Iron Metabolism in Cancer


1
Division of Anti-Tumor Pharmacology, State Key Laboratory of Drug Research, Shanghai Institute of Materia Medica, Chinese Academy of Sciences, Shanghai 201203, China
2
University of Chinese Academy of Sciences, Beijing 100049, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2019, 20(1), 95; https://doi.org/10.3390/ijms20010095
Submission received: 26 November 2018
/
Revised: 21 December 2018
/
Accepted: 22 December 2018
/
Published: 27 December 2018
(This article belongs to the Special Issue Drug Discovery, Development and Regulatory Affairs)
Abstract
:Demanded as an essential trace element that supports cell growth and basic functions, iron can be harmful and cancerogenic though. By exchanging between its different oxidized forms, iron overload induces free radical formation, lipid peroxidation, DNA, and protein damages, leading to carcinogenesis or ferroptosis. Iron also plays profound roles in modulating tumor microenvironment and metastasis, maintaining genomic stability and controlling epigenetics. in order to meet the high requirement of iron, neoplastic cells have remodeled iron metabolism pathways, including acquisition, storage, and efflux, which makes manipulating iron homeostasis a considerable approach for cancer therapy. Several iron chelators and iron oxide nanoparticles (IONPs) has recently been developed for cancer intervention and presented considerable effects. This review summarizes some latest findings about iron metabolism function and regulation mechanism in cancer and the application of iron chelators and IONPs in cancer diagnosis and therapy.

{kind=link}
{kind=link}
{kind=link}
1. Introduction
The past decade has been described as the “golden age” of iron metabolism, due to the discovery of new iron-related proteins and regulatory mechanisms [1]. Efforts have been made to decipher physiological and molecular function of iron in cancer development. Multiple iron metabolism-associated proteins have been proved to participate in malignant tumor initiation, proliferation, and metastasis. Compared to normal cells, tumor cells differ in the expressions or activities of many iron-related proteins. These alterations generally contribute to a relatively high level of intracellular iron availability and facilitate the functions of iron-dependent proteins, which are involved in numerous physiological processes including DNA synthesis and repair, cell cycle regulation, angiogenesis, metastasis, tumor microenvironment, and epigenetic remodeling [2,3]. Consequently, iron homeostasis modulations including iron depletion and iron metabolism-targeted treatments have exhibit potent and broad anti-tumor effect, which makes it a potential and largely undeveloped therapeutic target for cancer pharmacological therapy. Some iron chelators and IONPs have already been put into clinical evaluation for curing hematological malignancies and other cancer types, and have lately shown enormous potential in combination with traditional chemotherapy and emerging immunotherapy [4,5]. Nevertheless, there exists more to be mined about iron homeostasis regulation and its role in cancer physiology, and ways to make it glow in cancer treatment. In this review, we integrate some latest expounded iron metabolism pathways and its major physiologies associated with cancer progression, tumor microenvironment, and epigenetic regulation. We then summarize some novel iron modulators in development and iron chelators in combined therapy, which could provide new therapeutic options for cancer intervention.
2. Regulation of Iron Homeostasis in Cancer
As a trace element, iron is necessary for cell basic function and especially highly required for malignant cancer cells, in which some pivotal changes about iron import and output have been identified. Generally, iron in the systemic iron pool is bound to transferrin (TF). Then, iron-loaded TF forms complex with transferrin receptor 1 (TfR-1) on the cell plasma membrane, which is internalized by endocytosis [6,7]. Whereas cancer cells have some alterant pathways in maintaining cellular iron balance. In non-small-cell lung carcinoma cells (NSCLC), epidermal growth factor receptor (EGFR) is demonstrated to affect iron metabolism by directly binding and re-distributing TfR-1. EGFR inactivation reduces TfR-1 level on the cellular surface, engendering iron import decrease and cell cycle arrest [8]. CD133 (cluster of differentiation 133), the pentaspan stem cell marker and a marker of tumor-initiating cells in a number of human cancers, can also inhibit iron intracellular uptake by interacting with TfR-1 and implicating in its endocytosis, thus participating in iron metabolism [9].
In the endosome, Fe3+ is reduced to Fe2+ by iron reductase, mainly by some members of the metalloreductases six-transmembrane epithelial antigen of prostate (STEAP1-4) family [10,11]. STEAP1 and STEAP2 are highly expressed in various human cancer types, such as colon, breast, cervix, prostate, pancreas, bladder, ovary, testis, and Ewing sarcoma [12,13,14]. STEAP3 is overexpressed in malignant gliomas, and STEAP3 knockdown suppresses glioma cell proliferation, clonality and metastasis in vitro and tumor growth in vivo. STEAP3 induces cancer epithelial–mesenchymal transition (EMT) by activating STAT3-FoxM1 axis, promoting TfR-1 expression and thus elevating cellular iron content [15]. STEAP4 is activated under hypoxia condition and leads to mitochondrial iron imbalance, enhances reactive oxygen species (ROS) production, and increases the incidence of colitis-associated colon cancer in mouse models [16]. Several promising STEAPs-targeting strategies in cancer therapy include monoclonal antibodies (mAbs), antibody-drug conjugates, DNA and small noncoding RNAs (ncRNAs) vaccines [17,18]. Once Fe3+ has been reduced to Fe2+ in the endosome, it is transported across the endosome into the cytosol via divalent metal-ion transporter 1 (DMT1), Zrt- and Irt-like protein 14 (ZIP14) or ZIP8 [19,20]. DMT1 functions as a main iron transporter and pharmacological inhibition of DMT1 suppresses colon tumor growth by suppressing JAK-STAT3 signaling [21,22]. The iron obtained through DMT1 constitutes the cytoplasmic labile iron pool (LIP) in which iron is metabolically active.
Most iron in the active form is finally utilized in various physiological processes such as DNA synthesis, mitochondrial oxidative metabolism and cytoplasmic ferritin for storage. Ferritin is an iron-containing protein with multiple functions in iron delivery, cell proliferation, angiogenesis, and immunosuppression. Under the case of cancer, ferritin is detected in high concentration in plasma in many patients, and its higher level correlates with higher clinical tumor stage and poorer patients’ outcome [23,24]. Iron–sulfur biogenesis is another common form of iron utilization. NEET proteins belong to a novel iron–sulfur (2Fe-2S) protein family that regulate iron and redox homeostasis and are involved in cancer progression. It has been revealed that NEET proteins NAF-1 and mitoNEET can promote cancer cell proliferation and metastasis by increasing mitochondrial iron accumulation. They represent a key regulatory link among the maintenance of high iron and ROS level in cancer cells [25,26].
Excess iron that is not utilized or stored can be exported across the membrane through ferroportin (FPN), an only-known iron efflux pump cooperated with ferroxidases named hephaestin (HEPH) or ceruloplasmin (CP) to maintain cellular iron homeostasis [27,28]. FPN is dramatically suppressed in many cancer types [29]. FPN overexpression induces autophagy and activates p53 and its downstream target p21, thus causing cell cycle arrest and stress-induced DNA-damage in prostate cancer [30]. Reduced FPN level in triple-negative breast cancer cells (TNBC) stimulates proliferation and epithelial-mesenchymal transition (EMT) as indicated by increased E-cadherin and decreased N-cadherin, Twist and Slug expression [31]. Some metal elements have been reported to modulate FPN’s transport activity, such as Ca2+ and Cadmium (Cd) [31,32]. These findings enrich our knowledge of FPN in mediating iron output and are conducive to the strategy development of manipulating FPN therapeutically in cancer.
Iron homeostasis has been demonstrated to be regulated under different levels. On the cellular level, iron metabolism is predominantly under post-transcriptional control by the iron responsive element-iron regulatory protein (IRE-IRP) system [33,34,35]. Under iron-low conditions, IRPs bind to IREs of mRNAs encoding ferritin subunits, FPN, DMT1 and TfR-1. Binding stabilizes TfR-1 and DMT1 mRNAs, whilst inhibits ferritin and FPN translation, which leads to an elevation in iron uptake and availability and a reduction in iron storage and efflux [36,37,38]. On the systemic level, iron homeostasis is mainly governed by hepcidin, a key iron sensing and regulatory hormone. Hepcidin facilitates FPN degradation and thus prevents iron export from gut enterocytes, reticuloendothelial cells (macrophages) and hepatocytes into circulation [39]. Hepcidin synthesized by tumors or liver contributes to cancer proliferation and progression. Significant genetic variants in the BMP/Smad4/Hamp hepcidin-regulating pathway could help predict the outcomes in NSCLC patients under definitive radiotherapy [40]. Studies have shown that regulating hepcidin level to reduce iron availability in the neoplastic cells may be a novel strategy in the anticancer treatment [41]. As acknowledged in tumor cells, proteins those raise intracellular iron contents (TfR-1, DMT1, hepcidin) are extensively upregulated, whereas those lower iron levels (FPN, hephaestin (HEPH)) are downregulated. Otherwise, they are identified as promising predictors for the clinical prognosis of patients with breast, liver, lung, colon, brain, prostate, ovarian, gastric and pancreatic cancer, leukemia, and glioma [19,42,43,44,45].
3. Iron and Epigenetic Regulation in Cancer
3.1. Iron Plays an Important Role in Cancer Epigenetics
When exist in redox-active form, iron can catalyze Fenton-type reactions and produce highly reactive free radicals, able to oxidize and attack most cellular components [46]. Fe-S cluster contributes the active center of various enzymes that are necessary for mitochondrial oxidative metabolism and epigenetic modulation. Its biogenesis disruption results in decreased overall histone acetylation by decreasing levels of histone acetyltransferase ELP3 (elongator complex protein 3) and succinate dehydrogenase, increased DNA methylation by reducing the activity of DNA demethyltransferase DME, and increased acetylation of a-tubulin in the cytosol by elevating the tubulin acetyltransferase MEC17 [47,48]. Moreover, an unbiased genetic screen shows that maintenance of iron homeostasis is sufficient to stabilize facultative heterochromatin assembly and appropriate genome-wide gene control. Iron overload may reshape the genome and epigenome by disrupting p53-mediated DNA repair pathway and enhancing DNA hypomethylation, forming a feasible environment for transposons or transposable elements [49,50]. These results uncover the mechanism of iron in globally remodeling the genome in a dynamic way.
Iron depletion has been shown to induce global histone and DNA methylation changes in cancer cells without affecting expression levels of histone and DNA methyltransferases or demethylases, implying the influence on their enzymatic activity [51,52]. Iron- and 2-oxoglutarate (2-OG)-dependent oxidative histone demethylation mediated by JmjC family are such enzymes, whose dynamic processes have been linked to the epigenetic regulation of tumor formation and development [53,54]. For instance, the JmjC-domain-containing protein JHDM2A/KDM3A, that specifically demethylates histone H3 at lysine 9 (H3K9), is iron-dependent and regulates androgen receptor transcription in prostate cancer cells [55]. JARID1B/KDM5B, which selectively demethylates H3K4me3, serves as a good prognosis predictor in NSCLC, breast and ovarian cancer [56,57,58]. The enzymatic activities of the JmjC family are significantly weakened in the absence of α-ketoglutarate, Fe (II) or ascorbic acid, or in the presence of iron chelators. Moreover, ten-eleven translocation (TET) proteins are also Fe(II)- and 2-OG-dependent dioxygenases that oxidize 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC), and 5-carboxylcytosine (5caC), thereby generating active DNA demethylation, which is linked to aberrant silencing of tumor suppressor genes in cancer [59,60]. The deprivation of cellular LIP using specific chelator blocks the Tet-mediated 5mC oxidation [61]. Thus, iron is directly participated in the nucleus to promote oxidative demethylation of correlative DNA and histone residues throughout chromatin, which has been revealed to control cancer EMT status by regulating the expression of related proteins in a dynamic manner [62].
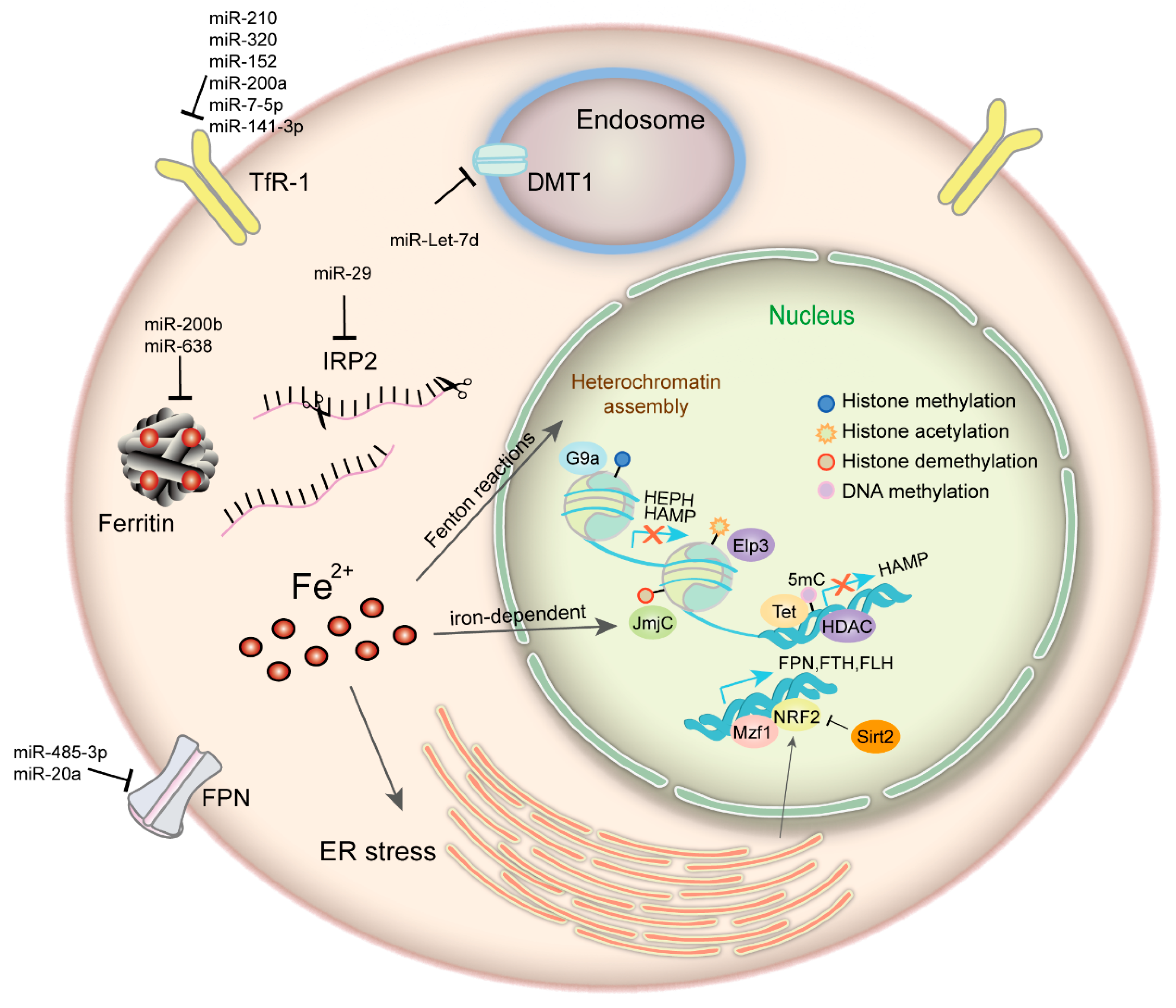
3.2. Epigenetic Regulation of Iron Metabolism-Related Proteins
From the above, iron makes a difference to epigenetics, whilst iron homeostasis is under epigenetic regulation which may take diverse forms, such as DNA methylation, histone modification, post-transcriptional control and some transcription factors working as cooperators (Figure 1). The HAMP gene, which encodes hepcidin, is transcriptionally repressed and closely associated with the hypermethylated signature on the gene promoter region in human hepatocellular carcinoma [63]. Histone deacetylase 1 (HDAC1) is defined as a novel hepcidin suppressor by binding to SMAD4, effect of which is independent of BMP/SMAD1/5/8 signaling and without deacetylation of SMAD4 or histone-H3 on the hepcidin promoter [64]. Histone deacetylase 3 (HDAC3) and its cofactor NCOR1 can also inhibit hepcidin expression, involving reversible loss of H3K9ac and H3K4me3 at the hepcidin gene promoter [65]. Hepcidin is also regulated by the E4BP4/G9a/SOSTDC1/hepcidin pathway which causes cellular iron dysfunction and is an essential link in thyroid cancer growth [66]. Apart from hepcidin, there are other iron-metabolism-related proteins under epigenetic control being identified. FTH is manipulated through histone acetylation by MBD5 (a methyl-CpG binding protein) forming complex with histone acetyltransferase KAT2A (lysine acetyltransferase 2A) [67]. Another MBD family member, methyl-CpG binding protein 2 (MECP2) deficiency is reported to mediate brain iron metabolism by inducing oxidative stress [68]. Our previous study shows that ferroxidase hephaestin (HEPH) is repressed by G9a, a H3K9 methyltransferase, which forms complex with transcription factor YY1 and HDAC1, leading to cellular LIP increase and promoting breast cancer proliferation [69]. The activity of IRP1 is modulated by NAD-dependent deacetylase sirtuin-3 (mitochondrial SIRT3), thus affects cellular iron metabolism. SIRT3 loss increases ROS production, bringing about elevated IRP1 binding to IREs and increased TfR-1 expression as a consequence [70]. Mitochondrial ferritin (FTMT) appears regulated by a complex mechanism involving epigenetic events, such as histone de-acetylases recruitment and GC islands accumulation at its promoter, and interplay between transcription factors, such as CREB, SP1, and YY1 [71]. Nuclear factor erythroid 2-like 2 (NRF2) and myeloid zinc finger-1 (MZF-1) could impact cancer cell growth by transcriptionally regulating FPN, FTH, and FTL expression in prostate and breast cancer [72]. FPN transcription is inhibited by transcription factor BACH1 (Btb and Cnc Homology 1) and activated by NRF2 [73]. Deacetylase SIRT2 can deacetylate and repress NRF2 nuclear localization, reducing FPN expression and iron export, thus maintaining cancer cellular iron levels [74]. While BACH1 is involved in iron and heme regulatory system since it is inactivated through directly binding to heme in erythroblast and K562 cells [75].
A set of miRNAs that functions in RNA silencing and post-transcriptional regulation of gene expression have been demonstrated to be involved in the regulation of iron metabolism. According to the recent studies, some miRNA levels correlates negatively with iron intake. In vitro experiments indicate that enhanced miR-320 level can repress TfR-1 expression and inhibit cell proliferation [76]. miR-7-5p and miR-141-3p target 3'-TfR-1 IREs and downregulate TfR-1 mRNA and protein level [77]. miR-210, miR-200a, miR-152 can also repress TfR-1 expression [78,79,80]. In erythroid cells, miR-Let-7d is involved in the refined regulation of iron uptake by targeting DMT1-IRE isoform. They are all inversely correlated during erythroid differentiation of CD34+ cells, K562 and HEL cells [81]. miRNAs in some way can also modulate iron utilization and storage. miR-638 overexpression reduces FTH protein level in prostate cancer [82]. miR-200b inhibits FTH expression and increases cancer cells sensitivity to chemotherapy [83]. The Fe-S cluster assembly protein (ISCU1/2) is suppressed by miR-210, leading to Fe-S cluster biogenesis disruption and iron metabolism alterations [84]. Furthermore, the liver specific miR-122 directly targets HFE and HJV and contributes to the regulation of systemic iron homeostasis by decreasing hepcidin mRNA expression [85]. The iron-binding glycoprotein lactoferrin has been identified as a direct target of miR-214 in MCF-7 cells. Nevertheless, its receptor is post-transcriptionally regulated by miR-584 in Caco-2 cells [86,87]. Except for regulating iron uptake, utilization and storage, miRNAs also take part in iron export regulation. miR-20a inhibits FPN by binding to highly conserved target sites in its 3′-UTR (untranslated region) [88]. Reinforced expression of miR-20a may diminish iron efflux, contributing to intracellular iron retention, which favors lung cancer cell growth and proliferation. miR-485-3p overexpression can also repress FPN expression and give rise to elevated intracellular ferritin levels and LIP content [89]. To be concluded, iron metabolism and epigenetic control are closely interplayed and mutual restricted in cancer.
4. Role of Iron in Tumor Cell Biology
4.1. Iron in Cancer Cell Cycle and DNA Metabolism
Due to higher rates of proliferation and DNA synthesis, neoplastic cells have higher requirements of iron than normal cells. Functioning in various electron transfer systems, iron is indispensable in maintaining the activity of iron- or heme-containing enzymes. They refer to amounts family of ferrous iron dioxygenases and each possesses active site dependent on relatively labile iron, including enzymes involved in DNA replication and repair, Jumonji (JmjC) domain-containing histone demethylases involved in epigenetic modification and mitochondrial oxidases involved in respiratory complexes [90,91,92]. To be more detailed, the proteins participated in DNA synthesis and repair procedures include DNA polymerases, DNA helicases, and the small catalytically active subunit of ribonucleotide reductase (RRM2) [92,93]. Moreover, mammalian cells contain a RRM2 subunit which is p53-inducible (p53R2) and can be activated in case of DNA damage. Interestingly, p53R2 is vulnerable to iron depletion, precipitating it into a potential target for iron chelation therapy in tumors with wild-type p53 [91]. Meanwhile, it is known that ROS generated in the Fenton reaction by excess iron can attack DNA, causing mutations and damages, inactivating tumor suppressor genes or activating oncogenes. Furthermore, iron plays a crucial role in regulating cell cycle by affecting both the formation and activity of the cyclin proteins (cyclin A, B, D, and E) and cyclin-dependent kinase (CDKs) complexes. Intracellular iron depletion by chelators results in hypo-phosphorylation of the retinoblastoma protein (pRb), decreased expressions of cyclins, p21 and c-myc, thus causes G1/S cell cycle arrest particularly [94,95]. Nevertheless, other researches also show that IRP2 depletion give rise to induction of p15, p21, and p27, leading to prostate cancer cells accumulation in G0/G1 [96]. Collectively, iron presents a close relationship with DNA metabolism and cell cycle process.
4.2. Iron in Cancer Cell Demise
To keep cellular redox homeostasis, iron and ROS levels are both strictly manipulated. When the balance is disrupted, the cell may go into canceration or demise. Since its first demonstration in 2012, ferroptosis has been well characterized as a type of programmed cell death caused by accumulation of iron-induced lipid peroxidation and metabolic constraints, which is genetically and biochemically distinct from apoptosis, autophagy, necroptosis, and necrosis [97,98]. Ferroptosis has been reported to be able to inhibit some cancer types, such as hepatocellular carcinoma (HCC), pancreatic carcinoma, breast cancer, and prostate cancer [99]. The glutathione (GSH) redox system is pivotal to restrain ferroptosis under the case of impaired lipid metabolism. Inhibition of GSH synthesis or glutathione peroxidase 4 (GPX4) is sufficient to trigger ferroptotic cell death [100]. The Fe-S cluster biosynthetic enzyme NFS1 is also critical to prevent ferroptosis by restricting iron influx from intracellular stores, cooperating with GSH synthesis [101]. Furthermore, wild-type p53 negatively regulates the expression of the cystine importer SLC7A11, which increases cell sensitivity to ferroptosis. p53 can also suppress ferroptosis through transcription-dependent and -independent mechanisms, implying the bidirectional and context-dependent control of ferroptosis and oxidative stress by p53 [102]. Recent studies reveal that the ferroptotic agent-induced endoplasmic reticulum (ER) stress contributes to the cross-talk among ferroptosis and other types of cell death, such as apoptosis [103]. Ferroptotic and apoptotic agents interact through the PERK-eIF2a-ATF4-CHOP-PUMA pathway caused by ER stress response and effectively enhance each tumoricidal efficacy, implicating a novel combined therapeutic strategy for cancer [103]. Furthermore, since cancer stem cells (CSCs) are iron-rich and iron-dependent, ferroptotic agent salinomycin and its derivative, ironomycin, exhibit selective and potent therapeutic effect against breast CSCs by accumulating iron-mediated lysosomal ROS via Fenton reaction and causing ferroptosis [104,105]. These unprecedented findings manifest the druggability of remodeling iron homeostasis in the context of CSCs.
Ferritinophagy refers to the autophagic degradation of ferritin protein that reserves iron and maintains balance when iron is depleted [106]. The process is mediated by an autophagy cargo receptor named nuclear receptor coactivator 4 (NCOA4), which binds to ferritin heavy chain (FTH) in the autophagosome and delivers it for degradation in the lysosome, thus releasing iron for physiological demands [107]. It has been revealed that ferritinophagy evokes an iron-driven intra-lysosomal oxidative reaction, resulting in LIP upregulation and lipid peroxidation, and finally cell demise [108].
Another iron-oxy-related cell death is ferrosenescence. It is defined as follows: excess iron directly degrades p53 and blocks the p53-mediated DNA repair, causing genomic disintegration and DNA damage in the cell. Aside from genomic alterations, ferrosenescence generates epigenomic changes by inducing global DNA hypomethylation through upregulating DNA methyltransferase 3A and 3B (DNMT-3A and DNMT-3B) and remobilizing transposable elements through miR-29/p53 pathway [50,109]. The ferrosenescence-associated genome disintegration eventually leads to cell demise by ferroptosis or apoptosis.
4.3. Iron in Tumor Metastasis and Angiogenesis
Iron plays an important role in matrix degradation and cancer metastasis by stimulating or stabilizing some metalloprotease activities. Iron overload increases metalloprotease-2 (MMP-2) and metalloprotease-9 (MMP-9) activity in a dose-dependent manner partly through activation of AP-1 via ERK/Akt pathway [110]. A recent study shows that FPN overexpression attenuates the LIP and ROS production and inhibits EMT, as reflected by significantly decreased representative EMT markers, such as SNAIL1, TWIST1, ZEB2, and vimentin [31]. FTH overexpression also leads to a suppression of EMT, which suggests that labile iron is beneficial for tumor migration [111]. However, some studies indicate that iron may inhibit vascular endothelial growth factor (VEGF)-induced endothelial cell proliferation, migration, tube formation, and sprouting [112]. Moreover, iron deficiency significantly promotes VEGF expression by stabilizing hypoxia-inducible factor-1α (HIF-1α) [113]. Therefore, iron has two sides on tumor metastasis under different circumstances which needs further research.
The iron-regulated metastasis suppressor N-myc down-stream-regulated gene 1 (NDRG1) is a well-known metastasis suppressor that decreases metastases and improves patient prognosis in breast, prostate, pancreas, and colon cancer [114,115]. Another NDRG family protein, the Myc-repressed gene NDRG2, though not as extensively studied as NDRG1, has demonstrated tumor-suppressive functions in malignant carcinomas [116,117]. Similar to NDRG1, NDRG2 is also upregulated under iron depletion. NDRG2 can reduce the level of receptor gp130 and inactivate its downstream targets STAT3 and ERK1/2, which leads to decreased EMT and tumor metastasis [118].
More iron-related proteins participating in tumor metastasis and angiogenesis include follows: collagen lysyl hydroxylases (LH1-3), a Fe2+- and 2-oxoglutarate (2-OG)-dependent oxygenases can maintain extracellular matrix homeostasis and cell migration potential [119]; β2-microglobulin (β2-M) interacts with its receptor, hemochromatosis (HFE) protein, stimulates iron responsive HIF-1α signaling pathway and promotes cancer bone and soft tissue migration [120]; inflammatory mediator Lcn2 increases tube formation, cell migration, and angiogenesis in rat brain endothelial cells via iron and ROS-dependent mechanisms [121]. All these suggest a significant role of iron in tumor progression.
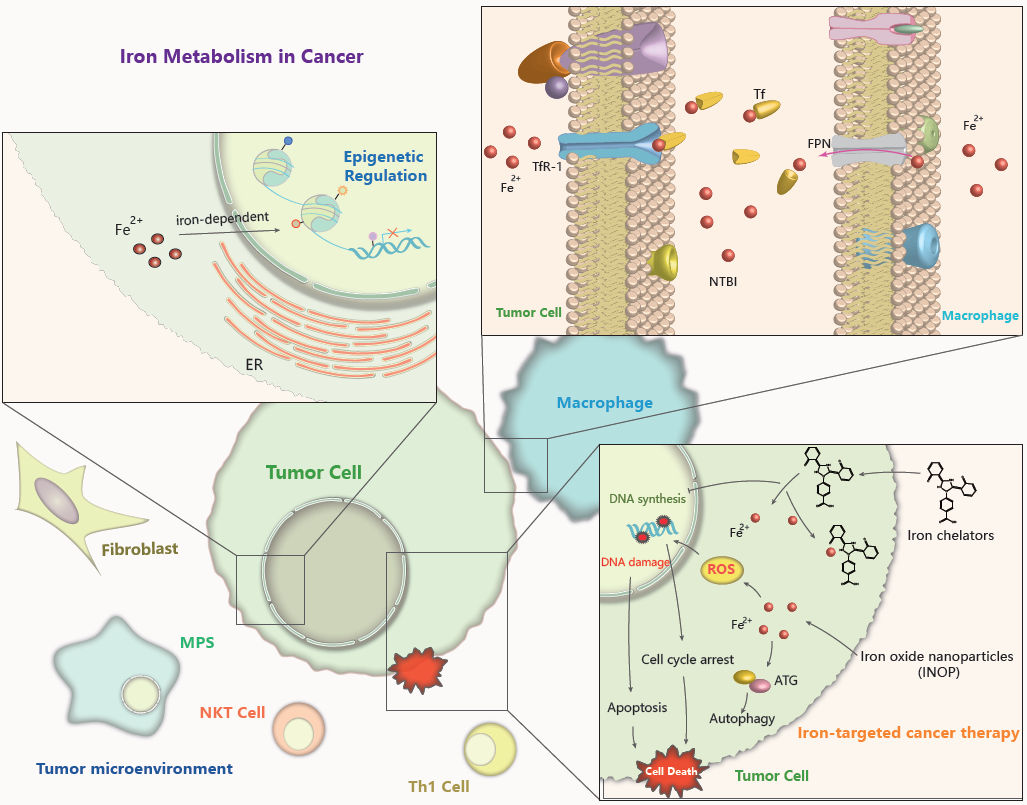
4.4. Iron in the Tumor Microenvironment
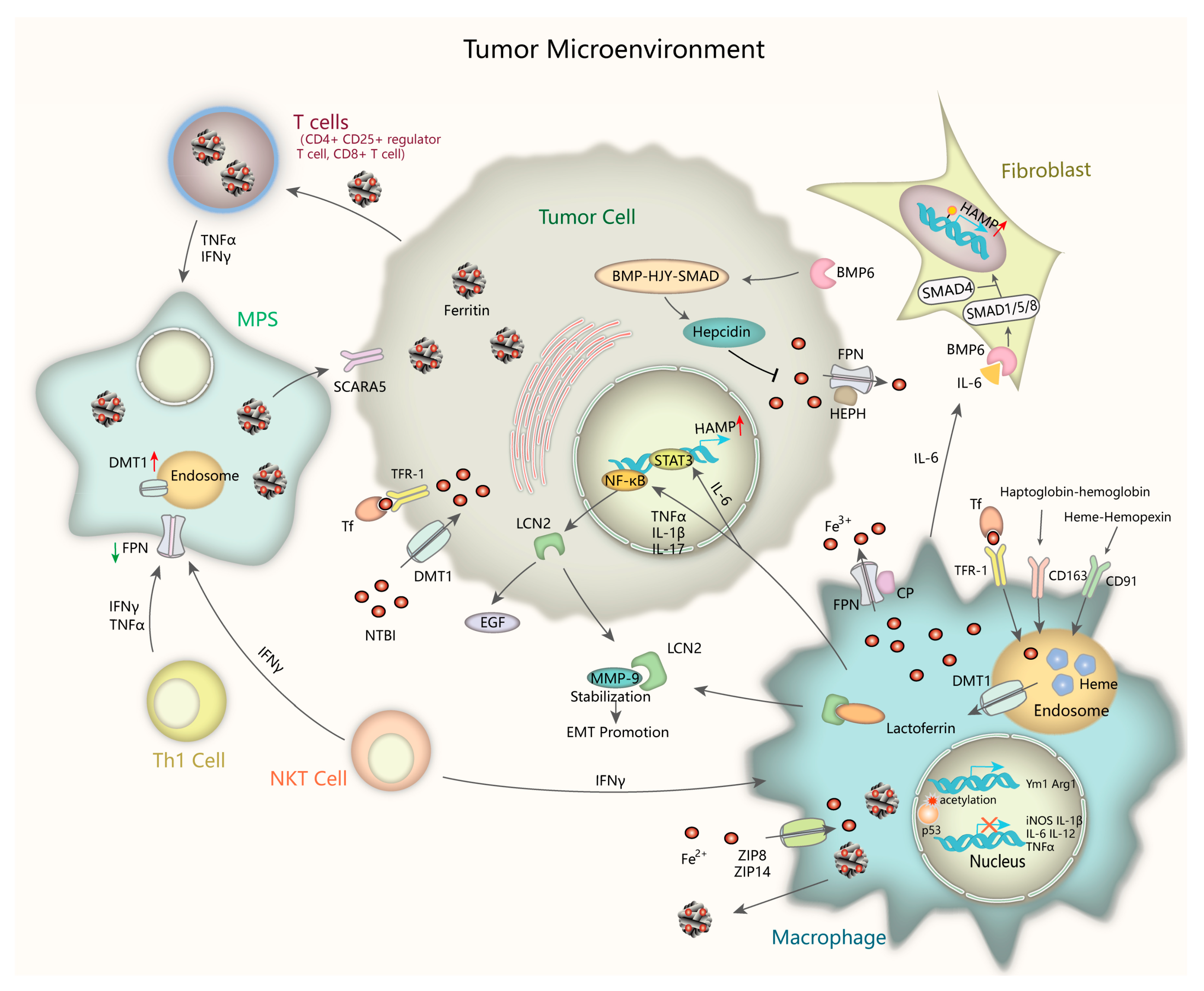
The tumor microenvironment refers to the extracellular matrix (ECM), other non-malignant cells such as immune cells, surrounding blood vessels, also signaling molecules and cytokines around the tumor cells. Cancer initiation and progression largely depend on extrinsic signaling from their cell niche. In the past several years, a more detailed understanding of the interaction between iron metabolism and tumor microenvironment has been addressed (Figure 2). Inflammatory stressors in the tumor microenvironment play a critical role in controlling iron metabolism and homeostatic pathway [122]. Elevated iron in cancer cells and pericarcinomatous compartments protect cancer cells from natural killer cell cytolysis by upregulating ferritin expression and by antagonizing tumor necrosis factor (TNFα)- and NO-induced cytotoxicity [123]. Immunologic factors like interleukin-6(IL-6) cause significant upregulation of hepcidin through IL-6-STAT3 pathway, results in intestinal iron uptake suppression and serum ferritin levels elevation [124]. The relationships among cancer, inflammation, and iron-related proteins such as ferritin could be complex, with ferritin either indicating iron overload that causes cancer, or indicating inflammation that causes cancer [125]. It has been revealed that the dysregulation of iron-related proteins in cancer cells, macrophages and lymphocytes are correlated with clinicopathological markers of poor patients’ outcome, such as hormone receptor absence and tumor metastasis presence, extending the meaning of iron homeostasis in the tumor microenvironment [126].
The mononuclear phagocyte system (MPS, including macrophages, monocytes, and their precursor cells) dramatically participate in maintaining iron homeostasis by recycling iron from hemoglobin of damaged or senescent erythrocytes [127]. Cytokines like interferon-γ (IFN-γ) and tumor necrosis factor (TNF) secreted by many cell types, including Th1 cells, natural killer T (NKT) cells, monocytes, and macrophages, increase DMT1 expression whereas decrease FPN level, thus resulting in iron sequestration in the MPS. Macrophages, cancer cell and T-lymphocytes uptake and reserve non-transferrin bound iron (NTBI) through non-transferrin-bound iron transporters such as ZIP14 and DMT1, functioning as circulating iron isoforms to avoid different tissues from iron-induced cytotoxicity [128,129,130]. What we known is that macrophages play the leading role in taking up, metabolizing, storing, and releasing iron. Classically activated macrophages (M1 macrophages) sequester iron by absorbing iron-loaded TF via TfR-1 or taking up Fe2+ via zinc transporters ZIP8 and ZIP14 [131,132]. In alternatively activated macrophages (M2 macrophages), the major component of tumor-associated macrophages (TAMs), consume of hemopexin-heme via CD91 or haptoglobin-hemoglobin via CD163 into endosomes as well as phagocytosis of senescent erythrocytes into erythrophagosomes lead to heme accumulation in the cytosol, which is another important source of iron [133,134,135]. Intracellular Fe2+ is oxidized to Fe3+ by ferroxidase ceruloplasmin (CP) and exported from M2 macrophages through FPN, then binds to TF efficiently. Iron efflux via FPN is blocked by hepcidin through degradation, thus increasing iron storage in intracellular ferritin. An alternative channel of iron export is secretion of ferritin-bound iron which is abundant in TAMs. It has been revealed iron-enriched status increases M2 phenotype marker Arg1 and Ym1 expression and promotes M2 polarization, whereas it represses M1 proinflammatory response [136,137]. Though a recent study has pointed out that iron overload induces macrophage polarization to a pro-inflammatory phenotype through promoting ROS production, enhancing p300/CBP acetyltransferase activity and increasing p53 acetylation [138], iron indeed modulates the inflammatory response outcome. Iron-loaded TAMs (iTAMs) infiltration is reported to correlate with tumor regression in NSCLC patients. Iron delivery system targeted to TAMs has been proven to be an effective adjuvant therapeutic strategy to reinforce anti-tumor immune responses [139].
From the foregoing, ferritin is rich in TAMs, which have recently been demonstrated to have critical roles in tumor progression and drug resistance [140]. Inflammatory cytokines can regulate the expression of ferritin on two levels: transcriptional level (principally H-ferritin, FTH) and translational level (FTH and L-ferritin, FTL) [141]. FTH is highly expressed in the melanoma patients’ serum and is correlated with increased circulating CD4+CD25+ regulator T cells, contributing to their immune functions [142]. Further studies indicated that the proliferation of T cells such as CD8+ T cells requires intracellular iron stored in FTH [143]. FTL gathered in cancer lesions to promote proliferation is potentially taken up from plasma via specific receptors, such as scavenger receptor class A member-5 (SCARA5) from the release of TAMs, particularly in response to pro-inflammatory cytokines [140,144]. The increased iron uptake reinforces the IL-6 paracrine loop between TAMs and breast cancer cells, leading to intensive de novo acquired chemo-resistance [145]. Lactoferrin (Lf), an iron-binding glycoprotein, has multiple functions in innate immunomodulation by modulating cytokines production of granulocyte macrophage colony stimulating factor (GM-CSF), IL-1, TNF, and IL-6 by macrophages, regulating natural killer cell activity, and inhibiting antibody synthesis and T cells maturation [146,147].
The innate immune protein Lipocalin 2 (Lcn2) has emerged as a critical iron regulatory protein under tumorigenic and inflammatory conditions. TNF-α, IL-17, and IL-1β secreted from TAMs can induce Lcn2 expression by activating NF-κB pathway, suggesting that Lcn2 is a type I acute phase protein [148,149,150]. Lcn2 deficiency is shown to accentuate spontaneous colitis and promote colonic tumorigenesis in IL-10-deficient mice [151]. Lcn2 can also be released into the extracellular matrix and promote iron internalization and sequestration through known receptors such as megalin, contributing to cancer cell survival and metastasis. Mechanically, Lcn2 stabilizes and binds to MMP-9, resulting in matrix degradation and tumor EMT [152]. TAMs can secret Lcn2 and elevate intracellular iron concentration in tumor cells via Lcn2 as transporter [153]. In the TAMs, Lcn2 can colocalize with lactoferrin and release pro-inflammatory cytokines into the microenvironment [154]. It can also be stored as monomers or homodimers in the neutrophil-specific granules [155]. These concomitant events present the tumorigenic and immunological effects of Lcn2, thus facilitating tumor growth and metastasis. Neutrophil gelatinase-associated lipocalin (NGAL), another kind of lipocalin which is strongly expressed in thyroid carcinomas promotes leukocytes recruitment in tumor microenvironment through increased intracellular iron uptake and, consequentky, more chemokines production [156]. Furthermore, NGAL forms a complex with MMP-9 like Lcn2 and increases its stability which is crucial in cancer cell invasion as well as response to chemotherapy [157].
Apart from Lcn2, Hepcidin is another important factor in the intricate relationship between iron metabolism and tumor microenvironment. The core axis of hepcidin control is the BMP-HJV-SMAD signaling pathway and makes it the major target for pharmacologic intervene in cancer [158,159]. Activation of JAK/STAT3 signaling by inflammatory stimuli IL-6 can also enhance transcriptional activity of hepcidin gene (HAMP) [160]. Tumor-associated fibroblasts induce hepcidin expression via paracrine IL-6-BMP signaling, and this induction facilitates breast cancer cells growth [161]. Whereas the immunophilin FKBP12 represses hepcidin expression by binding the BMP type I receptor ALK2 and blocking BMP-SMAD pathway in hepatoma cells [162]. These findings may pave the way for using hepcidin targeting as a novel treatment for iron homeostasis in tumor tissue and the tumor microenvironment.
5. Iron Manipulating Strategies in Cancer
5.1. Iron is a Target for Oncotherapy
Both iron and ROS are cautiously managed to maintain balance or to stabilize their functions, thus a potential kind of cancer therapy is focused on disrupting the redox homeostasis by introducing or eliminating iron in the cell. Iron overload presented as ferric ammonium citrate (FAC) or iron complexes, remarkably inhibits cell survival in various cancer types [5,163,164]. Previous studies have shown that FAC induces cytoplasmic vacuolation formation in an ATG5/ATG7-dependent manner, with elevated LC3-II (autophagic marker) [159]. The synthetic Fe2+-polypyridyl complexes are able to inhibit glioblastoma tumor growth, and significantly induce TRAIL-mediated cell apoptosis by stimulating p38 and p53 and suppressing ERK pathway [165]. Furthermore, the iron complexes significantly inhibit tumor growth in vivo through enhanced cell apoptosis without evident systematic toxicities as confirmed by histological and pathological analysis [165,166].
On the other hand, iron chelators that deplete cellular iron level by binding iron with high affinity have been shown to suppress the proliferation of aggressive tumors like neuroblastoma and breast cancer, and lead to G1/S cell cycle arrest and apoptosis, suggesting that iron-deprivation may be an promising therapeutic strategy [167,168]. A number of studies have revealed that iron chelation can affect the AKT, ERK, p38, TGF-β, STAT3, JNK, Wnt signaling, and autophagic pathways to consequently suppress tumor growth and metastasis [137,169]. In clinical use, there are three drugs—deferasirox (DFX), deferiprone (DFP), and deferoxamine (DFO)—being evaluated for cancer treatment. DFX may selectively target the NF-κB pathway and induce highly specific apoptosis in myeloid leukemia, hepatoma and mantle cell lymphoma cells [170]. DFP has been reported to inhibit prostate cancer cell and TNBC cells proliferation and migration by decreasing oxygen consumption rate (OCR) and impairing mitochondrial function [171,172]. Megadose of DFO treatment disturbs intracellular iron homeostasis, induces apoptosis and represses growth in breast cancer cell lines [173]. Moreover, it has been suggested that iron chelation could impair not only tumor cells but also tumor microenvironment by affecting the polarized state of TAMs [174]. Taken together, high-dose iron chelators treatment instantaneously decreases LIP content to a level low enough to (1) cause remarkable disorder in cellular iron homeostasis; (2) induce DNA damage, cell cycle arrest and apoptosis; (3) modulate global histone methylation; (4) remodel cancer microenvironment; (5) inhibit cell growth and proliferation in both nonmetastatic and metastatic tumors.
Confronted with the efficacy of iron chelators as cancer therapy, a considerable body preclinical and clinical evidence points out that iron chelators also have nonnegligible toxic side-effects [175]. For example, iron depletion by DFO activates HIF-1α pathway and induces uPA and MMP-2 expression, which results in enhanced metastasis by degrading the extracellular matrix [176]. DFO treatment in the clinical trials has been reported to be able to increase the potent angiogenic factor VEGF level, leading to toxicities such as anemia and edema [177,178,179]. Therefore, novel types of iron chelators with fewer side-effects need to be discovered and more clinical evidences need to be confirmed and resolved. Further work is urgently required to find new iron chelators with maximized antitumor activity against a wide range of iron-overloaded cancer types, improve the oral activity, optimize the therapeutic schedule and clarify safety matters relating to iron deficiency anemia, panleukopenia, and edema. Therefore, the challenge is to design novel iron-scavenging agents those selectively kill cancer cells whilst leaving normal cells unaffected.
5.2. Combination Therapies and Novel Iron Modulators
Though the anti-tumor effect of iron chelators is very limited, iron deprivation in combination with chemotherapy has been certificated to improve each effectiveness without raising toxicity. DFX synergizes with standard chemotherapeutic agents such as doxorubicin, cisplatin, and carboplatin to suppress cell growth and cause apoptosis and autophagy in TNBC cells and other cancer types [180,181]. Moreover, in breast cancer patient-derived xenograft models, it has been reported that tumor recurrences are delayed by the combination without enhancing the side-effects of chemotherapies or impairing systematic iron homeostasis of the mice [182,183]. DFX can also synergistically repress pancreatic cancer cell proliferation with gemcitabine, a standard chemotherapy for pancreatic cancer, in vitro and in vivo [184].
Other than iron chelator combination strategy, iron and iron-based materials have been extensively studied for drug targeting and diagnostic applications [185,186]. Researchers have paid particular attention to iron oxide nanoparticles (IONP) because of their superparamagnetic properties which can be used for diagnosis and treatment. These nanoparticles engender cytotoxicity and genotoxicity in cancer cells via increasing ROS generation, oxidative stress, DNA damage, chromosomal condensation, and caspase-3 activity [187,188,189]. Recent results indicate that the photothermal effect of IONP can cause autophagy of cancerous cells in a laser dose-dependent manner. Co-treatment of IONP and autophagy inhibitor under laser exposure may suppress the xenograft tumor growth and facilitate LC3 production and TUNEL signaling, demonstrating a promising combination therapeutic method of INOP agents and autophagy modulators [190,191]. Moreover, IONP can function as optimal delivery intermediaries. Superparamagnetic iron oxide (SPIO) provide a potential siRNA delivery system, which accumulate easily in orthotopic tumor tissues, and avoid serum nuclease degradation [192]. Modified PEI-SPIO with the carrier Gal, which has a specific receptor on HCC cells, can target the c-Met siRNA specifically to tumor sites and effectively suppress tumor growth in a rat orthotopic model [192]. Taking this a step further, IONP is also applied to mediate the antibody-dependent cell-mediated cytotoxicity (ADCC). Multiple half chains of trastuzumab are conjugated onto magnetic iron oxide nanoparticles (MNP-HC) to form novel biological-active systems to improve target specificity and anti-HER2 therapeutic potential [193]. A novel combined use of IONP (superparamagnetic Fe3O4 nanoparticles) as a vaccine delivery platform and immune potentiator greatly promotes immune cells activation and cytokines production, stimulating potent cellular immune responses, and antigen specific CTL responses [188,194]. Macrophages and DC cells, functioning as the most professional antigen presenting cells, can be activated by Fe3O4-OVA nanoparticles and release diverse pro-inflammatory cytokines—including IL-6, interferon (IFN)-γ, and TNF-α in vitro—thus causing more potent immune responses [195,196]. Another evidence shows that IONP can strongly inhibit the biosynthesis metabolism of macrophages [197]. Furthermore, in recent years, the application of IONPs to visualize cell migration with magnetic resonance imaging (MRI) has been used clinically, showing its value in medical diagnosis [198,199]. All these attempts open a new therapeutic opportunity of iron modulators in different types of cancer and especially in drug-resistant tumors.
6. Conclusions
As far as we know, iron diversely functions in tumor initiation, progression, metastasis, and microenvironment. A large amount of iron in cancer cells is needed for the proliferation and progress. The expressions of numerous iron metabolism-related proteins are aberrantly regulated in malignant tumors, and a variety of signaling pathways and physiological processes are altered by iron in cancer, manifesting the essential roles of iron in cancer development. Overall, aberrant iron homeostasis is to some extent a hallmark of cancer. Thus, strategies based on remodeling iron homeostasis should offer promising choices for cancer therapy. Apart from diminishing intracellular iron content by iron chelators, targeting iron-associated proteins for drug delivery or impairing the redox status by elevating intracellular iron level have all been confirmed as feasible ways for cancer treatment. Despite amounts of iron-related signaling in cancer development have been illustrated in the existing studies, detailed understandings on the mechanisms of iron homeostasis maintenance, iron-associated proteins functions, and global effect on epigenetics and microenvironment involved of iron require further exploration. Last but not least, the efficiency and safety of the strategies based on iron metabolism regulation for cancer therapy still need more efforts to be improved.
Funding
This review is funded by the National Natural Science Foundation of China (81773763, 81521005 and 81803557).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A Red Carpet for Iron Metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [Green Version]
- Recalcati, S.; Gammella, E.; Buratti, P.; Cairo, G. Molecular regulation of cellular iron balance. IUBMB Life 2017, 69, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Torti, S.V.; Torti, F.M. Iron and cancer: More ore to be mined. Nat. Rev. Cancer 2013, 13, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Martinkova, P.; Brtnicky, M.; Kynicky, J.; Pohanka, M. Iron Oxide Nanoparticles: Innovative Tool in Cancer Diagnosis and Therapy. Adv. Healthc. Mater. 2018, 7. [Google Scholar] [CrossRef]
- Viktorinova, A. Iron-mediated oxidative cell death is a potential contributor to neuronal dysfunction induced by neonatal hemolytic hyperbilirubinemia. Arch. Biochem. Biophys. 2018, 654, 185–193. [Google Scholar] [CrossRef]
- Hentze, M.W.; Muckenthaler, M.U.; Galy, B.; Camaschella, C. Two to tango: Regulation of Mammalian iron metabolism. Cell 2010, 142, 24–38. [Google Scholar] [CrossRef]
- Daniels, T.R.; Delgado, T.; Helguera, G.; Penichet, M.L. The transferrin receptor part II: Targeted delivery of therapeutic agents into cancer cells. Clin. Immunol. 2006, 121, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, J.; Song, F.; Tian, M.; Shi, B.; Jiang, H.; Xu, W.; Wang, H.; Zhou, M.; Pan, X.; et al. EGFR regulates iron homeostasis to promote cancer growth through redistribution of transferrin receptor 1. Cancer Lett. 2016, 381, 331–340. [Google Scholar] [CrossRef]
- Bourseau-Guilmain, E.; Griveau, A.; Benoit, J.P.; Garcion, E. The importance of the stem cell marker prominin-1/CD133 in the uptake of transferrin and in iron metabolism in human colon cancer CaCO2 cells. PLoS ONE 2011, 6, e25515. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, B.; Zhang, L.; Wang, S.; Dong, D.; Lv, H.; Shang, P. Alterations in Cellular Iron Metabolism Provide More Therapeutic Opportunities for Cancer. Int. J. Mol. Sci. 2018, 19, 1545. [Google Scholar] [CrossRef]
- Crielaard, B.J.; Lammers, T.; Rivella, S. Targeting iron metabolism in drug discovery and delivery. Nat. Rev. Drug Discov. 2017, 16, 400–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.Y.; Jiang, J.N.; Fang, X.D.; Ji, F.J. STEAP1 Regulates Tumorigenesis and Chemoresistance During Peritoneal Metastasis of Gastric Cancer. Front. Physiol. 2018, 9, 1132. [Google Scholar] [CrossRef]
- Burnell, S.E.A.; Spencer-Harty, S.; Howarth, S.; Bodger, O.; Kynaston, H.; Morgan, C.; Doak, S.H. STEAP2 Knockdown Reduces the Invasive Potential of Prostate Cancer Cells. Sci. Rep. 2018, 8, 6252. [Google Scholar] [CrossRef] [PubMed]
- Whiteland, H.; Spencer-Harty, S.; Morgan, C.; Kynaston, H.; Thomas, D.H.; Bose, P.; Fenn, N.; Lewis, P.; Jenkins, S.; Doak, S.H. A role for STEAP2 in prostate cancer progression. Clin. Exp. Metastasis 2014, 31, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Xu, R.; Wang, S.; Yang, N.; Ni, S.; Zhang, Q.; Xu, Y.; Zhang, X.; Zhang, C.; Wei, Y.; et al. Six-Transmembrane Epithelial Antigen of Prostate 3 Predicts Poor Prognosis and Promotes Glioblastoma Growth and Invasion. Neoplasia 2018, 20, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Bredell, B.X.; Anderson, E.R.; Martin, A.; Mays, C.; Nagao-Kitamoto, H.; Huang, S.; Gyorffy, B.; Greenson, J.K.; Hardiman, K.; et al. Quantitative proteomics identifies STEAP4 as a critical regulator of mitochondrial dysfunction linking inflammation and colon cancer. Proc. Natl. Acad. Sci. USA 2017, 114, E9608–E9617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, I.M.; Maia, C.J.; Santos, C.R. STEAP proteins: From structure to applications in cancer therapy. Mol. Cancer Res. 2012, 10, 573–587. [Google Scholar] [CrossRef]
- Barroca-Ferreira, J.; Pais, J.P.; Santos, M.M.; Goncalves, A.M.; Gomes, I.M.; Sousa, I.; Rocha, S.M.; Passarinha, L.A.; Maia, C.J. Targeting STEAP1 Protein in Human Cancer: Current Trends and Future Challenges. Curr. Cancer Drug Targets 2018, 18, 222–230. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, F. Iron homeostasis and tumorigenesis: Molecular mechanisms and therapeutic opportunities. Protein Cell 2015, 6, 88–100. [Google Scholar] [CrossRef]
- Brookes, M.J.; Hughes, S.; Turner, F.E.; Reynolds, G.; Sharma, N.; Ismail, T.; Berx, G.; McKie, A.T.; Hotchin, N.; Anderson, G.J.; et al. Modulation of iron transport proteins in human colorectal carcinogenesis. Gut 2006, 55, 1449–1460. [Google Scholar] [CrossRef] [Green Version]
- Wolff, N.A.; Garrick, M.D.; Zhao, L.; Garrick, L.M.; Ghio, A.J.; Thevenod, F. A role for divalent metal transporter (DMT1) in mitochondrial uptake of iron and manganese. Sci. Rep. 2018, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, X.; Ramakrishnan, S.K.; Weisz, K.; Triner, D.; Xie, L.; Attili, D.; Pant, A.; Gyorffy, B.; Zhan, M.; Carter-Su, C.; et al. Iron Uptake via DMT1 Integrates Cell Cycle with JAK-STAT3 Signaling to Promote Colorectal Tumorigenesis. Cell Metab. 2016, 24, 447–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, Z.; Hann, H.W.; Ye, Z.; Yin, C.; Wang, Y.; Fang, W.; Wan, S.; Wang, C.; Tao, K. Ferritin level prospectively predicts hepatocarcinogenesis in patients with chronic hepatitis B virus infection. Oncol. Lett. 2018, 16, 3499–3508. [Google Scholar] [CrossRef] [PubMed]
- Song, A.; Eo, W.; Kim, S.; Shim, B.; Lee, S. Significance of serum ferritin as a prognostic factor in advanced hepatobiliary cancer patients treated with Korean medicine: A retrospective cohort study. BMC Complement. Altern. Med. 2018, 18, 176. [Google Scholar] [CrossRef] [PubMed]
- Lipper, C.H.; Karmi, O.; Sohn, Y.S.; Darash-Yahana, M.; Lammert, H.; Song, L.; Liu, A.; Mittler, R.; Nechushtai, R.; Onuchic, J.N.; et al. Structure of the human monomeric NEET protein MiNT and its role in regulating iron and reactive oxygen species in cancer cells. Proc. Natl. Acad. Sci. USA 2018, 115, 272–277. [Google Scholar] [CrossRef]
- Mittler, R.; Darash-Yahana, M.; Sohn, Y.S.; Bai, F.; Song, L.; Cabantchik, I.Z.; Jennings, P.A.; Onuchic, J.N.; Nechushtai, R. NEET Proteins: A New Link Between Iron Metabolism, Reactive Oxygen Species, and Cancer. Antioxid. Redox Signal. 2018. [Google Scholar] [CrossRef]
- Gu, Z.; Wang, H.; Xia, J.; Yang, Y.; Jin, Z.; Xu, H.; Shi, J.; De Domenico, I.; Tricot, G.; Zhan, F. Decreased ferroportin promotes myeloma cell growth and osteoclast differentiation. Cancer Res. 2015, 75, 2211–2221. [Google Scholar] [CrossRef]
- Xue, D.; Zhou, C.X.; Shi, Y.B.; Lu, H.; He, X.Z. Decreased expression of ferroportin in prostate cancer. Oncol. Lett. 2015, 10, 913–916. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Zhang, S.; Chen, Y.; Zhang, D.; Yuan, L.; Cong, H.; Liu, S. An important role of the hepcidin-ferroportin signaling in affecting tumor growth and metastasis. Acta Biochim. Biophys. Sin. 2015, 47, 703–715. [Google Scholar] [CrossRef]
- Moussa, R.S.; Park, K.C.; Kovacevic, Z.; Richardson, D.R. Ironing out the role of the cyclin-dependent kinase inhibitor, p21 in cancer: Novel iron chelating agents to target p21 expression and activity. Free Radic. Boil. Med. 2018. [Google Scholar] [CrossRef]
- Shan, Z.; Wei, Z.; Shaikh, Z.A. Suppression of ferroportin expression by cadmium stimulates proliferation, EMT, and migration in triple-negative breast cancer cells. Toxicol. Appl. Pharmacol. 2018, 356, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, C.N.; Ruwe, T.A.; Shawki, A.; Xin, V.; Vieth, K.R.; Valore, E.V.; Qiao, B.; Ganz, T.; Nemeth, E.; Mackenzie, B.; et al. Calcium is an essential cofactor for metal efflux by the ferroportin transporter family. Nat. Commun. 2018, 9, 3075. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.; Galy, B.; Schwanhaeusser, B.; Blake, J.; Bahr-Ivacevic, T.; Benes, V.; Selbach, M.; Muckenthaler, M.U.; Hentze, M.W. Iron regulatory protein-1 and -2: Transcriptome-wide definition of binding mRNAs and shaping of the cellular proteome by iron regulatory proteins. Blood 2011, 118, e168–e179. [Google Scholar] [CrossRef] [PubMed]
- Kwok, J.C.; Richardson, D.R. The iron metabolism of neoplastic cells: Alterations that facilitate proliferation? Crit. Rev. Oncol. Hematol. 2002, 42, 65–78. [Google Scholar] [CrossRef]
- Horniblow, R.D.; Bedford, M.; Hollingworth, R.; Evans, S.; Sutton, E.; Lal, N.; Beggs, A.; Iqbal, T.H.; Tselepis, C. BRAF mutations are associated with increased iron regulatory protein-2 expression in colorectal tumorigenesis. Cancer Sci. 2017, 108, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Holmes-Hampton, G.P.; Ghosh, M.C.; Rouault, T.A. Methods for Studying Iron Regulatory Protein 1: An Important Protein in Human Iron Metabolism. Methods Enzym. 2018, 599, 139–155. [Google Scholar]
- Wilkinson, N.; Pantopoulos, K. The IRP/IRE system in vivo: Insights from mouse models. Front. Pharmacol. 2014, 5, 176. [Google Scholar] [CrossRef] [PubMed]
- Pantopoulos, K. Iron metabolism and the IRE/IRP regulatory system: An update. Ann. N. Y. Acad. Sci. 2004, 1012, 1–13. [Google Scholar] [CrossRef]
- Du, F.; Qian, Z.M.; Gong, Q.; Zhu, Z.J.; Lu, L.; Ke, Y. The iron regulatory hormone hepcidin inhibits expression of iron release as well as iron uptake proteins in J774 cells. J. Nutr. Biochem. 2012, 23, 1694–1700. [Google Scholar] [CrossRef]
- Yang, J.; Xu, T.; Gomez, D.R.; Yuan, X.; Nguyen, Q.N.; Jeter, M.; Song, Y.; Komaki, R.; Hu, Y.; Hahn, S.M.; et al. Nomograms incorporating genetic variants in BMP/Smad4/Hamp pathway to predict disease outcomes after definitive radiotherapy for non-small cell lung cancer. Cancer Med. 2018, 7, 2247–2255. [Google Scholar] [CrossRef]
- Vela, D.; Vela-Gaxha, Z. Differential regulation of hepcidin in cancer and non-cancer tissues and its clinical implications. Exp. Mol. Med. 2018, 50, e436. [Google Scholar] [CrossRef] [Green Version]
- Huang, X. Iron overload and its association with cancer risk in humans: Evidence for iron as a carcinogenic metal. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2003, 533, 153–171. [Google Scholar] [CrossRef]
- Cairo, G.; Recalcati, S. Iron-regulatory proteins: Molecular biology and pathophysiological implications. Expert Rev. Mol. Med. 2007, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kabat, G.C.; Rohan, T.E. Does excess iron play a role in breast carcinogenesis? An unresolved hypothesis. Cancer Causes Control 2007, 18, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.D.; Coffman, L.G.; Chou, J.W.; Black, M.A.; Bergh, J.; D’Agostino, R., Jr.; Torti, S.V.; Torti, F.M. An iron regulatory gene signature predicts outcome in breast cancer. Cancer Res. 2011, 71, 6728–6737. [Google Scholar] [CrossRef] [PubMed]
- Galaris, D.; Skiada, V.; Barbouti, A. Redox signaling and cancer: The role of “labile” iron. Cancer Lett. 2008, 266, 21–29. [Google Scholar] [CrossRef]
- Tong, W.H.; Maio, N.; Zhang, D.L.; Palmieri, E.M.; Ollivierre, H.; Ghosh, M.C.; McVicar, D.W.; Rouault, T.A. TLR-activated repression of Fe-S cluster biogenesis drives a metabolic shift and alters histone and tubulin acetylation. Blood Adv. 2018, 2, 1146–1156. [Google Scholar] [CrossRef] [Green Version]
- Buzas, D.M.; Nakamura, M.; Kinoshita, T. Epigenetic role for the conserved Fe-S cluster biogenesis protein AtDRE2 in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2014, 111, 13565–13570. [Google Scholar] [CrossRef] [Green Version]
- Sturm, A.; Ivics, Z.; Vellai, T. The mechanism of ageing: Primary role of transposable elements in genome disintegration. Cell. Mol. Life Sci. 2015, 72, 1839–1847. [Google Scholar] [CrossRef]
- Sfera, A.; Bullock, K.; Price, A.; Inderias, L.; Osorio, C. Ferrosenescence: The iron age of neurodegeneration? Mech. Ageing Dev. 2018, 174, 63–75. [Google Scholar] [CrossRef]
- Cao, L.L.; Liu, H.; Yue, Z.; Liu, L.; Pei, L.; Gu, J.; Wang, H.; Jia, M. Iron chelation inhibits cancer cell growth and modulates global histone methylation status in colorectal cancer. Biometals 2018, 31, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Rao, A. Connections between TET proteins and aberrant DNA modification in cancer. Trends Genet. 2014, 30, 464–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascella, B.; Mirica, L.M. Kinetic analysis of iron-dependent histone demethylases: Alpha-ketoglutarate substrate inhibition and potential relevance to the regulation of histone demethylation in cancer cells. Biochemistry 2012, 51, 8699–8701. [Google Scholar] [CrossRef] [PubMed]
- Clifton, I.J.; McDonough, M.A.; Ehrismann, D.; Kershaw, N.J.; Granatino, N.; Schofield, C.J. Structural studies on 2-oxoglutarate oxygenases and related double-stranded beta-helix fold proteins. J. Inorg. Biochem. 2006, 100, 644–669. [Google Scholar] [CrossRef]
- Wilson, S.; Fan, L.; Sahgal, N.; Qi, J.; Filipp, F.V. The histone demethylase KDM3A regulates the transcriptional program of the androgen receptor in prostate cancer cells. Oncotarget 2017, 8, 30328–30343. [Google Scholar] [CrossRef] [PubMed]
- Kuo, K.T.; Huang, W.C.; Bamodu, O.A.; Lee, W.H.; Wang, C.H.; Hsiao, M.; Wang, L.S.; Yeh, C.T. Histone demethylase JARID1B/KDM5B promotes aggressiveness of non-small cell lung cancer and serves as a good prognostic predictor. Clin. Epigenet. 2018, 10, 107. [Google Scholar] [CrossRef]
- Harmeyer, K.M.; Facompre, N.D.; Herlyn, M.; Basu, D. JARID1 Histone Demethylases: Emerging Targets in Cancer. Trends Cancer 2017, 3, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Mao, Y.; Du, G.; He, C.; Han, S. Overexpression of JARID1B is associated with poor prognosis and chemotherapy resistance in epithelial ovarian cancer. Tumour Boil. 2015, 36, 2465–2472. [Google Scholar] [CrossRef] [Green Version]
- Ponnaluri, V.K.; Maciejewski, J.P.; Mukherji, M. A mechanistic overview of TET-mediated 5-methylcytosine oxidation. Biochem. Biophys. Res. Commun. 2013, 436, 115–120. [Google Scholar] [CrossRef]
- Tamanaha, E.; Guan, S.; Marks, K.; Saleh, L. Distributive Processing by the Iron(II)/alpha-Ketoglutarate-Dependent Catalytic Domains of the TET Enzymes Is Consistent with Epigenetic Roles for Oxidized 5-Methylcytosine Bases. J. Am. Chem. Soc. 2016, 138, 9345–9348. [Google Scholar] [CrossRef]
- Zhao, B.; Yang, Y.; Wang, X.; Chong, Z.; Yin, R.; Song, S.H.; Zhao, C.; Li, C.; Huang, H.; Sun, B.F.; et al. Redox-active quinones induces genome-wide DNA methylation changes by an iron-mediated and Tet-dependent mechanism. Nucleic Acids Res. 2014, 42, 1593–1605. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; DesMarais, T.L.; Tong, Z.; Yao, Y.; Costa, M. Oxidative stress alters global histone modification and DNA methylation. Free Radic. Boil. Med. 2015, 82, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udali, S.; Guarini, P.; Ruzzenente, A.; Ferrarini, A.; Guglielmi, A.; Lotto, V.; Tononi, P.; Pattini, P.; Moruzzi, S.; Campagnaro, T.; et al. DNA methylation and gene expression profiles show novel regulatory pathways in hepatocellular carcinoma. Clin. Epigenet. 2015, 7, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Wu, Q.; Monga, J.; Xie, E.; Wang, H.; Wang, S.; Zhang, H.; Wang, Z.Y.; Zhou, T.; Shi, Y.; et al. HDAC1 Governs Iron Homeostasis Independent of Histone Deacetylation in Iron-Overload Murine Models. Antioxid. Redox Signal. 2018, 28, 1224–1237. [Google Scholar] [CrossRef] [PubMed]
- Pasricha, S.R.; Lim, P.J.; Duarte, T.L.; Casu, C.; Oosterhuis, D.; Mleczko-Sanecka, K.; Suciu, M.; Da Silva, A.R.; Al-Hourani, K.; Arezes, J.; et al. Hepcidin is regulated by promoter-associated histone acetylation and HDAC3. Nat. Commun. 2017, 8, 403. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Chen, J.; Feng, J.; Wang, J. E4BP4 promotes thyroid cancer proliferation by modulating iron homeostasis through repression of hepcidin. Cell Death Dis. 2018, 9, 987. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Wu, Q.; Guo, X.; Zhang, Z.; Shen, Y.; Wang, F. MBD5 regulates iron metabolism via methylation-independent genomic targeting of Fth1 through KAT2A in mice. Br. J. Haematol. 2014, 166, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Ohba, C.; Nabatame, S.; Iijima, Y.; Nishiyama, K.; Tsurusaki, Y.; Nakashima, M.; Miyake, N.; Tanaka, F.; Ozono, K.; Saitsu, H.; et al. De novo WDR45 mutation in a patient showing clinically Rett syndrome with childhood iron deposition in brain. J. Hum. Genet. 2014, 59, 292–295. [Google Scholar] [CrossRef]
- Wang, Y.F.; Zhang, J.; Su, Y.; Shen, Y.Y.; Jiang, D.X.; Hou, Y.Y.; Geng, M.Y.; Ding, J.; Chen, Y. G9a regulates breast cancer growth by modulating iron homeostasis through the repression of ferroxidase hephaestin. Nat. Commun. 2017, 8, 274. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.M.; Lee, J.; Finley, L.W.; Schmidt, P.J.; Fleming, M.D.; Haigis, M.C. SIRT3 regulates cellular iron metabolism and cancer growth by repressing iron regulatory protein 1. Oncogene 2015, 34, 2115–2124. [Google Scholar] [CrossRef]
- Guaraldo, M.; Santambrogio, P.; Rovelli, E.; Di Savino, A.; Saglio, G.; Cittaro, D.; Roetto, A.; Levi, S. Characterization of human mitochondrial ferritin promoter: Identification of transcription factors and evidences of epigenetic control. Sci. Rep. 2016, 6, 33432. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, S.; Wang, X.; Guo, W.; Wang, L.; Zhang, D.; Yuan, L.; Zhang, Z.; Xu, Y.; Liu, S. Disordered signaling governing ferroportin transcription favors breast cancer growth. Cell Signal. 2015, 27, 168–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Fujiwara, T.; Ota, U.; Hatta, S.; Ichikawa, S.; Kobayashi, M.; Okitsu, Y.; Fukuhara, N.; Onishi, Y.; Ishizuka, M.; et al. Dynamics of absorption, metabolism, and excretion of 5-aminolevulinic acid in human intestinal Caco-2 cells. Biochem. Biophys. Rep. 2017, 11, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Park, S.H.; Chang, H.C.; Shapiro, J.S.; Vassilopoulos, A.; Sawicki, K.T.; Chen, C.; Shang, M.; Burridge, P.W.; Epting, C.L.; et al. Sirtuin 2 regulates cellular iron homeostasis via deacetylation of transcription factor NRF2. J. Clin. Investig. 2017, 127, 1505–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Kato, H.; Hada, H.; Itoh-Nakadai, A.; Fujiwara, T.; Muto, A.; Inoguchi, Y.; Ichiyanagi, K.; Hojo, W.; Tomosugi, N.; et al. Iron-heme-Bach1 axis is involved in erythroblast adaptation to iron deficiency. Haematologica 2017, 102, 454–465. [Google Scholar] [CrossRef] [PubMed]
- Schaar, D.G.; Medina, D.J.; Moore, D.F.; Strair, R.K.; Ting, Y. miR-320 targets transferrin receptor 1 (CD71) and inhibits cell proliferation. Exp. Hematol. 2009, 37, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, M.; Bogdan, A.R.; Hashimoto, K.; Tsuji, Y. Regulation of transferrin receptor-1 mRNA by the interplay between IRE-binding proteins and miR-7/miR-141 in the 3’-IRE stem-loops. RNA 2018, 24, 468–479. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Kosaka, N.; Ochiya, T.; Kato, T. Micromanaging Iron Homeostasis: Hypoxia-inducible micro-RNA-210 suppresses iron homeostasis-related proteins. J. Boil. Chem. 2012, 287, 34110–34119. [Google Scholar] [CrossRef]
- Shpyleva, S.; Pogribna, M.; Cozart, C.; Bryant, M.S.; Muskhelishvili, L.; Tryndyak, V.P.; Ross, S.A.; Beland, F.A.; Pogribny, I.P. Interstrain differences in the progression of nonalcoholic steatohepatitis to fibrosis in mice are associated with altered hepatic iron metabolism. J. Nutr. Biochem. 2014, 25, 1235–1242. [Google Scholar] [CrossRef]
- Kindrat, I.; Tryndyak, V.; de Conti, A.; Shpyleva, S.; Mudalige, T.K.; Kobets, T.; Erstenyuk, A.M.; Beland, F.A.; Pogribny, I.P. MicroRNA-152-mediated dysregulation of hepatic transferrin receptor 1 in liver carcinogenesis. Oncotarget 2016, 7, 1276–1287. [Google Scholar] [CrossRef]
- Andolfo, I.; De Falco, L.; Asci, R.; Russo, R.; Colucci, S.; Gorrese, M.; Zollo, M.; Iolascon, A. Regulation of divalent metal transporter 1 (DMT1) non-IRE isoform by the microRNA Let-7d in erythroid cells. Haematologica 2010, 95, 1244–1252. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.J.; Kwok, Z.H.; Chew, X.H.; Zhang, B.; Liu, C.; Soong, T.W.; Yang, H.; Tay, Y. A FTH1 gene:pseudogene:microRNA network regulates tumorigenesis in prostate cancer. Nucleic Acids Res. 2018, 46, 1998–2011. [Google Scholar] [CrossRef] [PubMed]
- Shpyleva, S.I.; Tryndyak, V.P.; Kovalchuk, O.; Starlard-Davenport, A.; Chekhun, V.F.; Beland, F.A.; Pogribny, I.P. Role of ferritin alterations in human breast cancer cells. Breast Cancer Res. Treat. 2011, 126, 63–71. [Google Scholar] [CrossRef] [PubMed]
- White, K.; Lu, Y.; Annis, S.; Hale, A.E.; Chau, B.N.; Dahlman, J.E.; Hemann, C.; Opotowsky, A.R.; Vargas, S.O.; Rosas, I.; et al. Genetic and hypoxic alterations of the microRNA-210-ISCU1/2 axis promote iron-sulfur deficiency and pulmonary hypertension. EMBO Mol. Med. 2015, 7, 695–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castoldi, M.; Vujic Spasic, M.; Altamura, S.; Elmen, J.; Lindow, M.; Kiss, J.; Stolte, J.; Sparla, R.; D’Alessandro, L.A.; Klingmuller, U.; et al. The liver-specific microRNA miR-122 controls systemic iron homeostasis in mice. J. Clin. Investig. 2011, 121, 1386–1396. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Du, X.; Lonnerdal, B. miR-214 regulates lactoferrin expression and pro-apoptotic function in mammary epithelial cells. J. Nutr. 2010, 140, 1552–1556. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Lonnerdal, B. miR-584 mediates post-transcriptional expression of lactoferrin receptor in Caco-2 cells and in mouse small intestine during the perinatal period. Int. J. Biochem. Cell Boil. 2010, 42, 1363–1369. [Google Scholar] [CrossRef]
- Babu, K.R.; Muckenthaler, M.U. miR-20a regulates expression of the iron exporter ferroportin in lung cancer. J. Mol. Med. 2016, 94, 347–359. [Google Scholar] [CrossRef]
- Sangokoya, C.; Doss, J.F.; Chi, J.T. Iron-responsive miR-485-3p regulates cellular iron homeostasis by targeting ferroportin. PLoS Genet. 2013, 9, e1003408. [Google Scholar] [CrossRef]
- Thevenod, F. Iron and Its Role in Cancer Defense: A Double-Edged Sword. Met. Ions Life Sci. 2018, 18. [Google Scholar] [CrossRef]
- Le, N.T.; Richardson, D.R. The role of iron in cell cycle progression and the proliferation of neoplastic cells. Biochim. Biophys. Acta 2002, 1603, 31–46. [Google Scholar] [CrossRef]
- Yu, Y.; Kovacevic, Z.; Richardson, D.R. Tuning cell cycle regulation with an iron key. Cell Cycle 2007, 6, 1982–1994. [Google Scholar] [CrossRef] [PubMed]
- Steegmann-Olmedillas, J.L. The role of iron in tumour cell proliferation. Clin. Transl. Oncol. 2011, 13, 71–76. [Google Scholar] [CrossRef]
- Merlot, A.M.; Kalinowski, D.S.; Richardson, D.R. Novel chelators for cancer treatment: Where are we now? Antioxid. Redox Signal. 2013, 18, 973–1006. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Chou, C.H.; Shun, C.T.; Mao, T.L.; Wen, W.F.; Chen, C.D.; Chen, S.U.; Yang, Y.S.; Ho, H.N. Iron suppresses ovarian granulosa cell proliferation and arrests cell cycle through regulating p38 mitogen-activated protein kinase/p53/p21 pathway. Biol. Reprod. 2017, 97, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Manz, D.H.; Torti, S.V.; Torti, F.M. Iron-responsive element-binding protein 2 plays an essential role in regulating prostate cancer cell growth. Oncotarget 2017, 8, 82231–82243. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Song, J.; Yung, B.C.; Zhou, Z.; Wu, A.; Chen, X. Emerging Strategies of Cancer Therapy Based on Ferroptosis. Adv. Mater. 2018, 30, e1704007. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Lu, B.; Chen, X.B.; Ying, M.D.; He, Q.J.; Cao, J.; Yang, B. The Role of Ferroptosis in Cancer Development and Treatment Response. Front. Pharmacol. 2017, 8, 992. [Google Scholar] [CrossRef]
- Seibt, T.M.; Proneth, B.; Conrad, M. Role of GPX4 in ferroptosis and its pharmacological implication. Free Radic. Boil. Med. 2018. [Google Scholar] [CrossRef]
- Alvarez, S.W.; Sviderskiy, V.O.; Terzi, E.M.; Papagiannakopoulos, T.; Moreira, A.L.; Adams, S.; Sabatini, D.M.; Birsoy, K.; Possemato, R. NFS1 undergoes positive selection in lung tumours and protects cells from ferroptosis. Nature 2017, 551, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Kroemer, G.; Tang, D. The tumor suppressor protein p53 and the ferroptosis network. Free Radic. Boil. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Lee, D.H.; Lee, Y.S.; Jo, M.J.; Jeong, Y.A.; Kwon, W.T.; Choudry, H.A.; Bartlett, D.L.; Lee, Y.J. Molecular crosstalk between ferroptosis and apoptosis: Emerging role of ER stress-induced p53-independent PUMA expression. Oncotarget 2017, 8, 115164–115178. [Google Scholar] [CrossRef] [PubMed]
- El Hout, M.; Dos Santos, L.; Hamai, A.; Mehrpour, M. A promising new approach to cancer therapy: Targeting iron metabolism in cancer stem cells. Semin. Cancer Biol. 2018, 53, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Borgstrom, B.; Stegmayr, J.; Abassi, Y.; Kruszyk, M.; Leffler, H.; Persson, L.; Albinsson, S.; Massoumi, R.; Scheblykin, I.G.; et al. The Molecular Basis for Inhibition of Stemlike Cancer Cells by Salinomycin. ACS Cent. Sci. 2018, 4, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Chen, Z.; Wu, D.; Chen, L. Ferritinophagy/ferroptosis: Iron-related newcomers in human diseases. J. Cell. Physiol. 2018, 233, 9179–9190. [Google Scholar] [CrossRef]
- Ryu, M.S.; Duck, K.A.; Philpott, C.C. Ferritin iron regulators, PCBP1 and NCOA4, respond to cellular iron status in developing red cells. Blood Cells Mol. Dis. 2018, 69, 75–81. [Google Scholar] [CrossRef]
- Huang, T.; Sun, Y.; Li, Y.; Wang, T.; Fu, Y.; Li, C.; Li, C. Growth Inhibition of a Novel Iron Chelator, DpdtC, against Hepatoma Carcinoma Cell Lines Partly Attributed to Ferritinophagy-Mediated Lysosomal ROS Generation. Oxid. Med. Cell. Longev. 2018, 2018, 4928703. [Google Scholar] [CrossRef]
- Wylie, A.; Jones, A.E.; D’Brot, A.; Lu, W.J.; Kurtz, P.; Moran, J.V.; Rakheja, D.; Chen, K.S.; Hammer, R.E.; Comerford, S.A.; et al. p53 genes function to restrain mobile elements. Genes Dev. 2016, 30, 64–77. [Google Scholar] [CrossRef]
- Kaomongkolgit, R.; Cheepsunthorn, P.; Pavasant, P.; Sanchavanakit, N. Iron increases MMP-9 expression through activation of AP-1 via ERK/Akt pathway in human head and neck squamous carcinoma cells. Oral Oncol. 2008, 44, 587–594. [Google Scholar] [CrossRef]
- Shan, L. Bimodal lentiviral vector encoding myc-tagged human ferritin heavy chain and green fluorescent protein (GFP). In Molecular Imaging and Contrast Agent Database (MICAD); National Center for Biotechnology Information (US): Bethesda, MD, USA, 2004. [Google Scholar]
- Lin, R.; Huang, J.; Wang, L.; Li, Y.; Lipowska, M.; Wu, H.; Yang, J.; Mao, H. Bevacizumab and near infrared probe conjugated iron oxide nanoparticles for vascular endothelial growth factor targeted MR and optical imaging. Biomater. Sci. 2018, 6, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, K.; Khare, V.; Evstatiev, R.; Kulnigg-Dabsch, S.; Jambrich, M.; Strobl, H.; Gasche, C. Increased expression of HIF2alpha during iron deficiency-associated megakaryocytic differentiation. J. Thromb. Haemost. 2015, 13, 1113–1127. [Google Scholar] [CrossRef] [PubMed]
- Menezes, S.V.; Sahni, S.; Kovacevic, Z.; Richardson, D.R. Interplay of the iron-regulated metastasis suppressor NDRG1 with epidermal growth factor receptor (EGFR) and oncogenic signaling. J. Boil. Chem. 2017, 292, 12772–12782. [Google Scholar] [CrossRef]
- Sahni, S.; Krishan, S.; Richardson, D.R. NDRG1 as a molecular target to inhibit the epithelial-mesenchymal transition: The case for developing inhibitors of metastasis. Future Med. Chem. 2014, 6, 1241–1244. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.Y.; Xia, R.H.; Han, J.; Sun, B.; Sun, S.Y.; Li, J. The MYB/miR-130a/NDRG2 axis modulates tumor proliferation and metastatic potential in salivary adenoid cystic carcinoma. Cell Death Dis. 2018, 9, 917. [Google Scholar] [CrossRef]
- Yang, C.L.; Zheng, X.L.; Ye, K.; Ge, H.; Sun, Y.N.; Lu, Y.F.; Fan, Q.X. NDRG2 suppresses proliferation, migration, invasion and epithelial-mesenchymal transition of esophageal cancer cells through regulating the AKT/XIAP signaling pathway. Int. J. Biochem. Cell Boil. 2018, 99, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Lei, L.; Hong, L.L.; Ling, Z.Q. Potential role of NDRG2 in reprogramming cancer metabolism and epithelial-to-mesenchymal transition. Histol. Histopathol. 2018, 33, 655–663. [Google Scholar]
- Guo, H.F.; Tsai, C.L.; Terajima, M.; Tan, X.; Banerjee, P.; Miller, M.D.; Liu, X.; Yu, J.; Byemerwa, J.; Alvarado, S.; et al. Pro-metastatic collagen lysyl hydroxylase dimer assemblies stabilized by Fe(2+)-binding. Nat. Commun. 2018, 9, 512. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, B.; Yang, M.; Shi, H.; Gan, W. beta2-microglobulin induces epithelial-mesenchymal transition in human renal proximal tubule epithelial cells in vitro. BMC Nephrol. 2015, 16, 60. [Google Scholar] [CrossRef]
- Song, E.; Ramos, S.V.; Huang, X.; Liu, Y.; Botta, A.; Sung, H.K.; Turnbull, P.C.; Wheeler, M.B.; Berger, T.; Wilson, D.J.; et al. Holo-lipocalin-2-derived siderophores increase mitochondrial ROS and impair oxidative phosphorylation in rat cardiomyocytes. Proc. Natl. Acad. Sci. USA 2018, 115, 1576–1581. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Iron homeostasis in host defence and inflammation. Nat. Rev. Immunol. 2015, 15, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.P.; Elliott, R.L. Decreased Iron in Cancer Cells and Their Microenvironment Improves Cytolysis of Breast Cancer Cells by Natural Killer Cells. Anticancer. Res. 2017, 37, 2297–2305. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.M.; He, X.; Liang, T.; Wu, K.C.; Yan, Y.C.; Lu, L.N.; Yang, G.; Luo, Q.Q.; Yung, W.H.; Ke, Y. Lipopolysaccharides upregulate hepcidin in neuron via microglia and the IL-6/STAT3 signaling pathway. Mol. Neurobiol. 2014, 50, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Bj, R.; Dw, V.P.; Lm, G.; Ac, L.; Ra, G.R.; Ahn, C.; Kl, F.; Jf, H. Plasma ferritin concentration is positively associated with in vivo fatty acid mobilization and insulin resistance in obese women. Exp. Physiol. 2018, 103, 1443–1447. [Google Scholar]
- Marques, O.; Porto, G.; Rema, A.; Faria, F.; Cruz Paula, A.; Gomez-Lazaro, M.; Silva, P.; Martins da Silva, B.; Lopes, C. Local iron homeostasis in the breast ductal carcinoma microenvironment. BMC Cancer 2016, 16, 187. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.N.; Lei, Y.H.; Chang, Y.Z. The regulation of iron metabolism in the mononuclear phagocyte system. Expert Rev. Hematol. 2013, 6, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Arezes, J.; Costa, M.; Vieira, I.; Dias, V.; Kong, X.L.; Fernandes, R.; Vos, M.; Carlsson, A.; Rikers, Y.; Porto, G.; et al. Non-transferrin-bound iron (NTBI) uptake by T lymphocytes: Evidence for the selective acquisition of oligomeric ferric citrate species. PLoS ONE 2013, 8, e79870. [Google Scholar] [CrossRef]
- Daher, R.; Karim, Z. Iron metabolism: State of the art. Transfus Clin. Boil. 2017, 24, 115–119. [Google Scholar] [CrossRef]
- Knutson, M.D. Non-transferrin-bound iron transporters. Free Radic. Boil. Med. 2018. [Google Scholar] [CrossRef]
- Sayadi, A.; Nguyen, A.T.; Bard, F.A.; Bard-Chapeau, E.A. Zip14 expression induced by lipopolysaccharides in macrophages attenuates inflammatory response. Inflamm. Res. 2013, 62, 133–143. [Google Scholar] [CrossRef]
- Pyle, C.J.; Azad, A.K.; Papp, A.C.; Sadee, W.; Knoell, D.L.; Schlesinger, L.S. Elemental Ingredients in the Macrophage Cocktail: Role of ZIP8 in Host Response to Mycobacterium tuberculosis. Int. J. Mol. Sci. 2017, 18, 2375. [Google Scholar] [CrossRef] [PubMed]
- Hvidberg, V.; Maniecki, M.B.; Jacobsen, C.; Hojrup, P.; Moller, H.J.; Moestrup, S.K. Identification of the receptor scavenging hemopexin-heme complexes. Blood 2005, 106, 2572–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, M.J.; Andersen, C.B.; Moestrup, S.K. CD163 binding to haptoglobin-hemoglobin complexes involves a dual-point electrostatic receptor-ligand pairing. J. Boil. Chem. 2013, 288, 18834–18841. [Google Scholar] [CrossRef]
- White, C.; Yuan, X.; Schmidt, P.J.; Bresciani, E.; Samuel, T.K.; Campagna, D.; Hall, C.; Bishop, K.; Calicchio, M.L.; Lapierre, A.; et al. HRG1 is essential for heme transport from the phagolysosome of macrophages during erythrophagocytosis. Cell Metab. 2013, 17, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Mertens, C.; Brune, B. Macrophage iron homeostasis and polarization in the context of cancer. Immunobiology 2015, 220, 295–304. [Google Scholar] [CrossRef]
- Agoro, R.; Taleb, M.; Quesniaux, V.F.J.; Mura, C. Cell iron status influences macrophage polarization. PLoS ONE 2018, 13, e0196921. [Google Scholar] [CrossRef]
- Zhou, Y.; Que, K.T.; Zhang, Z.; Yi, Z.J.; Zhao, P.X.; You, Y.; Gong, J.P.; Liu, Z.J. Iron overloaded polarizes macrophage to proinflammation phenotype through ROS/acetyl-p53 pathway. Cancer Med. 2018, 7, 4012–4022. [Google Scholar] [CrossRef]
- Costa da Silva, M.; Breckwoldt, M.O.; Vinchi, F.; Correia, M.P.; Stojanovic, A.; Thielmann, C.M.; Meister, M.; Muley, T.; Warth, A.; Platten, M.; et al. Iron Induces Anti-tumor Activity in Tumor-Associated Macrophages. Front. Immunol. 2017, 8, 1479. [Google Scholar] [CrossRef] [PubMed]
- Tang, X. Tumor-associated macrophages as potential diagnostic and prognostic biomarkers in breast cancer. Cancer Lett. 2013, 332, 3–10. [Google Scholar] [CrossRef]
- Dignass, A.; Farrag, K.; Stein, J. Limitations of Serum Ferritin in Diagnosing Iron Deficiency in Inflammatory Conditions. Int. J. Chronic Dis. 2018, 2018, 9394060. [Google Scholar] [CrossRef]
- Meng, F.; Zhen, S.; Song, B. HBV-specific CD4+ cytotoxic T cells in hepatocellular carcinoma are less cytolytic toward tumor cells and suppress CD8+ T cell-mediated antitumor immunity. APMIS 2017, 125, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.Q.; De Marchi, T.; Timmermans, A.M.; Beekhof, R.; Trapman-Jansen, A.M.; Foekens, R.; Look, M.P.; van Deurzen, C.H.; Span, P.N.; Sweep, F.C.; et al. Ferritin heavy chain in triple negative breast cancer: A favorable prognostic marker that relates to a cluster of differentiation 8 positive (CD8+) effector T-cell response. Mol. Cell. Proteom. 2014, 13, 1814–1827. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Paragas, N.; Ned, R.M.; Qiu, A.; Viltard, M.; Leete, T.; Drexler, I.R.; Chen, X.; Sanna-Cherchi, S.; Mohammed, F.; et al. Scara5 is a ferritin receptor mediating non-transferrin iron delivery. Dev. Cell 2009, 16, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; He, K.; Liu, P.; Xu, L.X. Iron participated in breast cancer chemoresistance by reinforcing IL-6 paracrine loop. Biochem. Biophys. Res. Commun. 2016, 475, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D. Overview of Lactoferrin as a Natural Immune Modulator. J. Pediatr. 2016, 173, S10–S15. [Google Scholar] [CrossRef]
- Wisgrill, L.; Wessely, I.; Spittler, A.; Forster-Waldl, E.; Berger, A.; Sadeghi, K. Human lactoferrin attenuates the proinflammatory response of neonatal monocyte-derived macrophages. Clin. Exp. Immunol. 2018, 192, 315–324. [Google Scholar] [CrossRef]
- Borkham-Kamphorst, E.; van de Leur, E.; Zimmermann, H.W.; Karlmark, K.R.; Tihaa, L.; Haas, U.; Tacke, F.; Berger, T.; Mak, T.W.; Weiskirchen, R. Protective effects of lipocalin-2 (LCN2) in acute liver injury suggest a novel function in liver homeostasis. Biochim. Biophys. Acta 2013, 1832, 660–673. [Google Scholar] [CrossRef] [Green Version]
- Karlsen, J.R.; Borregaard, N.; Cowland, J.B. Induction of neutrophil gelatinase-associated lipocalin expression by co-stimulation with interleukin-17 and tumor necrosis factor-alpha is controlled by IkappaB-zeta but neither by C/EBP-beta nor C/EBP-delta. J. Boil. Chem. 2010, 285, 14088–14100. [Google Scholar] [CrossRef]
- Johansen, C.; Bertelsen, T.; Ljungberg, C.; Mose, M.; Iversen, L. Characterization of TNF-alpha- and IL-17A-Mediated Synergistic Induction of DEFB4 Gene Expression in Human Keratinocytes through IkappaBzeta. J. Investig. Dermatol. 2016, 136, 1608–1616. [Google Scholar] [CrossRef]
- Moschen, A.R.; Gerner, R.R.; Wang, J.; Klepsch, V.; Adolph, T.E.; Reider, S.J.; Hackl, H.; Pfister, A.; Schilling, J.; Moser, P.L.; et al. Lipocalin 2 Protects from Inflammation and Tumorigenesis Associated with Gut Microbiota Alterations. Cell Host Microbe 2016, 19, 455–469. [Google Scholar] [CrossRef]
- Koh, S.A.; Lee, K.H. HGF mediated upregulation of lipocalin 2 regulates MMP9 through nuclear factor-kappaB activation. Oncol. Rep. 2015, 34, 2179–2187. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; He, K.; Li, J.; Cheng, M.; Song, H.; Liu, J.; Liu, P. Tumor associated macrophages deliver iron to tumor cells via Lcn2. Int. J. Physiol. Pathophysiol. Pharmacol. 2018, 10, 105–114. [Google Scholar] [PubMed]
- Lisowska-Myjak, B.; Skarzynska, E.; Wilczynska, P.; Jakimiuk, A. Correlation between the concentrations of lactoferrin and neutrophil gelatinase-associated lipocalin in meconium. Biometals 2018, 31, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.C.; Chaze, T.; Coic, Y.M.; Injarabian, L.; Jonsson, F.; Lombion, N.; Selimoglu-Buet, D.; Souphron, J.; Ridley, C.; Vonaesch, P.; et al. MUB40 Binds to Lactoferrin and Stands as a Specific Neutrophil Marker. Cell Chem. Boil. 2018, 25, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, F.; Pisa, L.; Mellone, S.; Cillo, M.; Lepore, A.; Leonardi, A. NGAL promotes recruitment of tumor infiltrating leukocytes. Oncotarget 2018, 9, 30761–30772. [Google Scholar] [CrossRef] [PubMed]
- Candido, S.; Abrams, S.L.; Steelman, L.S.; Lertpiriyapong, K.; Fitzgerald, T.L.; Martelli, A.M.; Cocco, L.; Montalto, G.; Cervello, M.; Polesel, J.; et al. Roles of NGAL and MMP-9 in the tumor microenvironment and sensitivity to targeted therapy. Biochim. Biophys. Acta 2016, 1863, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Fillebeen, C.; Wilkinson, N.; Charlebois, E.; Katsarou, A.; Wagner, J.; Pantopoulos, K. Hepcidin-mediated hypoferremic response to acute inflammation requires a threshold of Bmp6/Hjv/Smad signaling. Blood 2018, 132, 1829–1841. [Google Scholar] [CrossRef]
- Arosio, P. New signaling pathways for hepcidin regulation. Blood 2014, 123, 1433–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, E.; Lu, Y.; Yan, M.; Pan, X.; Cheng, X. Increased expression of hepcidin and associated upregulation of JAK/STAT3 signaling in human gastric cancer. Oncol. Lett. 2018, 15, 2236–2244. [Google Scholar] [CrossRef] [PubMed]
- Blanchette-Farra, N.; Kita, D.; Konstorum, A.; Tesfay, L.; Lemler, D.; Hegde, P.; Claffey, K.P.; Torti, F.M.; Torti, S.V. Contribution of three-dimensional architecture and tumor-associated fibroblasts to hepcidin regulation in breast cancer. Oncogene 2018, 37, 4013–4032. [Google Scholar] [CrossRef] [PubMed]
- Colucci, S.; Pagani, A.; Pettinato, M.; Artuso, I.; Nai, A.; Camaschella, C.; Silvestri, L. The immunophilin FKBP12 inhibits hepcidin expression by binding the BMP type I receptor ALK2 in hepatocytes. Blood 2017, 130, 2111–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neshastehriz, A.; Khosravi, Z.; Ghaznavi, H.; Shakeri-Zadeh, A. Gold-coated iron oxide nanoparticles trigger apoptosis in the process of thermo-radiotherapy of U87-MG human glioma cells. Radiat. Environ. Biophys. 2018, 57, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Torti, S.V.; Manz, D.H.; Paul, B.T.; Blanchette-Farra, N.; Torti, F.M. Iron and Cancer. Annu. Rev. Nutr. 2018, 38, 97–125. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Wang, Y.; Lai, H.; Li, X.; Chen, T. Iron(II)-Polypyridyl Complexes Inhibit the Growth of Glioblastoma Tumor and Enhance TRAIL-Induced Cell Apoptosis. Chem. Asian J. 2018, 13, 2730–2738. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, F.; Wei, Z.; Li, X.; Zhao, H.; Lv, H.; Ge, R.; Ma, H.; Zhang, H.; Yang, B.; et al. Magnetic delivery of Fe3O4@polydopamine nanoparticle-loaded natural killer cells suggest a promising anticancer treatment. Biomater. Sci. 2018, 6, 2714–2725. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Zhao, H.Y.; Du, J.; Wang, F. Anti-tumor activities of four chelating agents against human neuroblastoma cells. In Vivo 2005, 19, 233–236. [Google Scholar] [PubMed]
- Li, P.; Zheng, X.; Shou, K.; Niu, Y.; Jian, C.; Zhao, Y.; Yi, W.; Hu, X.; Yu, A. The iron chelator Dp44mT suppresses osteosarcoma's proliferation, invasion and migration: in vitro and in vivo. Am. J. Transl. Res. 2016, 8, 5370–5385. [Google Scholar] [PubMed]
- Lui, G.Y.; Kovacevic, Z.; Richardson, V.; Merlot, A.M.; Kalinowski, D.S.; Richardson, D.R. Targeting cancer by binding iron: Dissecting cellular signaling pathways. Oncotarget 2015, 6, 18748–18779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breccia, M.; Alimena, G. Efficacy and safety of deferasirox in myelodysplastic syndromes. Ann. Hematol. 2013, 92, 863–870. [Google Scholar] [CrossRef]
- Simoes, R.V.; Veeraperumal, S.; Serganova, I.S.; Kruchevsky, N.; Varshavsky, J.; Blasberg, R.G.; Ackerstaff, E.; Koutcher, J.A. Inhibition of prostate cancer proliferation by Deferiprone. NMR Biomed. 2017, 30. [Google Scholar] [CrossRef] [PubMed]
- Knickle, A.; Fernando, W.; Greenshields, A.L.; Rupasinghe, H.P.V.; Hoskin, D.W. Myricetin-induced apoptosis of triple-negative breast cancer cells is mediated by the iron-dependent generation of reactive oxygen species from hydrogen peroxide. Food Chem. Toxicol. 2018, 118, 154–167. [Google Scholar] [CrossRef]
- Bajbouj, K.; Shafarin, J.; Hamad, M. High-Dose Deferoxamine Treatment Disrupts Intracellular Iron Homeostasis, Reduces Growth, and Induces Apoptosis in Metastatic and Nonmetastatic Breast Cancer Cell Lines. Technol. Cancer Res. Treat. 2018, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertens, C.; Akam, E.A.; Rehwald, C.; Brune, B.; Tomat, E.; Jung, M. Intracellular Iron Chelation Modulates the Macrophage Iron Phenotype with Consequences on Tumor Progression. PLoS ONE 2016, 11, e0166164. [Google Scholar] [CrossRef] [PubMed]
- Busti, F.; Marchi, G.; Ugolini, S.; Castagna, A.; Girelli, D. Anemia and Iron Deficiency in Cancer Patients: Role of Iron Replacement Therapy. Pharmaceuticals 2018, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.H.; Huang, P.H.; Hsu, Y.J.; Peng, Y.J.; Lee, C.H.; Wang, J.C.; Chen, J.W.; Lin, S.J. Inhibition of hypoxia inducible factor-1alpha attenuates abdominal aortic aneurysm progression through the down-regulation of matrix metalloproteinases. Sci. Rep. 2016, 6, 28612. [Google Scholar] [CrossRef] [PubMed]
- Kontoghiorghes, G.J. Ethical issues and risk/benefit assessment of iron chelation therapy: Advances with deferiprone/deferoxamine combinations and concerns about the safety, efficacy and costs of deferasirox. Hemoglobin 2008, 32, 1–15. [Google Scholar] [CrossRef]
- Di Nicola, M.; Barteselli, G.; Dell’Arti, L.; Ratiglia, R.; Viola, F. Functional and Structural Abnormalities in Deferoxamine Retinopathy: A Review of the Literature. BioMed Res. Int. 2015, 2015, 249617. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.L.; Hatef, A.; Imran ul-Haq, M.; Nair, N.; Unniappan, S.; Kizhakkedathu, J.N. Clinically approved iron chelators influence zebrafish mortality, hatching morphology and cardiac function. PLoS ONE 2014, 9, e109880. [Google Scholar] [CrossRef]
- Tury, S.; Assayag, F.; Bonin, F.; Chateau-Joubert, S.; Servely, J.L.; Vacher, S.; Becette, V.; Caly, M.; Rapinat, A.; Gentien, D.; et al. The iron chelator deferasirox synergises with chemotherapy to treat triple-negative breast cancers. J. Pathol. 2018, 246, 103–114. [Google Scholar] [CrossRef]
- Piro, E.; Lentini, M.; Levato, L.; Russo, A.; Molica, S. Sustained Erythroid Response in a Patient with Myelofibrosis Receiving Concomitant Treatment with Ruxolitinib and Deferasirox. Chemotherapy 2018, 63, 107–110. [Google Scholar] [CrossRef]
- Chang, Y.C.; Lo, W.J.; Huang, Y.T.; Lin, C.L.; Feng, C.C.; Lin, H.T.; Cheng, H.C.; Yeh, S.P. Deferasirox has strong anti-leukemia activity but may antagonize theanti-leukemia effect of doxorubicin. Leuk. Lymphoma 2017, 58, 1–12. [Google Scholar] [CrossRef]
- Ikeda, R.; Vermeulen, L.C.; Jiang, Z.; Lau, E.; Kolesar, J.M. Gemcitabine and paclitaxel suppress the production of vascular endothelial growth factor induced by deferoxamine in human non-small cell lung cancer A549 cells. Exp. Ther. Med. 2010, 1, 853–857. [Google Scholar] [CrossRef] [Green Version]
- Shinoda, S.; Kaino, S.; Amano, S.; Harima, H.; Matsumoto, T.; Fujisawa, K.; Takami, T.; Yamamoto, N.; Yamasaki, T.; Sakaida, I. Deferasirox, an oral iron chelator, with gemcitabine synergistically inhibits pancreatic cancer cell growth in vitro and in vivo. Oncotarget 2018, 9, 28434–28444. [Google Scholar] [CrossRef] [PubMed]
- Eyvazzadeh, N.; Shakeri-Zadeh, A.; Fekrazad, R.; Amini, E.; Ghaznavi, H.; Kamran Kamrava, S. Gold-coated magnetic nanoparticle as a nanotheranostic agent for magnetic resonance imaging and photothermal therapy of cancer. Lasers Med. Sci. 2017, 32, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Wang, X.; Zhou, S.; Zhang, Y. IONP-doped nanoparticles for highly effective NIR-controlled drug release and combination tumor therapy. Int. J. Nanomed. 2017, 12, 3751–3766. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhou, Z.; Mao, H.; Yang, L. Magnetic nanoparticles for precision oncology: Theranostic magnetic iron oxide nanoparticles for image-guided and targeted cancer therapy. Nanomedicine 2017, 12, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, X.; Cheng, Y.; Guo, X.; Yuan, W. Iron Oxide Nanoparticles-Based Vaccine Delivery for Cancer Treatment. Mol. Pharm. 2018, 15, 1791–1799. [Google Scholar] [CrossRef]
- Alarifi, S.; Ali, D.; Alkahtani, S.; Alhader, M.S. Iron oxide nanoparticles induce oxidative stress, DNA damage, and caspase activation in the human breast cancer cell line. Boil. Trace Element Res. 2014, 159, 416–424. [Google Scholar] [CrossRef]
- Ren, X.; Chen, Y.; Peng, H.; Fang, X.; Zhang, X.; Chen, Q.; Wang, X.; Yang, W.; Sha, X. Blocking Autophagic Flux Enhances Iron Oxide Nanoparticle Photothermal Therapeutic Efficiency in Cancer Treatment. ACS Appl. Mater. Interfaces 2018, 10, 27701–27711. [Google Scholar] [CrossRef]
- Tsai, M.F.; Hsu, C.; Yeh, C.S.; Hsiao, Y.J.; Su, C.H.; Wang, L.F. Tuning the Distance of Rattle-Shaped IONP@Shell-in-Shell Nanoparticles for Magnetically-Targeted Photothermal Therapy in the Second Near-Infrared Window. ACS Appl. Mater. Interfaces 2018, 10, 1508–1519. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Duan, J.; Wang, J.; Liu, Q.; Shang, R.; Yang, X.; Lu, P.; Xia, C.; Wang, L.; Dou, K. Superparamagnetic iron oxide nanoparticles modified with polyethylenimine and galactose for siRNA targeted delivery in hepatocellular carcinoma therapy. Int. J. Nanomed. 2018, 13, 1851–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truffi, M.; Colombo, M.; Sorrentino, L.; Pandolfi, L.; Mazzucchelli, S.; Pappalardo, F.; Pacini, C.; Allevi, R.; Bonizzi, A.; Corsi, F.; et al. Multivalent exposure of trastuzumab on iron oxide nanoparticles improves antitumor potential and reduces resistance in HER2-positive breast cancer cells. Sci. Rep. 2018, 8, 6563. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhanl, D.; Du, Y. The immunotherapeutic effect of Fe3O4 nanoparticles as adjuvants on mice H22 live cancer. J. Nanosci. Nanotechnol. 2010, 10, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Jin, N.; Wang, J.; Chen, B.A.; Ding, J.H.; Gao, C.; Cheng, J.; Zhao, G.; Bao, W.; Gao, F.; Xia, G.H.; et al. Influence of magnetic Fe3O4 nanoparticle on functions of lymphocytes and macrophages in mice. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2010, 18, 136–139. [Google Scholar] [PubMed]
- Jeong, J.; Kwon, E.K.; Cheong, T.C.; Park, H.; Cho, N.H.; Kim, W. Synthesis of multifunctional Fe(3)O(4)-CdSe/ZnS nanoclusters coated with lipid A toward dendritic cell-based immunotherapy. ACS Appl. Mater. Interfaces 2014, 6, 5297–5307. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Z.; Gu, N.; Wang, J. Effects of DMSA-coated Fe3O4 magnetic nanoparticles on global gene expression of mouse macrophage RAW264.7 cells. Toxicol. Lett. 2011, 205, 130–139. [Google Scholar] [CrossRef]
- Jaidev, L.R.; Chellappan, D.R.; Bhavsar, D.V.; Ranganathan, R.; Sivanantham, B.; Subramanian, A.; Sharma, U.; Jagannathan, N.R.; Krishnan, U.M.; Sethuraman, S. Multi-functional nanoparticles as theranostic agents for the treatment & imaging of pancreatic cancer. Acta Biomater. 2017, 49, 422–433. [Google Scholar] [PubMed]
- Zheng, X.C.; Ren, W.; Zhang, S.; Zhong, T.; Duan, X.C.; Yin, Y.F.; Xu, M.Q.; Hao, Y.L.; Li, Z.T.; Li, H.; et al. The theranostic efficiency of tumor-specific, pH-responsive, peptide-modified, liposome-containing paclitaxel and superparamagnetic iron oxide nanoparticles. Int. J. Nanomed. 2018, 13, 1495–1504. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Iron and epigenetic regulation. Iron can modulate heterochromatin assembly mediated by Fenton reactions and induce global histone methylation changes through iron-dependent JmjC-domain-containing epigenetic modifying enzymes in cancer cells. Multiple miRNAs have been demonstrated to regulate iron metabolism-related proteins. DNA methylation, histone acetylation/methylation modification, and some transcription factors such as NRF2 and MZF-1 function corporately to maintain cellular iron metabolism in cancer. TfR, transferrin receptor 1; IRP2, iron regulatory protein; FPN, ferroportin; DMT1, divalentmetal transporter 1; ER, endoplasmic reticulum; HDAC, histone deacetylase; HEPH, hephaestin; Elp3, elongator complex protein 3; 5mC, 5-methylcytosine; TET, ten-eleven translocation protein.
Figure 1.
Iron and epigenetic regulation. Iron can modulate heterochromatin assembly mediated by Fenton reactions and induce global histone methylation changes through iron-dependent JmjC-domain-containing epigenetic modifying enzymes in cancer cells. Multiple miRNAs have been demonstrated to regulate iron metabolism-related proteins. DNA methylation, histone acetylation/methylation modification, and some transcription factors such as NRF2 and MZF-1 function corporately to maintain cellular iron metabolism in cancer. TfR, transferrin receptor 1; IRP2, iron regulatory protein; FPN, ferroportin; DMT1, divalentmetal transporter 1; ER, endoplasmic reticulum; HDAC, histone deacetylase; HEPH, hephaestin; Elp3, elongator complex protein 3; 5mC, 5-methylcytosine; TET, ten-eleven translocation protein.

Figure 2.
Iron handling in the tumor microenvironment. Tumor microenvironment compartments play a critical role in controlling iron metabolism. Inflammatory cytokines upregulate Lcn2 via NF-κB pathway. After releasing out of the cell, Lcn2 sequesters iron and stabilize MMP-9, promoting cell survival and matrix degradation leading to EMT. M2 macrophages are major sites of taking up, metabolizing, storing, and exporting iron. They supply iron to accelerate tumor growth by multiple transport pathways. Tumor-associated fibroblasts contribute to hepcidin induction via paracrine IL-6/BMP/SMAD signaling. Circulating T cells has accumulated H-ferritin to maintain proper immune functions. Th1 cells and NKT cells can secret cytokines like IFN-γ and TNF to the environment, which increase DMT1 whereas decrease FPN level, thus resulting in iron sequestration in the MPS. Tumor-associated fibroblasts induce hepcidin expression via paracrine IL-6-BMP-SMAD signaling. Lcn2, Lipocalin 2; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; MMP-9, matrix metalloproteinases-9; EMT, epithelial-mesenchymal transition; IL-6, interleukin-6; NKT, natural killer T cells; IFN—γ, interferon-γ; TNF, tumor necrosis factor; FPN, ferroportin; MPS, mononuclear phagocyte system.
Figure 2.
Iron handling in the tumor microenvironment. Tumor microenvironment compartments play a critical role in controlling iron metabolism. Inflammatory cytokines upregulate Lcn2 via NF-κB pathway. After releasing out of the cell, Lcn2 sequesters iron and stabilize MMP-9, promoting cell survival and matrix degradation leading to EMT. M2 macrophages are major sites of taking up, metabolizing, storing, and exporting iron. They supply iron to accelerate tumor growth by multiple transport pathways. Tumor-associated fibroblasts contribute to hepcidin induction via paracrine IL-6/BMP/SMAD signaling. Circulating T cells has accumulated H-ferritin to maintain proper immune functions. Th1 cells and NKT cells can secret cytokines like IFN-γ and TNF to the environment, which increase DMT1 whereas decrease FPN level, thus resulting in iron sequestration in the MPS. Tumor-associated fibroblasts induce hepcidin expression via paracrine IL-6-BMP-SMAD signaling. Lcn2, Lipocalin 2; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; MMP-9, matrix metalloproteinases-9; EMT, epithelial-mesenchymal transition; IL-6, interleukin-6; NKT, natural killer T cells; IFN—γ, interferon-γ; TNF, tumor necrosis factor; FPN, ferroportin; MPS, mononuclear phagocyte system.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.; Yu, L.; Ding, J.; Chen, Y. Iron Metabolism in Cancer. Int. J. Mol. Sci. 2019, 20, 95. https://doi.org/10.3390/ijms20010095
AMA Style
Wang Y, Yu L, Ding J, Chen Y. Iron Metabolism in Cancer. International Journal of Molecular Sciences. 2019; 20(1):95. https://doi.org/10.3390/ijms20010095
Chicago/Turabian StyleWang, Yafang, Lei Yu, Jian Ding, and Yi Chen. 2019. "Iron Metabolism in Cancer" International Journal of Molecular Sciences 20, no. 1: 95. https://doi.org/10.3390/ijms20010095
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.