MicroRNAs in Human Placental Development and Pregnancy Complications

Abstract
:1. Introduction
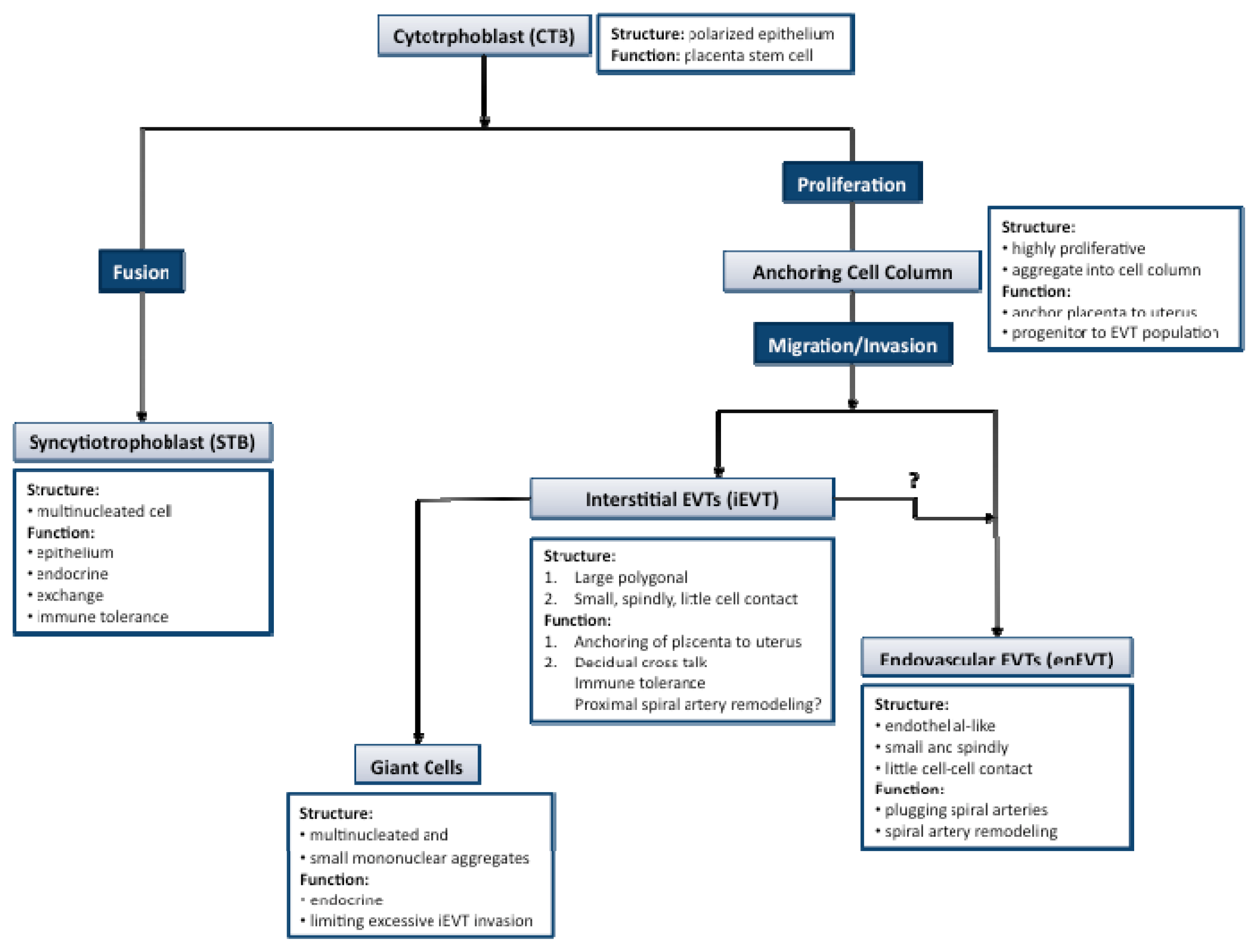
2. Key Processes in Human Placental Development
3. Expression of MiRNAs in Human Placenta
3.1. MicroRNA Biogenesis and Mechanism of Gene Regulation
3.2. Detection and Identification of MiRNAs in Placenta
3.3. MicroRNA Expression during Different Stages of Placental Development
3.4. Regulation of MiRNA Expression in Placenta
4. Regulation of Placental Development and Function by MiRNAs
4.1. MicroRNAs and Trophoblast Cell Proliferation and Apoptosis
4.2. MicroRNAs and Trophoblast Cell Migration and Invasion
4.3. MicroRNAs and Placental Angiogenesis
5. Implication of MiRNAs in Compromised Pregnancies
5.1. Aberrant Expression of MiRNAs in Preeclampsia
5.2. Potential Contribution of MiRNAs to Preeclampsia
5.3. MicroRNAs and Other Gestational Disorders
5.4. Circulating MiRNA as Potential Biomarkers for Pregnancy and Pregnancy-Associated Disorders
6. Summary and Perspectives
Acknowledgments
References
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar]
- Cross, J.C. Formation of the placenta and extraembryonic membranes. Annu. N. Y. Acad. Sci 1998, 857, 23–32. [Google Scholar]
- Murphy, V.E.; Smith, R.; Giles, W.B.; Clifton, V.L. Endocrine regulation of human fetal growth: The role of the mother, placenta, and fetus. Endocr. Rev 2006, 27, 141–169. [Google Scholar]
- Dey, S.K.; Lim, H.; Das, S.K.; Reese, J.; Paria, B.C.; Daikoku, T.; Wang, H. Molecular cues to implantation. Endocr. Rev 2004, 25, 341–373. [Google Scholar]
- Pijnenborg, R. Implantation and immunology: Maternal inflammatory and immune cellular responses to implantation and trophoblast invasion. Reprod. Biomed. Online 2002, 4, 14–17. [Google Scholar]
- Rossant, J.; Cross, J.C. Placental development: Lessons from mouse mutants. Nat. Rev. Genet 2001, 2, 538–548. [Google Scholar]
- Steegers, E.A.; von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Pre-eclampsia. Lancet 2010, 376, 631–644. [Google Scholar]
- James, J.L.; Whitley, G.S.; Cartwright, J.E. Pre-eclampsia: Fitting together the placental, immune and cardiovascular pieces. J. Pathol 2010, 221, 363–378. [Google Scholar]
- Zhong, Y.; Tuuli, M.; Odibo, A.O. First-trimester assessment of placenta function and the prediction of preeclampsia and intrauterine growth restriction. Prenat. Diagn 2010, 30, 293–308. [Google Scholar]
- Mouillet, J.F.; Chu, T.; Hubel, C.A.; Nelson, D.M.; Parks, W.T.; Sadovsky, Y. The levels of hypoxia-regulated microRNAs in plasma of pregnant women with fetal growth restriction. Placenta 2010, 31, 781–784. [Google Scholar]
- Lee, D.C.; Romero, R.; Kim, J.S.; Tarca, A.L.; Montenegro, D.; Pineles, B.L.; Kim, E.; Lee, J.; Kim, S.Y.; Draghici, S.; et al. MiR-210 targets iron-sulfur cluster scaffold homologue in human trophoblast cell lines: Siderosis of interstitial trophoblasts as a novel pathology of preterm preeclampsia and small-for-gestational-age pregnancies. Am. J. Pathol 2011, 179, 590–602. [Google Scholar]
- Saenger, P.; Czernichow, P.; Hughes, I.; Reiter, E.O. Small for gestational age: Short stature and beyond. Endocr. Rev 2007, 28, 219–251. [Google Scholar]
- Mayor-Lynn, K.; Toloubeydokhti, T.; Cruz, A.C.; Chegini, N. Expression profile of microRNAs and mRNAs in human placentas from pregnancies complicated by preeclampsia and preterm labor. Reprod. Sci 2011, 18, 46–56. [Google Scholar]
- Gauster, M.; Desoye, G.; Tötsch, M.; Hiden, U. The placenta and gestational diabetes mellitus. Curr. Diab. Rep 2012, 12, 16–23. [Google Scholar]
- Fu, G.; Ye, G.; Nadeem, L.; Ji, L.; Manchanda, T.; Wang, Y.; Zhao, Y.; Qiao, J.; Wnag, Y.-L.; Lye, S.; et al. MicroRNA-376c impairs transforming growth factor-beta and nodal signaling to promote trophoblast cell proliferation and invasion. Hypertension 2013. [Google Scholar] [CrossRef]
- Mouillet, J.F.; Chu, T.; Sadovsky, Y. Expression patterns of placental microRNAs. Birth Defects Res. A 2011, 91, 737–743. [Google Scholar]
- Enquobahrie, D.A.; Abetew, D.F.; Sorensen, T.K.; Willoughby, D.; Chidambaram, K.; Williams, M.A. Placental microRNA expression in pregnancies complicated by preeclampsia. Am. J. Obstet. Gynecol. 2011, 204, 178.e12–178.e21. [Google Scholar]
- Luo, L.; Ye, G.; Nadeem, L.; Fu, G.; Yang, B.B.; Honarparvar, E.; Dunk, C.; Lye, S.; Peng, C. MicroRNA-378a-5p promotes trophoblast cell survival, migration and invasion by targeting Nodal. J. Cell Sci 2012, 125, 3124–3132. [Google Scholar]
- Bai, Y.; Yang, W.; Yang, H.; Liao, Q.; Ye, G.; Fu, G.; Ji, L.; Xu, P.; Wang, H.; Li, Y.; et al. Downregulated miR-195 detected in preeclamptic placenta affects trophoblast cell invasion via modulating ActRIIA expression. PLoS One 2012, 7, e38875. [Google Scholar]
- Barad, O.; Meiri, E.; Avniel, A.; Aharonov, R.; Barzilai, A.; Bentwich, I.; Einav, U.; Gilad, S.; Hurban, P.; Karov, Y.; et al. MicroRNA expression detected by oligonucleotide microarrays: System establishment and expression profiling in human tissues. Genome Res 2004, 14, 2486–2494. [Google Scholar]
- Donker, R.B.; Mouillet, J.F.; Nelson, D.M.; Sadovsky, Y. The expression of Argonaute2 and related microRNA biogenesis proteins in normal and hypoxic trophoblasts. Mol. Hum. Reprod 2007, 13, 273–279. [Google Scholar]
- Li, P.; Guo, W.; Du, L.; Zhao, J.; Wang, Y.; Liu, L.; Hu, Y.; Hou, Y. MicroRNA-29b contributes to pre-eclampsia through its effects on apoptosis, invasion and angiogenesis of trophoblast cells. Clin. Sci 2013, 124, 27–40. [Google Scholar]
- Dai, Y.; Qiu, Z.; Diao, Z.; Shen, L.; Xue, P.; Sun, H.; Hu, Y. MicroRNA-155 inhibits proliferation and migration of human extravillous trophoblast derived HTR-8/SVneo cells via down-regulating cyclin D1. Placenta 2012, 33, 824–829. [Google Scholar]
- Hu, Y.; Li, P.; Hao, S.; Liu, L.; Zhao, J.; Hou, Y. Differential expression of microRNAs in the placentae of Chinese patients with severe pre-eclampsia. Clin. Chem. Lab. Med 2009, 47, 923–929. [Google Scholar]
- Zhu, X.M.; Han, T.; Sargent, I.L.; Yin, G.W.; Yao, Y.Q. Differential expression profile of microRNAs in human placentas from preeclamptic pregnancies vs normal pregnancies. Am. J. Obstet. Gynecol. 2009, 200, 661.e1–661.e7. [Google Scholar]
- Norwitz, E.R. Defective implantation and placentation: Laying the blueprint for pregnancy complications. Reprod. Biomed. Online 2007, 14, 101–109. [Google Scholar]
- Red-Horse, K.; Zhou, Y.; Genbacev, O.; Prakobphol, A.; Foulk, R.; McMaster, M.; Fisher, S.J. Trophoblast differentiation during embryo implantation and formation of the maternal-fetal interface. J. Clin. Invest 2004, 114, 744–754. [Google Scholar]
- Clancy, K.B. Reproductive ecology and the endometrium: Physiology, variation, and new directions. Am. J. Phys. Anthropol 2009, 140, 137–154. [Google Scholar]
- Anin, S.; Vince, G.; Quenby, S. Trophoblast invasion. Hum. Fert 2004, 7, 169–174. [Google Scholar]
- Knofler, M. Critical growth factors and signalling pathways controlling human trophoblast invasion. Int. J. Dev. Biol 2010, 54, 269–280. [Google Scholar]
- Mayhew, T.M. Estimating oxygen diffusive conductances of gas-exchange systems: A stereological approach illustrated with the human placenta. Ann. Anat. 2012. [Google Scholar] [CrossRef]
- Guibourdenche, J.; Fournier, T.; Malassiné, A.; Evain-Brion, D. Development and hormonal functions of the human placenta. Folia Histochem. Cytobiol 2009, 47, S35–S40. [Google Scholar]
- Nakamura, O. Children’s immunology, what can we learn from animal studies (1): Decidual cells induce specific immune system of feto-maternal interface. J. Toxicol. Sci 2009, 34, SP331–339. [Google Scholar]
- Richart, R. Studies of placental morphogenesis. I. Radioautographic studies of human placenta utilizing tritiated thymidine. Proc. Soc. Exp. Biol. Med 1961, 106, 829–831. [Google Scholar]
- Kar, M.; Ghosh, D.; Sengupta, J. Histochemical and morphological examination of proliferation and apoptosis in human first trimester villous trophoblast. Hum. Reprod 2007, 22, 2814–2823. [Google Scholar]
- Kemp, B.; Kertschanska, S.; Kadyrov, M.; Rath, W.; Kaufmann, P.; Huppertz, B. Invasive depth of extravillous trophoblast correlates with cellular phenotype: A comparison of intra- and extrauterine implantation sites. Histochem. Cell Biol 2002, 117, 401–414. [Google Scholar]
- Huppertz, B. The feto-maternal interface: Setting the stage for potential immune interactions. Semin. Immunopathol 2007, 29, 83–94. [Google Scholar]
- Damsky, C.H.; Fisher, S.J. Trophoblast pseudo-vasculogenesis: Faking it with endothelial adhesion receptors. Curr. Opin. Cell Biol 1998, 10, 660–666. [Google Scholar]
- Damsky, C.H.; Fitzgerald, M.L.; Fisher, S.J. Distribution patterns of extracellular matrix components and adhesion receptors are intricately modulated during first trimester cytotrophoblast differentiation along the invasive pathway, in vivo. J. Clin. Invest 1992, 89, 210–222. [Google Scholar]
- Trowsdale, J.; Moffett, A. NK receptor interactions with MHC class I molecules in pregnancy. Semin. Immunol 2008, 20, 317–320. [Google Scholar]
- Fisher, S.J.; Leitch, M.S.; Kantor, M.S.; Basbaum, C.B.; Kramer, R.H. Degradation of extracellular matrix by the trophoblastic cells of first-trimester human placentas. J. Cell. Biochem 1985, 27, 31–41. [Google Scholar]
- Al-Lamki, R.S.; Skepper, J.N.; Burton, G.J. Are human placental bed giant cells merely aggregates of small mononuclear trophoblast cells? An ultrastructural and immunocytochemical study. Hum. Reprod 1999, 14, 496–504. [Google Scholar]
- Cartwright, J.E.; Fraser, R.; Leslie, K.; Wallace, A.E.; James, J.L. Remodelling at the maternal-fetal interface: Relevance to human pregnancy disorders. Reproduction 2010, 140, 803–803. [Google Scholar]
- Kaufmann, P.; Black, S.; Huppertz, B. Endovascular trophoblast invasion: Implications for the pathogenesis of intrauterine growth retardation and preeclampsia. Biol. Reprod 2003, 69, 1–7. [Google Scholar]
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The uterine spiral arteries in human pregnancy: Facts and controversies. Placenta 2006, 27, 939–958. [Google Scholar]
- Zhou, Y.; Fisher, S.J.; Janatpour, M.; Genbacev, O.; Dejana, E.; Wheelock, M.; Damsky, C.H. Human cytotrophoblasts adopt a vascular phenotype as they differentiate. A strategy for successful endovascular invasion? J. Clin. Invest 1997, 99, 2139–2151. [Google Scholar]
- Khankin, E.V.; Royle, C.; Karumanchi, S.A. Placental vasculature in health and disease. Semin. Thromb. Hemost 2010, 36, 309–320. [Google Scholar]
- Kam, E.P.Y.; Gardner, L.; Loke, Y.W.; King, A. The role of trophoblast in the physiological change in decidual spiral arteries. Hum. Reprod 1999, 14, 2131–2138. [Google Scholar]
- Reynolds, L.P.; Redmer, D.A. Angiogenesis in the Placenta. Biol. Reprod 2001, 64, 1033–1040. [Google Scholar]
- Danihel, L.; Gomolcák, P.; Korbel, M.; Pruzinec, J.; Vojtassák, J.; Janík, P.; Babál, P. Expression of proliferation and apoptotic markers in human placenta during pregnancy. Acta Histochem 2002, 104, 335–338. [Google Scholar]
- Huppertz, B. IFPA award in placentology lecture: Biology of the placental syncytiotrophoblast—Myths and facts. Placenta 2010, 31, S75–S81. [Google Scholar]
- Allaire, A.D.; Ballenger, K.A.; Wells, S.R.; McMahon, M.J.; Lessey, B.A. Placental apoptosis in preeclampsia. Obstet. Gynecol 2000, 96, 271–276. [Google Scholar]
- Chen, C.P.; Bajoria, R.; Aplin, J.D. Decreased vascularization and cell proliferation in placentas of intrauterine growth-restricted fetuses with abnormal umbilical artery flow velocity waveforms. Am. J. Obstet. Gynecol 2002, 187, 764–769. [Google Scholar]
- Krebs, C.; Macara, L.M.; Leiser, R.; Bowman, A.W.; Greer, I.A.; Kingdom, J.C. Intrauterine growth restriction with absent end-diastolic flow velocity in the umbilical artery is associated with maldevelopment of the placental terminal villous tree. Am. J. Obstet. Gynecol 1996, 175, 1534–1542. [Google Scholar]
- Jackson, M.R.; Walsh, A.J.; Morrow, R.J.; Mullen, J.B.; Lye, S.J.; Ritchie, J.W. Reduced placental villous tree elaboration in small-for-gestational-age pregnancies: Relationship with umbilical artery Doppler waveforms. Am. J. Obstet. Gynecol 1995, 172, 518–525. [Google Scholar]
- Mouillet, J.F.; Chu, T.; Nelson, D.M.; Mishima, T.; Sadovsky, Y. MiR-205 silences MED1 in hypoxic primary human trophoblasts. FASEB J 2010, 24, 2030–2039. [Google Scholar]
- Forbes, K.; Farrokhnia, F.; Aplin, J.D.; Westwood, M. Dicer-dependent miRNAs provide an endogenous restraint on cytotrophoblast proliferation. Placenta 2012, 33, 581–585. [Google Scholar]
- Ji, L.; Brkić, J.; Liu, M.; Fu, G.; Peng, C.; Wang, Y.L. Placental trophoblast cell differentiation: Physiological regulation and pathological relevance to preeclampsia. Mol. Aspects Med. 2012. [Google Scholar] [CrossRef]
- Seitz, H.; Royo, H.; Bortolin, M.L.; Lin, S.P.; Ferguson-Smith, A.C.; Cavaillé, J. A large imprinted microRNA gene cluster at the mouse Dlk1-Gtl2 domain. Genome Res 2004, 14, 1741–1748. [Google Scholar]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar]
- Cai, X.; Hagedorn, C.H.; Cullen, B.R. Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA 2004, 10, 1957–1966. [Google Scholar]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar]
- Lund, E.; Güttinger, S.; Calado, A.; Dahlberg, J.E.; Kutay, U. Nuclear export of microRNA precursors. Science 2004, 303, 95–98. [Google Scholar]
- Denli, A.M.; Tops, B.B.; Plasterk, R.H.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar]
- Gregory, R.I.; Yan, K.P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The microprocessor complex mediates the genesis of microRNAs. Nature 2004, 432, 235–240. [Google Scholar]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, 835–840. [Google Scholar] [Green Version]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet 2004, 5, 522–531. [Google Scholar]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar]
- Huang, Y.; Zou, Q.; Song, H.; Song, F.; Wang, L.; Zhang, G.; Shen, X. A study of miRNAs targets prediction and experimental validation. Protein Cell 2010, 1, 979–986. [Google Scholar]
- Nilsen, T.W. Mechanisms of microRNA-mediated gene regulation in animal cells. Trends Genet 2007, 23, 243–249. [Google Scholar]
- Zhao, S.; Liu, M.F. Mechanisms of microRNA-mediated gene regulation. Sci. China C Life Dci 2009, 52, 1111–1116. [Google Scholar]
- Valencia-Sanchez, M.A.; Liu, J.; Hannon, G.; Parker, R. Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev 2006, 20, 515–524. [Google Scholar]
- Zeng, Y.; Yi, R.; Cullen, B.R. MicroRNAs and small interfering RNAs can inhibit mRNA expression by similar mechanisms. Proc. Natl. Acad. Sci. USA 2003, 100, 9779–9784. [Google Scholar]
- Lytle, J.R.; Yario, T.A.; Steitz, J.A. Target mRNAs are repressed as efficiently by microRNA-binding sites in the 5′ UTR as in the 3′ UTR. Proc. Natl. Acad. Sci. USA 2007, 104, 9667–9672. [Google Scholar]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: MicroRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar]
- Lee, R.C.; Ambros, V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001, 294, 862–864. [Google Scholar]
- Lau, N.C.; Lim, L.P.; Weinstein, E.G.; Bartel, D.P. An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans. Science 2001, 294, 858–862. [Google Scholar]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–1414. [Google Scholar]
- Guo, L.; Yang, Q.; Lu, J.; Li, H.; Ge, Q.; Gu, W.; Bai, Y.; Lu, Z. A comprehensive survey of miRNA repertoire and 3′ addition events in the placentas of patients with pre-eclampsia from high-throughput sequencing. PLoS One 2011, 6, e21072. [Google Scholar]
- Luo, S.S.; Ishibashi, O.; Ishikawa, G.; Ishikawa, T.; Katayama, A.; Mishima, T.; Takizawa, T.; Shigihara, T.; Goto, T.; Izumi, A.; et al. Human villous trophoblasts express and secrete placenta-specific microRNAs into maternal circulation via exosomes. Biol. Reprod. 2009, 81, 717–729. [Google Scholar]
- Pineles, B.L.; Romero, R.; Montenegro, D.; Tarca, A.L.; Han, Y.M.; Kim, Y.M.; Draghici, S.; Espinoza, J.; Kusanovic, J.P.; Mittal, P.; et al. Distinct subsets of microRNAs are expressed differentially in the human placentas of patients with preeclampsia. Am. J. Obstet. Gynecol 2007, 196, 261.e1–261.e6. [Google Scholar]
- Umemura, K.; Ishioka, S.; Endo, T.; Ezaka, Y.; Takahashi, M.; Saito, T. Roles of microRNA-34a in the pathogenesis of placenta accreta. J. Obstet. Gynaecol. Res 2013, 39, 67–74. [Google Scholar]
- Tolstrup, N.; Nielsen, P.S.; Kolberg, J.G.; Frankel, A.M.; Vissing, H.; Kauppinen, S. OligoDesign: Optimal design of LNA (locked nucleic acid) oligonucleotide capture probes for gene expression profiling. Nucleic Acids Res 2003, 31, 3758–6372. [Google Scholar]
- Varallyay, E.; Burgyan, J.; Havelda, Z. MicroRNA detection by northern blotting using locked nucleic acid probes. Nat. Protoc 2008, 3, 190–196. [Google Scholar]
- Miska, E.A.; Alvarez-Saavedra, E.; Townsend, M.; Yoshii, A.; Sestan, N.; Rakic, P.; Constantine-Paton, M.; Horvitz, H.R. Microarray analysis of microRNA expression in the developing mammalian brain. Genome Biol 2004, 5, R68. [Google Scholar]
- Kloosterman, W.P.; Wienholds, E.; de Bruijn, E.; Kauppinen, S.; Plasterk, R.H. In situ detection of miRNAs in animal embryos using LNA-modified oligonucleotide probes. Nat. Methods 2006, 3, 27–29. [Google Scholar]
- Wienholds, E.; Kloosterman, W.P.; Miska, E.; Alvarez-Saavedra, E.; Berezikov, E.; de Bruijn, E.; Horvitz, H.R.; Kauppinen, S.; Plasterk, R.H. MicroRNA expression in zebrafish embryonic development. Science 2005, 309, 310–311. [Google Scholar]
- Morales-Prieto, D.M.; Chaiwangyen, W.; Ospina-Prieto, S.; Schneider, U.; Herrmann, J.; Gruhn, B.; Markert, U.R. MicroRNA expression profiles of trophoblastic cells. Placenta 2012, 33, 725–734. [Google Scholar]
- Donker, R.B.; Mouillet, J.F.; Chu, T.; Hubel, C.A.; Stolz, D.B.; Morelli, A.E.; Sadovsky, Y. The expression profile of C19MC microRNAs in primary human trophoblast cells and exosomes. Mol. Hum. Reprod 2012, 18, 417–424. [Google Scholar]
- Bentwich, I.; Avniel, A.; Karov, Y.; Aharonov, R.; Gilad, S.; Barad, O.; Barzilai, A.; Einat, P.; Einav, U.; Meiri, E.; et al. Identification of hundreds of conserved and nonconserved human microRNAs. Nat. Genet 2005, 37, 766–770. [Google Scholar]
- Bortolin-Cavaillé, M.L.; Dance, M.; Weber, M.; Cavaillé, J. C19MC microRNAs are processed from introns of large Pol-II, non-protein-coding transcripts. Nucleic Acids Res 2009, 37, 3464–3463. [Google Scholar]
- Noguer-Dance, M.; Abu-Amero, S.; Al-Khtib, M.; Lefèvre, A.; Coullin, P.; Moore, G.E.; Cavaillé, J. The primate-specific microRNA gene cluster (C19MC) is imprinted in the placenta. Hum. Mol. Genet 2010, 19, 3566–3582. [Google Scholar]
- Lewis, A.; Mitsuya, K.; Umlauf, D.; Smith, P.; Dean, W.; Walter, J.; Higgins, M.; Feil, R.; Reik, W. Imprinting on distal chromosome 7 in the placenta involves repressive histone methylation independent of DNA methylation. Nat. Genet 2004, 36, 1291–1295. [Google Scholar]
- Genbacev, O.; Joslin, R.; Damsky, C.H.; Polliotti, B.M.; Fisher, S.J. Hypoxia alters early gestation human cytotrophoblast differentiation/invasion in vitro and models the placental defects that occur in preeclampsia. J. Clin. Invest 1996, 97, 540–50. [Google Scholar]
- Caniggia, I.; Winter, J.; Lye, S.J.; Post, M. Oxygen and placental development during the first trimester: Implications for the pathophysiology of pre-eclampsia. Placenta 2000, 21, S25–S30. [Google Scholar]
- Adelman, D.M.; Gertsenstein, M.; Nagy, A.; Simon, M.C.; Maltepe, E. Placental cell fates are regulated in vivo by HIF-mediated hypoxia responses. Genes Dev 2000, 14, 3191–3203. [Google Scholar]
- Chan, S.Y.; Loscalzo, J. MicroRNA-210: A unique and pleiotropic hypoxamir. Cell Cycle 2010, 9, 1072–1083. [Google Scholar]
- Devlin, C.; Greco, S.; Martelli, F.; Ivan, M. MiR-210: More than a silent player in hypoxia. IUBMB Life 2011, 63, 94–100. [Google Scholar]
- Camps, C.; Buffa, F.M.; Colella, S.; Moore, J.; Sotiriou, C.; Sheldon, H.; Harris, A.L.; Gleadle, J.M.; Ragoussis, J. hsa-miR-210 Is induced by hypoxia and is an independent prognostic factor in breast cancer. Clin. Cancer Res 2008, 14, 1340–1348. [Google Scholar]
- Cummins, E.P.; Taylor, C.T. Hypoxia-responsive transcription factors. Pflugers Arch 2005, 450, 363–371. [Google Scholar]
- Kulshreshtha, R.; Ferracin, M.; Wojcik, S.E.; Garzon, R.; Alder, H.; Agosto-Perez, F.J.; Davuluri, R.; Liu, C.G.; Croce, C.M.; Negrini, M.; et al. A microRNA signature of hypoxia. Mol. Cell. Biol 2007, 27, 1859–1867. [Google Scholar]
- Kelly, T.J.; Souza, A.L.; Clish, C.B.; Puigserver, P. A hypoxia-induced positive feedback loop promotes hypoxia-inducible factor 1alpha stability through miR-210 suppression of glycerol-3-phosphate dehydrogenase 1-like. Mol. Cell. Biol 2011, 31, 2696–2706. [Google Scholar]
- Zhang, Y.; Fei, M.; Xue, G.; Zhou, Q.; Jia, Y.; Li, L.; Xin, H.; Sun, S. Elevated levels of hypoxia-inducible microRNA-210 in pre-eclampsia: New insights into molecular mechanisms for the disease. J. Cell. Mol. Med 2012, 16, 249–259. [Google Scholar]
- Bamberger, A.M.; Bamberger, C.M.; Aupers, S.; Milde-Langosch, K.; Löning, T.; Makrigiannakis, A. Expression pattern of the activating protein-1 family of transcription factors in the human placenta. Mol. Hum. Reprod 2004, 10, 223–228. [Google Scholar]
- Marzioni, D.; Todros, T.; Cardaropoli, S.; Rolfo, A.; Lorenzi, T.; Ciarmela, P.; Romagnoli, R.; Paulesu, L.; Castellucci, M. Activating protein-1 family of transcription factors in the human placenta complicated by preeclampsia with and without fetal growth restriction. Placenta 2010, 31, 919–927. [Google Scholar]
- Dai, Y.; Diao, Z.; Sun, H.; Li, R.; Qiu, Z.; Hu, Y. MicroRNA-155 is involved in the remodelling of human-trophoblast-derived HTR-8/SVneo cells induced by lipopolysaccharides. Hum. Reprod 2011, 26, 1882–1891. [Google Scholar]
- Morales-Prieto, D.M.; Schleussner, E.; Markert, U.R. Reduction in miR-141 is induced by leukemia inhibitory factor and inhibits proliferation in choriocarcinoma cell line JEG-3. Am. J. Reprod. Immunol 2011, 66, 57–62. [Google Scholar]
- Avissar-Whiting, M.; Veiga, K.R.; Uhl, K.M.; Maccani, M.A.; Gagne, L.A.; Moen, E.L.; Marsit, C.J. Bisphenol A exposure leads to specific microRNA alterations in placental cells. Reprod. Toxicol 2010, 29, 401–406. [Google Scholar]
- Maccani, M.A.; Avissar-Whiting, M.; Banister, C.E.; McGonnigal, B.; Padbury, J.F.; Marsit, C.J. Maternal cigarette smoking during pregnancy is associated with downregulation of miR-16, miR-21, and miR-146a in the placenta. Epigenetics 2010, 5, 583–589. [Google Scholar]
- Tsai, K.W.; Kao, H.W.; Chen, H.C.; Chen, S.J.; Lin, W.C. Epigenetic control of the expression of a primate-specific microRNA cluster in human cancer cells. Epigenetics 2009, 4, 587–592. [Google Scholar]
- Muralimanoharan, S.; Maloyan, A.; Mele, J.; Guo, C.; Myatt, L.G.; Myatt, L. MIR-210 modulates mitochondrial respiration in placenta with preeclampsia. Placenta 2012, 33, 816–823. [Google Scholar]
- Ishibashi, O.; Ohkuchi, A.; Ali, M.M.; Kurashina, R.; Luo, S.S.; Ishikawa, T.; Takizawa, T.; Hirashima, C.; Takahashi, K.; Migita, M.; et al. Hydroxysteroid (17-beta) dehydrogenase 1 is dysregulated by miR-210 and miR-518c that are aberrantly expressed in preeclamptic placentas: A novel marker for predicting preeclampsia. Hypertension 2012, 59, 265–273. [Google Scholar]
- Gao, W.L.; Liu, M.; Yang, Y.; Yang, H.; Liao, Q.; Bai, Y.; Li, Y.X.; Li, D.; Peng, C.; Wang, Y.L. The imprinted H19 gene regulates human placental trophoblast cell proliferation via encoding miR-675 that targets Nodal Modulator 1 (NOMO1). RNA Biol 2012, 9, 1002–1010. [Google Scholar]
- Maccani, M.A.; Padbury, J.F.; Marsit, C.J. miR-16 and miR-21 expression in the placenta is associated with fetal growth. PLoS One 2011, 6, e21210. [Google Scholar]
- Zhang, Y.; Diao, Z.; Su, L.; Sun, H.; Li, R.; Cui, H.; Hu, Y. MicroRNA-155 contributes to preeclampsia by down-regulating CYR61. Am. J. Obstet. Gynecol 2010, 202, 466.e1–466.e7. [Google Scholar]
- Wang, Y.; Fan, H.; Zhao, G.; Liu, D.; Du, L.; Wang, Z.; Hu, Y.; Hou, Y. MiR-16 inhibits the proliferation and angiogenesis-regulating potential of mesenchymal stem cells in severe pre-eclampsia. FEBS J 2012, 289, 4510–4524. [Google Scholar]
- Nadeem, L.; Munir, S.; Fu, G.; Dunk, C.; Baczyk, D.; Caniggia, I.; Lye, S.; Peng, C. Nodal signals through activin receptor-like kinase 7 to inhibit trophoblast migration and invasion: Implication in the pathogenesis of preeclampsia. Am. J. Pathol 2011, 178, 1177–1189. [Google Scholar]
- Munir, S.; Xu, G.; Wu, Y.; Yang, B.; Lala, P.K.; Peng, C. Nodal and ALK7 inhibit proliferation and induce apoptosis in human trophoblast cells. J. Biol. Chem 2004, 279, 31277–31286. [Google Scholar]
- Yu, L.; Li, D.; Liao, Q.P.; Yang, H.X.; Cao, B.; Fu, G.; Ye, G.; Bai, Y.; Wang, H.; Cui, N.; et al. High levels of activin a detected in preeclamptic placenta induce trophoblast cell apoptosis by promoting nodal signaling. J. Clin. Endocrinol. Metab 2012, 97, E1370–E1379. [Google Scholar]
- Xu, S.; Linher-Melville, K.; Yang, B.B.; Wu, D.; Li, J. Micro-RNA378 (miR-378) regulates ovarian estradiol production by targeting aromatase. Endocrinology 2011, 152, 3941–3951. [Google Scholar]
- Lee, D.Y.; Deng, Z.; Wang, C.H.; Yang, B.B. MicroRNA-378 promotes cell survival, tumor growth, and angiogenesis by targeting SuFu and Fus-1 expression. Proc. Natl. Acad. Sci. USA 2007, 104, 20350–20355. [Google Scholar]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol 2012, 14, 659–665. [Google Scholar]
- Forbes, K.; Westwood, M.; Baker, P.N.; Aplin, J.D. Insulin-like growth factor I and II regulate the life cycle of trophoblast in the developing human placenta. Am. J. Physiol. Cell Physiol 2008, 294, C1313–C1322. [Google Scholar]
- Segura, M.F.; Hanniford, D.; Menendez, S.; Reavie, L.; Zou, X.; Alvarez-Diaz, S.; Zakrzewski, J.; Blochin, E.; Rose, A.; Bogunovic, D.; et al. Aberrant miR-182 expression promotes melanoma metastasis by repressing FOXO3 and microphthalmia-associated transcription factor. Proc. Natl. Acad. Sci. USA 2009, 106, 1814–1819. [Google Scholar]
- Chelbi, S.T.; Vaiman, D. Genetic and epigenetic factors contribute to the onset of preeclampsia. Mol. Cell Endocrinol 2008, 282, 120–129. [Google Scholar]
- Nadeem, L.; Brkic, J.; Chen, Y.F.; Bui, T.; Munir, S.; Peng, C. Cytoplasmic mislocalization of p27 and cdk2 mediates the anti-migratory and anti-proliferative effects of Nodal in human trophoblast cells. J. Cell Sci 2012. [Google Scholar] [CrossRef]
- Caniggia, I.; Mostachfi, H.; Winter, J.; Gassmann, M.; Lye, S.J.; Kuliszewski, M.; Post, M. Hypoxia-inducible factor-1 mediates the biological effects of oxygen on human trophoblast differentiation through TGFbeta(3). J. Clin. Invest 2000, 105, 577–587. [Google Scholar]
- Oh, S.P.; Yeo, C.Y.; Lee, Y.; Schrewe, H.; Whitman, M.; Li, E. Activin type IIA and IIB receptors mediate Gdf11 signaling in axial vertebral patterning. Genes Dev 2002, 16, 2749–2754. [Google Scholar]
- Pang, R.T.; Leung, C.O.; Ye, T.M.; Liu, W.; Chiu, P.C.; Lam, K.K.; Lee, K.F.; Yeung, W.S. MicroRNA-34a suppresses invasion through downregulation of Notch1 and Jagged1 in cervical carcinoma and choriocarcinoma cells. Carcinogenesis 2010, 31, 1037–1044. [Google Scholar]
- Montenegro, D.; Romero, R.; Kim, S.S.; Tarca, A.L.; Draghici, S.; Kusanovic, J.P.; Kim, J.S.; Lee, D.C.; Erez, O.; Gotsch, F.; et al. Expression patterns of microRNAs in the chorioamniotic membranes: A role for microRNAs in human pregnancy and parturition. J. Pathol 2009, 217, 113–121. [Google Scholar]
- Yang, W.J.; Yang, D.D.; Na, S.; Sandusky, G.E.; Zhang, Q.; Zhao, G. Dicer is required for embryonic angiogenesis during mouse development. J. Biol. Chem 2005, 280, 9330–9335. [Google Scholar]
- Red-Horse, K.; Kapidzic, M.; Zhou, Y.; Feng, K.T.; Singh, H.; Fisher, S.J. EPHB4 regulates chemokine-evoked trophoblast responses: A mechanism for incorporating the human placenta into the maternal circulation. Development 2005, 132, 4097–4106. [Google Scholar]
- Chennakesava, C.S.; di Santo, S.; Ziemiecki, A.; Schneider, H.; Andres, A.C. Differential expression of the receptor tyrosine kinase EphB4 and its ligand Ephrin-B2 during human placental development. Placenta 2006, 27, 959–967. [Google Scholar]
- Wang, W.; Feng, L.; Zhang, H.; Hachy, S.; Satohisa, S.; Laurent, L.C.; Parast, M.; Zheng, J.; Chen, D.B. Preeclampsia up-regulates angiogenesis-associated microRNA (i.e., miR-17, -20a, and -20b) that target ephrin-B2 and EPHB4 in human placenta. J. Clin. Endocrinol. Metab 2012, 97, E1051–E1059. [Google Scholar]
- Burton, G.J.; Charnock-Jones, D.S.; Jauniaux, E. Regulation of vascular growth and function in the human placenta. Reproduction 2009, 138. [Google Scholar]
- Muramatsu, F.; Kidoya, H.; Naito, H.; Sakimoto, S.; Takakura, N. microRNA-125b inhibits tube formation of blood vessels through translational suppression of VE-cadherin. Oncogene 2013, 32, 414–421. [Google Scholar]
- Alpini, G.; Glaser, S.S.; Zhang, J.P.; Francis, H.; Han, Y.; Gong, J.; Stokes, A.; Francis, T.; Hughart, N.; Hubble, L.; et al. Regulation of placenta growth factor by microRNA-125b in hepatocellular cancer. J. Hepatol 2011, 55, 1339–1345. [Google Scholar]
- Liu, L.Z.; Li, C.; Chen, Q.; Jing, Y.; Carpenter, R.; Jiang, Y.; Kung, H.-F.; Lai, L.; Jiang, B.-H. MiR-21 induced angiogenesis through AKT and ERK activation and HIF-1alpha expression. PLoS One 2011, 6, e19139. [Google Scholar]
- Fang, L.; Deng, Z.; Shatseva, T.; Yang, J.; Peng, C.; Du, W.W.; Yee, A.J.; Ang, L.C.; He, C.; Shan, S.W.; et al. MicroRNA miR-93 promotes tumor growth and angiogenesis by targeting integrin-beta8. Oncogene 2011, 30, 806–821. [Google Scholar]
- Zhu, J.; Motejlek, K.; Wang, D.; Zang, K.; Schmidt, A.; Reichardt, L.F. β8 integrins are required for vascular morphogenesis in mouse embryos. Development 2002, 129, 2891–2903. [Google Scholar]
- Babawale, M.O.; van Noorden, S.; Pignatelli, M.; Stamp, G.W.; Elder, M.G.; Sullivan, M.H. Morphological interactions of human first trimester placental villi co-cultured with decidual explants. Hum. Reprod 1996, 11, 444–450. [Google Scholar]
- Dunk, C.; Petkovic, L.; Baczyk, D.; Rossant, J.; Winterhager, E.; Lye, S. A novel in vitro model of trophoblast-mediated decidual blood vessel remodeling. Lab. Invest 2003, 83, 1821–1828. [Google Scholar]
- Zhao, C.; Dong, J.; Jiang, T.; Shi, Z.; Yu, B.; Zhu, Y.; Chen, D.; Xu, J.; Huo, R.; Dai, J. Early second-trimester serum miRNA profiling predicts gestational diabetes mellitus. PLoS One 2011, 6, e23925. [Google Scholar]
- Higashijima, A.; Miura, K.; Mishima, H.; Kinoshita, A.; Jo, O.; Abe, S.; Hasegawa, Y.; Miura, S.; Yamasaki, K.; Yoshida, A.; et al. Characterization of placenta-specific microRNAs in fetal growth restriction pregnancy. Prenat. Diagn 2013. [Google Scholar] [CrossRef]
- Sibai, B.; Dekker, G.; Kupferminc, M. Pre-eclampsia. Lancet 2005, 365, 785–799. [Google Scholar]
- Faye-Petersen, O.M. The placenta in preterm birth. J. Clin. Pathol 2008, 61, 1261–1275. [Google Scholar]
- Cui, Y.; Wang, W.; Dong, N.; Lou, J.; Srinivasan, D.K.; Cheng, W.; Huang, X.; Liu, M.; Fang, C.; Peng, J.; et al. Role of corin in trophoblast invasion and uterine spiral artery remodelling in pregnancy. Nature 2012, 484, 246–250. [Google Scholar]
- Morales Prieto, D.M.; Markert, U.R. MicroRNAs in pregnancy. J. Reprod. Immunol 2011, 88, 106–111. [Google Scholar]
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; et al. Serum microRNAs are promising novel biomarkers. PLoS One 2008, 3, e3148. [Google Scholar]
- Kotlabova, K.; Doucha, J.; Hromadnikova, I. Placental-specific microRNA in maternal circulation—Identification of appropriate pregnancy-associated microRNAs with diagnostic potential. J. Reprod. Immunol 2011, 89, 185–191. [Google Scholar]
- Chim, S.S.; Shing, T.K.; Hung, E.C.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.; Lo, Y.M. Detection and characterization of placental microRNAs in maternal plasma. Clin. Chem 2008, 54, 482–490. [Google Scholar]
- Miura, K.; Miura, S.; Yamasaki, K.; Higashijima, A.; Kinoshita, A.; Yoshiura, K.; Masuzaki, H. Identification of pregnancy-associated microRNAs in maternal plasma. Clin. Chem 2010, 56, 1767–1771. [Google Scholar]
- Pollheimer, J.; Knofler, M. Signalling pathways regulating the invasive differentiation of human trophoblasts: A review. Placenta 2005, 26, S21–S30. [Google Scholar]


{kind=link}
| miRNA | Function | Target genes | Cells tested | References |
|---|---|---|---|---|
| miR-210 | ↓ migration/invasion | EFNA3, HOXA9 | CTBs (1st trimester) | [105] |
| ↓ iron metabolism | ISCU | BeWo, Swan71, placental tissue | [12] | |
| ↓ mitochondrial respiration | ISCU | Primary trophoblasts | [113] | |
| ↓ steroid metabolism | HSD17B1 | BeWo | [114] | |
| miR-376c | ↑ proliferation/invasion/migration | ALK5, ALK7 | HTR8/SVneo, placental explants | [16] |
| miR-378a-5p | ↑ proliferation/invasion/migration | NODAL | HTR8/SVneo, placental explants | [19] |
| miR-195 | ↑ invasion | ActRIIA | HTR8/SVneo | [20] |
| miR-675 | ↓ proliferation | NOMO-1 | JEG-3 | [115] |
| miR-21 | ↑ proliferation/invasion | PTEN | TCL-1 | [90,116] |
| miR-155 | ↓ proliferation/migration | CCND1 | HTR8/SVneo | [117] |
| miR-16 | ↓ proliferation | CCNE1 | dMSC | [118] |
| ↓ invasion | HTR8/SVneo | |||
| ↓ angiogenesis | HUVEC | |||
| miR-34a | ↓ proliferation/invasion | NOTCH1, JAG1 | JAR | [119] |
| miR-29b | ↑ apoptosis | MCL1, MMP2, VEGFA, ITGB1 | HTR8/SVneo, BeWo | [23] |
| ↓ invasion | ||||
| ↓ angiogenesis | ||||
| Compromised pregnancy | Sample | Regulation | miRNAs | Method of detection | References |
|---|---|---|---|---|---|
| PE | Placenta | Up-regulated | miR-20b | Microarray & qRT-PCR | [136] |
| miR-16, miR-29b, miR-195, miR-26b, miR-181a, miR-335 and miR-222 | [25] | ||||
| miR-210, miR-152 and miR-518b | [26] | ||||
| miR-516a-5p, miR-512-3p, miR-2277 and miR-524-3p | Microarray | [136] | |||
| miR-182 and miR-210 | qRT-PCR | [12,83] | |||
| miR-17 and miR-20a | [136] | ||||
| miR-155 | [117] | ||||
| miR-210, miR-193b, miR-144*, miR-193*, miR-18a, miR-185, miR-19a, miR-590-5p, miR-142-3p, miR-451, miR-22*, miR-526b*, miR-520a-3p, miR-10b, miR-20a, miR-518f*, miR-146b-5p, miR-517c, miR-518c, miR-5258-5p, miR-519e* and miR-126* | High-throughput sequencing & qRT-PCR-based array | [114] | |||
| Plasma | Up-regulated | miR-210 | qRT-PCR | [105] | |
| Placenta | Down-regulated | miR-18a, miR-411, miR-377, miR-363 and miR-542-3p | Microarray & qRT-PCR | [26] | |
| miR-101, miR-10b, miR-218, miR-590, miR-204, miR-32, miR-126*, miR-19a, miR-154*, miR-625, miR-144, miR-195, miR-150, miR-1, miR-18b and miR-450 miR-151-3p, miR-146a, miR-192 and miR-34c-5p | Microarray | [26] | |||
| [136] | |||||
| miR-376c | qRT-PCR | [16] | |||
| miR-378a-5p | [19] | ||||
| miR-195 | [20] | ||||
| miR-675 | [115] | ||||
| Plasma | Down-regulated | miR-376c | qRT-PCR | [16] | |
| SGA | Placenta | Up-regulated | miR-210 | qRT-PCR | [12] |
| Down-regulated | miR-16 and miR-21 | qRT-PCR | [116] | ||
| PE + SGA | Placenta | Up-regulated | miR-210 | qRT-PCR | [12,83] |
| PTB | Fetal membranes | Up-regulated | miR-25, miR-338, miR-101, miR-449, miR-154, miR-135a, miR-142-3p, miR-202* and miR-136 | Microarray | [132] |
| miR-338, miR-449, miR-136 and miR-199a* | qRT-PCR | [132] | |||
| GDM | Serum | Down-regulated | miR-132, miR-29a and miR-222 | qRT-PCR | [145] |
| IUGR | Placenta | Down-regulated | miR-518b, miR-1323, miR-516b, miR-515-5p, miR-520h, miR-519d and miR-526b | qRT-PCR | [146] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fu, G.; Brkić, J.; Hayder, H.; Peng, C. MicroRNAs in Human Placental Development and Pregnancy Complications. Int. J. Mol. Sci. 2013, 14, 5519-5544. https://doi.org/10.3390/ijms14035519
Fu G, Brkić J, Hayder H, Peng C. MicroRNAs in Human Placental Development and Pregnancy Complications. International Journal of Molecular Sciences. 2013; 14(3):5519-5544. https://doi.org/10.3390/ijms14035519
Chicago/Turabian StyleFu, Guodong, Jelena Brkić, Heyam Hayder, and Chun Peng. 2013. "MicroRNAs in Human Placental Development and Pregnancy Complications" International Journal of Molecular Sciences 14, no. 3: 5519-5544. https://doi.org/10.3390/ijms14035519