptosis, and levels of DNA double-strand breaks, activated DNA damage checkpoint kinases pCHK1, pCHK2, cell-cycle inhibitors p21CIP1/WAF1 and p27KIP1 were assessed. Results: VPA markedly enhanced the DNA-damaging effect of the cisplatin-vinorelbine-IR combination and induced increased DSBs, and expression of pCHK2, pCHK1, p21CIP1/WAF1 and p27KIP1. These molecular changes led to cell-cycle arrest and increased apoptosis and consequently markedly curtailed cancer cell growth. Conclusion: VPA markedly enhances the anticancer activity of cisplatin-vinorelbine-IR combination. This finding has translational implications for enhancing the efficacy of anticancer treatment and for reducing side-effects by reducing doses of radiation and drugs.
Non-small cell lung cancer (NSCLC) accounts for 80%-85% of all lung cancer cases (1). A considerable number (up to 30%) of patients with NSCLC present with locally advanced, unresectable stage III disease (2). For these patients, concomitant chemoradiotherapy was shown to be the best strategy (3). Vinorelbine, a vinca alkaloid, in association with a platinum compounds such as cisplatin or carboplatin, together with thoracic radiotherapy is considered a relatively safe treatment regimen (4). However, treatment outcomes of NSCLC with concurrent chemoradiotherapy remain poor. Therefore, the search for new anticancer drug combinations for concurrent chemoradiotherapy of inoperable locally advanced NSCLC is mandatory and remains a key issue of public health concern.
Pre-clinical research has demonstrated that histone deacetylase (HDAC) inhibitors exhibit anticancer activity against NSCLC cell lines when used alone or in combination with conventional chemotherapeutic drugs or radiotherapy (5-7). Sodium valproate (VPA), a widely used anti-epileptic drug, also exhibits HDAC inhibitory activity and like other HDAC inhibitors exerts antitumor effects (8). The ability of VPA to sensitize cancer cells to the damaging effect of ionizing radiation (IR) and of other anti-neoplastic agents (9, 10) appears to be the most promising anticancer feature of this drug.
The main aim of the present pre-clinical study was to assess whether concomitant VPA administration enhances the anticancer activity of the conventional cisplatin-vinorelbine-IR treatment. The study was performed using the human NSCLC cell line A549. To appraise the efficacy of the suggested combination, we determined (i) cell proliferation, cell-cycle distribution profile and apoptosis; (ii) the DNA damage response (DDR) by measuring levels of DNA double-strand breaks (DSBs) and of activated CHK1 and CHK2 checkpoint kinases; and (iii) assessed the expression levels of the cell-cycle regulatory proteins cyclin-dependent kinase inhibitors p21CIP1/WAF1 and p27KIP1.
Materials and Methods
Materials. VPA, cisplatin, vinorelbine and 4-methylumbelliferyl phosphate (4-MUP), a fluorogenic substrate for alkaline phosphatase (AP), were purchased from Sigma (St. Louis, MO, USA). Rabbit monoclonal antibody raised against phospho-H2A.X (Ser139), γ-H2A.X, a marker of DSBs, and rabbit polyclonal antibodies against phosphorylated (i.e. activated) pCHK1 (Ser345) and pCHK2 (Thr68) were obtained from Cell Signaling Technology, Inc., Danvers, MA, USA. The antibodies against cell-cycle regulatory proteins p21CIP1/WAF1 and p27KIP1 were purchased from Lab Vision Corporation, Fremont, CA, USA. Mouse monoclonal antibody to glyceraldehyde 3-phosphate dehydrogenase (GAPDH), as a housekeeping protein, was obtained from EMD Millipore Corporation, Billerica, MA, USA. Rabbit monoclonal antibody to GAPDH was supplied by Cell Signaling Technology, Inc., Danvers, MA, USA. The secondary antibodies: anti-mouse and anti-rabbit Universal Immuno-Peroxidase Polymer, and anti-mouse and anti-rabbit Universal Immuno-Alkaline-Phosphatase Polymer were provided by Nichirei Biosciences, Inc., Tokyo, Japan. Amplex Ultrared Reagent, a fluorogenic substrate for horseradish-peroxidase (HRP), was purchased from Molecular Probes, Inc., Eugene, OR, USA.
Cell culture. The human NSCLC cell line A549 (CCL-185; American Type Culture Collection, Manassas, VA, USA) was cultured at 37°C and 5% CO2 in humidified air in RPMI-1640 medium supplemented with 10% FCS, 2 mM sodium pyruvate, 2 mM L-glutamine, penicillin (100 IU/ml) and streptomycin (100 μg/ml).
Cell proliferation assay. Five hundred A549 cells/well were seeded simultaneously into two 96-well tissue culture plates. One plate was used for the IR experiments and the other served as the non-irradiated control. Cells were grown overnight and pre-treated for 48-h with cisplatin (0.2 μg/ml), vinorelbine (2 nM) and VPA (1 mM) alone, and in different combinations. Next, cells were irradiated at 2.5 Gy using Clinac-18 linear accelerator (Varian Associates, Inc., Palo Alto, CA, USA) and incubated for an additional 72 h. Control cells were mock-treated. Cell proliferation was measured using the crystal violet assay (11). Briefly, cells were fixed, stained with crystal violet, the incorporated dye was solubilized and the absorbance was then measured at 550 nm using a microplate reader (VERSAmax; Molecular Devices Inc., Sunnyvale, CA, USA).
Cell-cycle distribution analysis and apoptosis measurements. Thirty-seven thousand A549 cells were seeded into 25-cm2 flasks. Cells were treated as described above for the cell proliferation assay. Cell-cycle distribution was determined by flow cytometric analysis of DNA content using propidium iodide. Apoptosis was assessed by measurement of the typical pre-G1 (<2N ploidy) fraction. These experiments were performed using Cytomix™ FC500 flow cytometer with CXP software (Beckman-Coulter, Brea, CA, USA).
Determination of specific protein expressions by cell-based ELISA. γ-H2A.X, pCHK1, pCHK2, p21CIP1/WAF1 and p27KIP1 were assessed by fluorometric cell-based ELISA (12) modified in our laboratory. Three thousand A549 cells/well were seeded into two 96-well fluorescence tissue culture plates and treated as outlined under the cell proliferation assay. Twenty-four hours after irradiation, cells were fixed, permeabilized and exposed simultaneously to two primary antibodies raised in different species (mouse and rabbit). One of these primary antibodies was against one of the specific proteins of interest and another primary antibody was against the housekeeping protein GAPDH. Cells were then exposed to a mixture of anti-mouse and anti-rabbit secondary antibodies. The secondary antibodies against primary antibodies to specific proteins were conjugated to HRP. The secondary antibodies against primary antibodies to GAPDH were conjugated to AP. Immunostaining was detected using two spectrally distinct fluorogenic substrates. For HRP-conjugate detection, Amplex Ultrared Reagent was added and for AP conjugate detection, 4-MUP was added. Fluorescence measurements were made in a 96-well fluorometer (SpectraFluor Plus, Tecan, Durham, NC, USA) using excitation/emission wavelengths of 535/595 nm for HRP and excitation/emission wavelengths of 360/465 nm for AP. Values for specific proteins were normalized to the quantity of housekeeping protein GAPDH by dividing the fluorescence of each specific protein by the fluorescence of GAPDH in the same well. Results are expressed in arbitrary units.
Statistical analysis. Unpaired t-test was used. Data are presented as the mean±SEM. A value of p<0.05 was considered statistically significant.
Results
Inhibition of cell proliferation. A549 cells were treated with low doses for cisplatin (0.2 μg/ml), vinorelbine (2 nM), VPA (1 mM) and IR (2.5 Gy). Single-drug treatments reduced cell growth by 7.5%-20% (Figure 1). The cisplatin-VPA combination was the most effective two-drug treatment, reducing A549 proliferation by 38.8%. However, the triple combination of cisplatin-vinorelbine-VPA was more effective and curtailed A549 cell growth by 46.4%. IR treatment-alone decreased cell growth by 17.5%. The combination of IR with VPA was more effective than the combination of IR with cisplatin (p<0.0001) or vinorelbine (p<0.004) in restraining cell growth. The combination of cisplatin-VPA followed by IR suppressed A549 cell growth more markedly than the combinations of cisplatin-vinorelbine and VPA-vinorelbine followed by IR (p<0.0001). Co-application of IR with all three drugs was the most effective treatment, impeding cell growth by 48.8% compared to the control (p<0.0001) and by 4.6%, compared to three-drug treatment alone (p<0.02). Finally, co-application of VPA with the conventional combination of cisplatin-vinorelbine-IR resulted in an additional 21.5%, inhibition of cell growth (p<0.0001).
Cell-cycle analysis and apoptosis measurements. Cell-cycle analysis showed a significant decrease of the G2-phase peak in cells treated with two-drug combinations (Figure 2). However, the three-drug combination was more effective and reduced the G1-phase peak significantly compared to two-drug treatments (p<0.03). Co-application of IR with the two-drug treatments resulted in an additional decrease of the G1-phase peak (p<0.05) compared to the same treatments without IR. Co-application of IR with the three-drug combination was the most effective and reduced the G1-phase peak by 27.7% compared to the control (p<0.0003) and by 6.8% compared to the three-drug treatment alone (p<0.003).
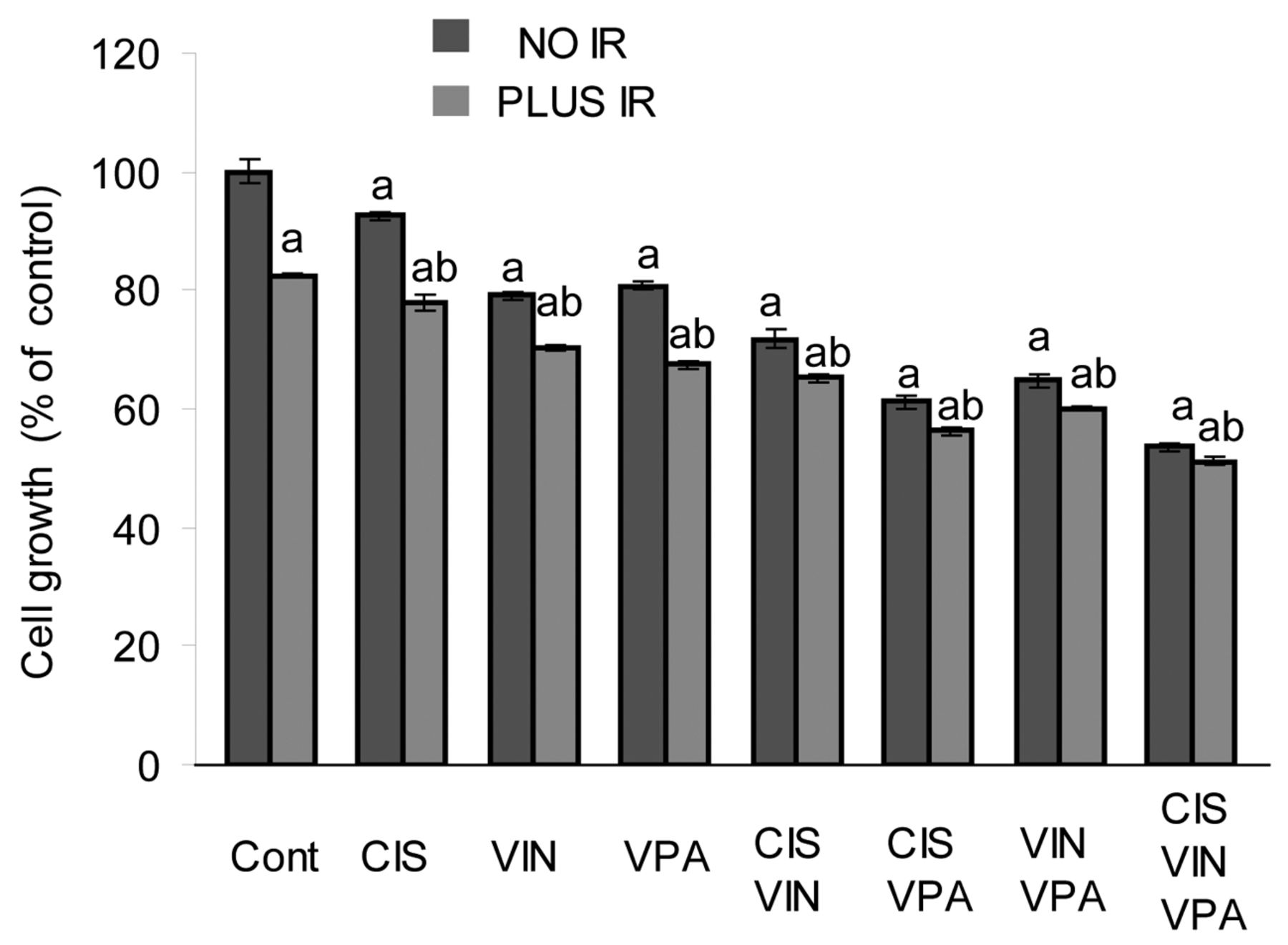
Effect of cisplatin (CIS), vinorelbine (VIN), sodium valproate (VPA) and ionizing radiation (IR) alone and in combined treatments on A549 cell growth. ap<0.005 compared to untreated cells (Cont), bp<0.02 compared to analogous treatment without IR.

Effect of cisplatin (CIS), vinorelbine (VIN), sodium valproate (VPA) and ionizing radiation (IR) alone and in combined treatments on cell-cycle distribution in A549 cells. A: Without IR, B: with IR. *p<0.05, **p<0.01, ***p<0.001 compared to untreated cells (Cont).

Effect of cisplatin (CIS), vinorelbine (VIN), sodium valproate (VPA) and ionizing radiation (IR) alone and in combined treatments on formation of DNA double-strand breaks in A549 cells. ap<0.04 compared to untreated cells (Cont), bp<0.05 compared to all other treatments without IR, cp<0.03 compared to cells treated with IR only, dp<0.0001 compared to all other treatments except the combination of CIS, VPA and IR, ep<0.03 compared to analogous treatment without IR.
It is worth noticing that the different combinations of cisplatin, vinorelbine and VPA with and without IR increased the percentage of S-phase cells. However, this effect reached the significance level only in cells treated with the combinations of cisplatin-vinorelbine-VPA, cisplatin-VPA-IR and cisplatin-vinorelbine-VPA-IR (Figure 2). No specific pattern of changes in the G2-M phase peak was observed despite statistically significant changes in some treatments. The most prominent finding pertaining to the cell-cycle experimental series was the increased expression of the pre-G1 peak, a well-established index of apoptosis. The increase of the pre-G1 peak became highly significant in cells treated with the combinations of drugs. The triple-drug treatment increased the percentage of apoptotic cells from 0.8% to 6.4% and was markedly more effective than two-drug combinations (p<0.0003). Co-application of IR with other treatments resulted in synergistically enhanced apoptosis (p<0.02) compared with treatments without IR. The highest apoptosis was found in cells treated with a combination of IR and all three drugs: the pre-G1 peak was 12-fold higher than in control cells.
DNA-damaging effects. Among the single-drug treatments only VPA significantly raised the level of γ-H2A.X, a marker of DSBs (13) (p<0.0001, Figure 3). Surprisingly, treatment with vinorelbine reduced the level of DSBs (p<0.03). The ability of vinorelbine to reduce the level of DSBs was also observed when it was co-applied with cisplatin, or VPA, or IR. The combination of VPA and cisplatin induced DSBs formation more markedly than VPA applied alone (p<0.004). Of note, the cisplatin-vinorelbine-VPA combination was more effective than the cisplatin-VPA combination (p<0.05). IR alone significantly raised the level of DSBs in A549 cells (p<0.03). The combination of IR with VPA increased the level of DSBs to a greater extent than the combination of IR with cisplatin (p<0.001). However, co-application of IR with cisplatin-VPA or with cisplatin-vinorelbine-VPA combination was more effective and raised the level of DSBs by 79.8% (p<0.0001) and by 91.6% (p<0.0001), respectively, compared to non-treated cells.
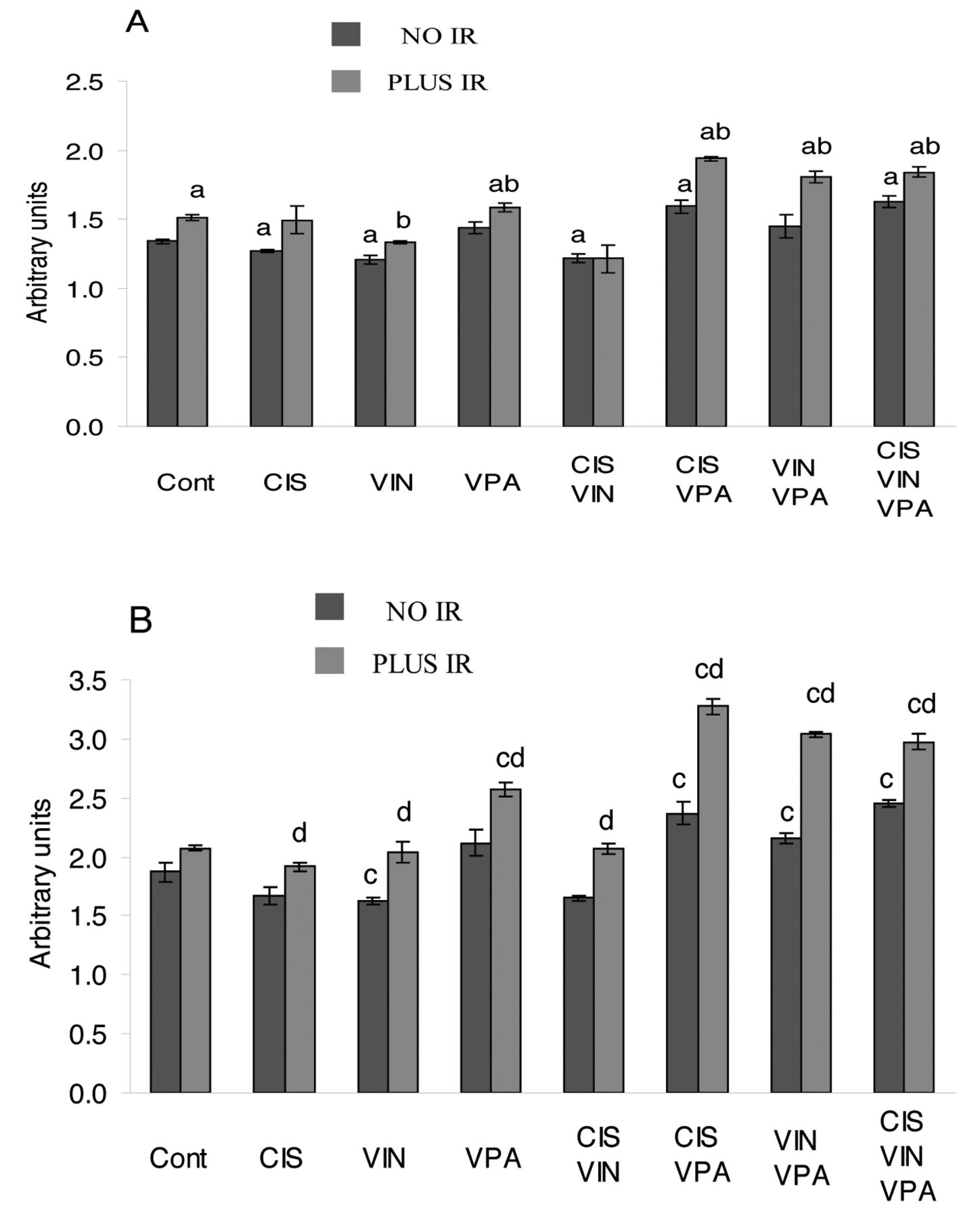
As predicted, changes in activated CHK2 and CHK1 kinases (Figure 4), components of ATM/CHK2 and ATR/CHK1 pathways were similar to those found in γ-H2A.X. Of note, the expression changes in pCHK2 were of a higher magnitude than those found in pCHK1. Similar to findings pertaining to γ-H2A.X, the combinations of cisplatin-VPA and cisplatin-vinorelbine-VPA substantially activated CHK2 and CHK1 kinases. Of note, maximal CHK2 and CHK1 activation was achieved in irradiated cells pre-treated with the combination of cisplatin-VPA (pCHK2, p<0.04 compared to all treatments; pCHK1, p<0.03 compared to all other treatments, except for the combination of cisplatin-vinorelbine-VPA-IR). The combination of cisplatin-VPA-IR increased levels of activated CHK2 and CHK1 kinases by 75% and by 45%, respectively,

Effect of cisplatin (CIS), vinorelbine (VIN), sodium valproate (VPA) and ionizing radiation (IR) alone and in combined treatments on levels of activated pCHK1 and pCHK2 in A549 cells. A: CHK1, B: CHK2. ap<0.05 compared to untreated cells (Cont) for CHK1, bp<0.04 compared to analogous treatment without IR for CHK1. cp<0.05 compared to untreated cells (Cont) for CHK2, dp<0.04 compared to analogous treatment without IR for CHK2.
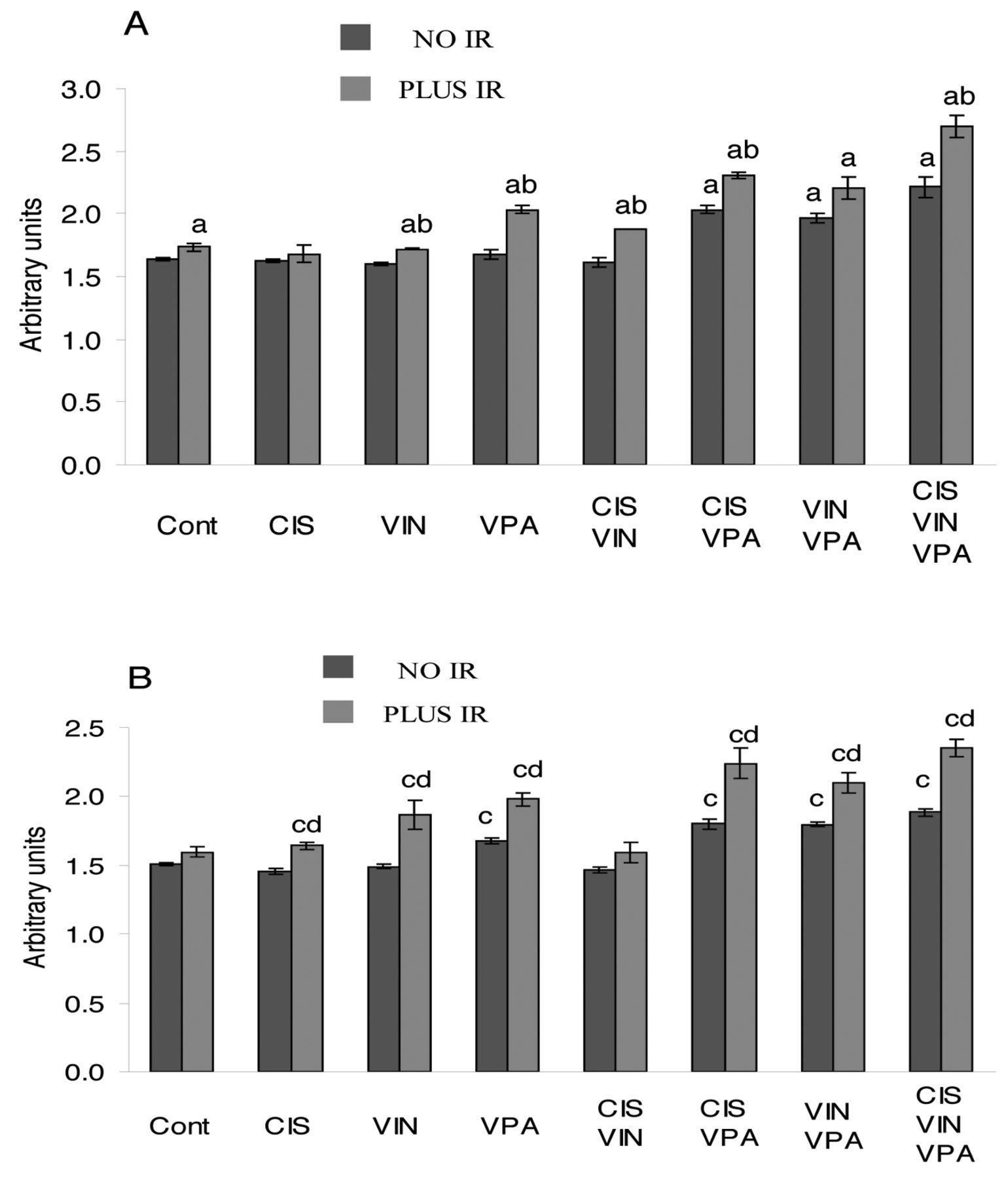
Expression of the cell-cycle inhibitors p21CIP1/WAF1 and p27KIP1. The results in Figure 5 show that only VPA as a single-agent significantly increased the p27KIP1 level (p<0.002). However, double combinations of VPA with cisplatin or vinorelbine, and the triple combination of cisplatin-vinorelbine-VPA were markedly more effective and considerably increased the level of p21CIP1/WAF1 (p<0.008) and p27KIP1 (p<0.04), compared to VPA treatment alone. IR applied alone slightly increased the level of p21CIP1/WAF1 (p<0.05) and p27KIP1 (p>0.08). However, co-application of IR with VPA or with the double combinations of cisplatin-VPA or vinorelbine-VPA significantly increased the level of p27KIP1 (p<0.02) compared to treatments without IR. Similar changes were observed in the p21CIP1/WAF1 level, except for the co-application of IR with vinorelbine-VPA treatment (p=0.08). The maximal levels of p21CIP1/WAF1 and p27KIP1 were achieved in cells treated with the four-agent combination of cisplatin-vinorelbine-VPA-IR: this treatment raised the p21CIP1/WAF1 level by 64.4% (p<0.0003) and the p27KIP1 level by 55.7% (p<0.0002).
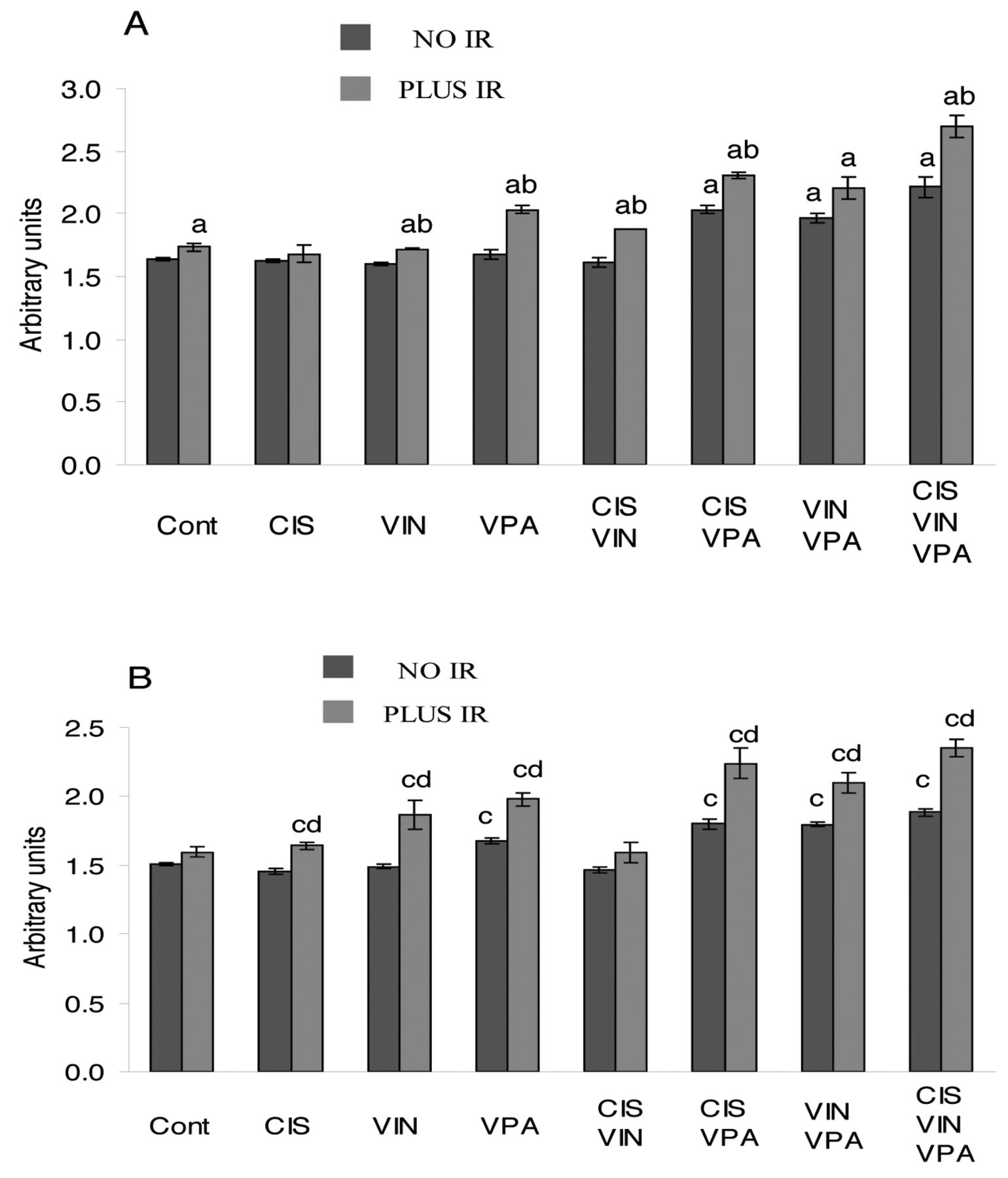
Effect of cisplatin (CIS), vinorelbine (VIN), sodium valproate (VPA) and ionizing radiation (IR) alone and in combined treatments on levels of p21CIP1/WAF1 and p27KIP1 in A549 cells. A: p21CIP1/WAF1, B: p27KIP1. ap<0.04 compared to untreated cells (Cont) for p21CIP1/WAF1, bp<0.02 compared to analogous treatment without IR for p21CIP1/WAF1. cp≤0.03 compared to untreated cells (Cont) for p27KIP1, dp<0.03 compared to analogous treatment without IR for p27KIP1.
Discussion
The present in vitro results showed that addition of VPA, an HDAC inhibitor, to the conventional treatment protocol consisting of the radiomimetic cisplatin, the anti-mitotic vinca alkaloid drug vinorelbine and IR substantially enhances their anticancer effect. Co-application of VPA with the combination of cisplatin-vinorelbine-IR reduced the growth of NSCLC A549 cells by 48.8%, while the same combination without VPA reduced A549 cell growth only by 34.8%. The difference between these treatments was highly significant.
The cell-cycle distribution experiments showed that the inhibition of A549 cell proliferation is mostly caused by enhanced apoptotic death. We found that the triple combination of cisplatin-vinorelbine-VPA followed by IR was the most effective and increased the percentage of apoptotic cells 12-fold. The suppression of cancer cell proliferation was also due to a delay of the S-phase transition.
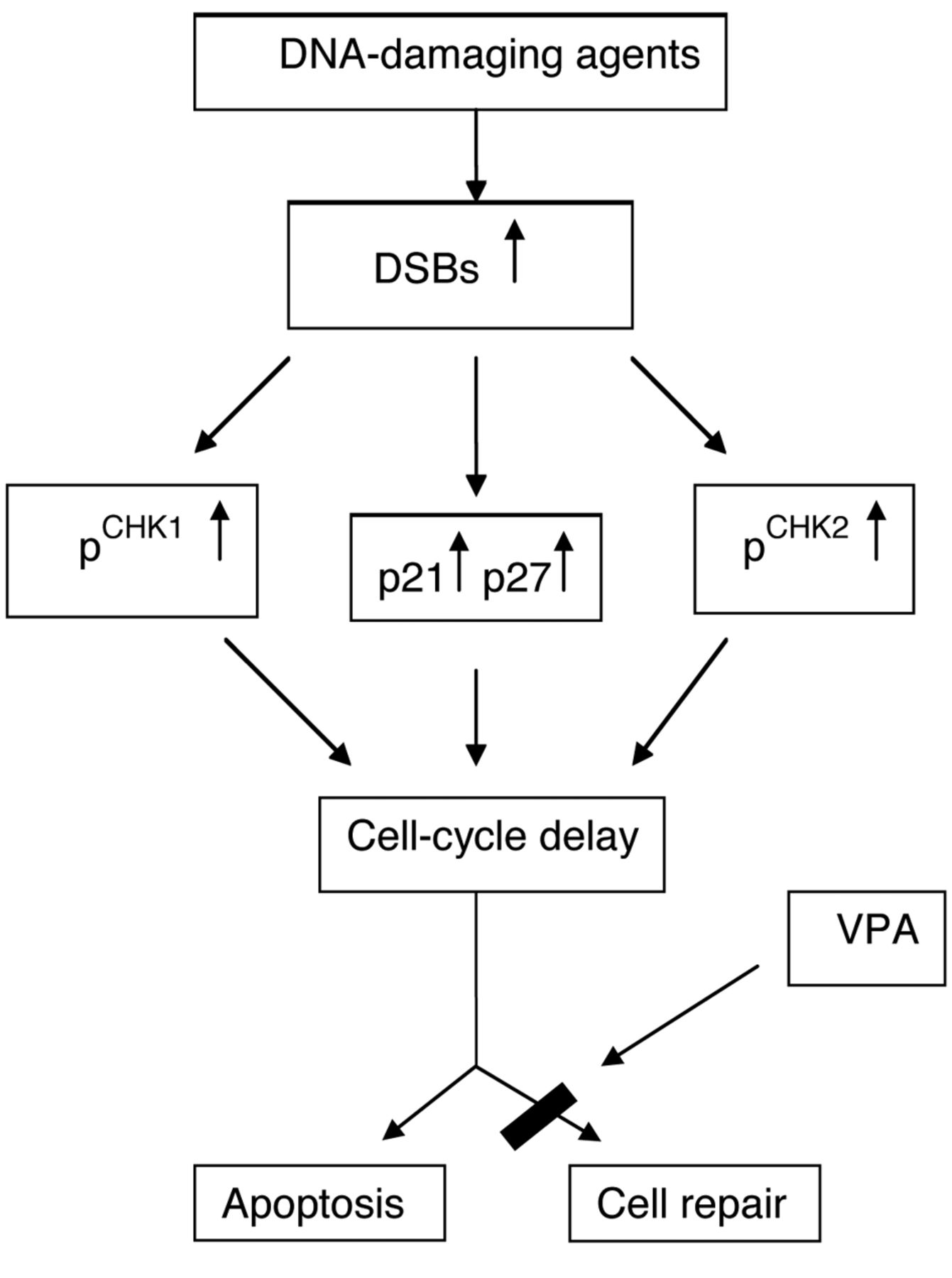
The present study demonstrates that both the enhanced apoptotic cell death and delay of cell-cycle progression are a consequence of the induction of the DDR signaling network (14) (Figure 6). The activation of ATM and ATR signaling pathways, the key elements of DDR, induces ‘checkpoint’ mechanisms (14) which delay or arrest the progression of cells through the cell-cycle and promote repair of DNA or apoptosis, if damage to DNA is severe and irreparable. Induction of DDR was demonstrated by elevated levels of γ-H2A.X, a hallmark of DSBs, and by the activation of CHK2 and CHK1 checkpoint kinases. The present findings indicate that combinations of cisplatin-VPA-IR and cisplatin-vinorelbine-VPA-IR were the most potent DNA-damaging treatments. The marked elevation of the level of DSBs could be explained by both the ability of IR and radiomimetic cisplatin to induce DSBs (15), and by the ability of VPA, an HDAC inhibitor, to blunt the capability of cancer cells to repair such damaged DNA (16). VPA itself also causes DSBs formation via the generation of reactive oxygen species (17). Activation of CHK2 was more prominent compared to CHK1. This finding is consistent with the well-established fact that DNA DSBs predominantly activate the ATM/CHK2 pathway, while single-stranded DNA breaks mainly activate the ATR/CHK1 signaling pathway (18). The present study showed that the anti-mitotic drug vinorelbine did not activate the DDR pathway. These results are supported by the study of Chiu et al. (19) who demonstrated that vinorelbine induces apoptosis independently of ATR and ATM pathways.

DNA damage response network activation. Induced DNA damage causes activation (phosphorylation) of the DNA damage checkpoint kinases CHK2, CHK1 and up-regulates the cyclin-dependent kinase inhibitors p21CIP1/WAF1 and p27KIP1. This results in cell-cycle progression delay and promotes DNA repair. Cells with irreparable damage are eliminated by apoptosis. DSBs – DNA double-strand breaks, VPA – sodium valproate.
Our findings revealed elevated levels of the cyclin-dependent kinase inhibitors p21CIP1/WAF1 and p27KIP1, especially in cells treated with the cisplatin-vinorelbine-VPA-IR combination. These results are consistent with the observation that DNA damage induces up-regulation of the cyclin-dependent kinase inhibitors p21CIP1/WAF1 and p27KIP1 (20). In turn, increased expression of p21CIP1/WAF1 and p27KIP1 promotes cell-cycle delay, providing a critical window-in-time for the repair of damaged DNA.
In conclusion, the present findings show that the addition of VPA, an HDAC inhibitor, to the conventional clinical treatment comprising cisplatin-vinorelbine followed by IR significantly enhances inhibition of NSCLC cell growth. This observation may provide a new applicable protocol for enhancing the efficacy of the combined anticancer chemoradiation treatment and for reducing the doses of IR and anticancer drugs required, thus alleviating the severity of side-effects in patients with locally advanced NSCLC.
Acknowledgements
This research was supported by joint grant of the Maria Rossi Ascoli Foundation and the Faculty of Health Sciences, Ben-Gurion University of the Negev, Israel.
- Received June 18, 2014.
- Revision received August 2, 2014.
- Accepted August 6, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}