

Abstract
Background/Aim: Oncolytic adenoviruses (Ads) (OAds) are gaining attention as an effective remedy for pancreatic cancer. Most OAds are based on human Ad serotype 5 (Ad5) (OAd5); however, two major drawbacks of OAd5 have been reported. Expression of coxsackievirus-adenovirus receptor, a primary infection receptor of Ad5, is often decreased on malignant tumor cells, including pancreatic cancers. More than 60% of adults have neutralizing antibodies against Ad5. Previously, we developed an OAd composed of Ad serotype 35 (Ad35) (OAd35). Ad35 recognizes CD46, which is often up-regulated on pancreatic cancers. In addition, only 20% or fewer adults have anti-Ad35 neutralizing antibodies. Materials and Methods: We examined the tumor cell lysis activities of OAd35 in the four human pancreatic cancer cell lines in the presence and absence of human serum. The tumor growth suppression effects of OAd35 after local and systemic administration were evaluated in nude mice bearing human pancreatic tumors. Results: OAd35 showed higher levels of tumor cell lysis activities than OAd5 in the human pancreatic cancer cell lines AsPC-1 and BxPC-3. Although the in vitro tumor cell lysis activities of OAd5 against MIA PaCa-2 and PANC-1 cells were strongly attenuated in the presence of human serum, OAd35 mediated comparable levels of tumor cell lysis in the presence and absence of human serum. Systemic administration of OAd5 did not mediate significant growth inhibition against the subcutaneous BxPC-3 tumor. On the other hand, OAd35 significantly suppressed tumor growth. Conclusion: OAd35 would be suitable as an alternative anticancer agent for pancreatic cancer.
- Oncolytic virus
- oncolytic adenovirus
- pancreatic cancer
- CD46
- anti-adenovirus antibody
Pancreatic cancer is the fourth leading cause of cancer-related death (1, 2). The 5-year survival rates are about 10% for all stages combined and 15-20% for surgically resected patients (3-6). Due to the lack of effective treatments without severe toxicity profiles, the 5-year survival rates of patients who cannot undergo surgical resection have remained unchanged (7, 8). Since pancreatic cancer is often an aggressive malignancy and does not exhibit obvious symptoms, many patients present with unresectable disease or distant metastases at the time of diagnosis. The 5-year survival rate of patients with unresectable disease is less than 5%. Moreover, undetectable distant metastases make a major contribution to the low 5-year survival rate of patients undergoing surgical resection (9). Thus, there is an urgent need for the development of novel, systemically deliverable antitumor agents that can kill not only the primary pancreatic cancer but also undetectable distant metastatic cancer.
Recently, oncolytic viruses (OVs), which replicate in tumor cells and mediate antitumor effects without apparent toxicity in normal cells, have received a lot of attention as an effective remedy for pancreatic cancer. OVs mediate not only direct tumor cell lysis but also activation of antitumor immunity, leading to efficient antitumor effects (10, 11). Among the many available types of OVs, an OV based on human adenovirus (Ad) (OAd) is the most widely used due to its attractive properties (12-15). Clinical trials using OAds for the treatment of pancreatic cancers have been conducted (16-18) or are ongoing [NCT03225989, NCT02749331].
Conventional OAds are based on species C human Ad serotype 5 (Ad5) (OAd5). However, there are two major concerns regarding the use of OAd5 to treat pancreatic cancer. First, the expression of the primary infection receptor of OAd5, the coxsackievirus-adenovirus receptor (CAR), is often decreased on malignant tumors, including pancreatic cancers (19, 20). Moreover, since CAR is involved in forming tight junctions via homophilic interactions, CAR expression is often reduced on pancreatic cancer cells with high metastatic potential. Second, more than 60% of adults have pre-existing neutralizing antibodies against Ad5 due to natural exposure (21-23). Although the impacts of neutralizing antibodies on the antitumor effects of OAd5 are controversial, anti-Ad5 neutralizing antibodies may hinder the systemic circulation of OAd5 and access to distant metastasis (24-26).
To overcome these drawbacks of OAd5, we developed OAd35 (27). OAd35 recognizes human complement regulatory protein CD46 as an infection receptor. CD46 expression is often up-regulated on malignant tumor cells (28, 29), including pancreatic cancer cells (30-33). A fiber-substituted Ad5 vector containing Ad35 proteins showed efficient transduction efficiencies in pancreatic cancer cells (34). Furthermore, seroprevalence against Ad35 in adults is only 20% or less (23, 35). These findings led us to hypothesize that OAd35 is a suitable OAd for pancreatic cancers.
In this study, we evaluated the in vitro tumor cell lysis activities of OAd35 in four human pancreatic cancer cell lines. In addition, we determined the effects of human serum on the in vitro tumor cell lysis activities of OAd35. Finally, the in vivo antitumor effects of OAd35 were examined following local and systemic administration in subcutaneous pancreatic cancer-bearing mice.
Materials and Methods
Cells. AsPC-1 and BxPC-3 cells were cultured in RPMI1640 (FUJIFILM Wako Pure Chemical, Osaka, Japan) supplemented with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific, San Jose, CA, USA), 100 μg/ml streptomycin (Nacalai Tesque, Kyoto, Japan), and 100 U/ml penicillin (Nacalai Tesque). MIA PaCa-2 and PANC-1 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) (FUJIFILM Wako Pure Chemical) supplemented with 10% FBS (Thermo Fisher Scientific), 100 μg/ml streptomycin (Nacalai Tesque), and 100 U/ml penicillin (Nacalai Tesque).
Oncolytic Ads. OAd5 and OAd35 containing the human telomerase reverse transcriptase (hTERT) promoter-driven E1A expression cassette were previously prepared (27). Virus particle (VP) titers were determined by the method of Maizel et al. (36). An OAd plasmid for a fiber-substituted OAd5 containing the fiber protein of Ad35 (OAd5-F35), pAdHM41-hmE1-E3(+)-F35, was constructed by using pHM5-hTERT-E1 (27) and pAdHM41-E3(+)-F35. pAdHM41-E3(+)-F35 was produced via homologous recombination between ClaI-digested pAdHM41-E3(+) (37) and ClaI/NdeI-digested pEco-ITR9 (38). OAd5-F35 was similarly produced as previously described (27).
Flow cytometric analysis. The expression levels of human CAR on tumor cells were measured by using mouse anti-CAR monoclonal antibody (RmcB) (Merck Millipore, Darmstadt, Germany) and phycoerythrin (PE)-labeled goat anti-mouse IgG secondary antibody (BD Pharmingen, San Diego, CA, USA). Cells were labeled with PE-conjugated mouse anti-CD46 monoclonal antibody (8E2) (Thermo Fisher Scientific) to determine the CD46 expression levels. Mouse IgG1, kappa isotype control antibody (BD Pharmingen), and PE-labeled mouse IgG1, kappa isotype control antibody (BioLegend, San Diego, CA, USA) were used as a negative control. Flow cytometric analysis was performed using a MACS Quant Analyzer (Miltenyi Biotec, Bergisch Gladbach, Germany). The data were analyzed using FlowJo™ v10.8 Software (BD Life Sciences, Ashland, OR, USA).
Tumor cell lysis activities of OAds in cultured tumor cells. Tumor cell lysis activities of OAds were determined by the crystal violet staining and WST-8 assay. Briefly, pancreatic cancer cells were seeded on a 24-well plate at a density of 0.5-1×105 cells/well. Following a 24-h incubation, cells were infected with OAds at the indicated VPs/cell. Cells were fixed with 4% paraformaldehyde phosphate buffer solution (FUJIFILM Wako Pure Chemical) for a 1-h incubation at room temperature following a 5-day infection. Then, the plates were treated with 1 ml of 2% crystal violet in 100% methanol.
In the WST-8 assay, cells were seeded on a 96-well plate at 0.5-1×104 cells/well. On the following day, cells were infected with OAds at 500 VP/cell. Cell viabilities were determined using a Cell Counting Kit-8 solution (Dojindo Laboratories, Kumamoto, Japan) on the indicated day points. Cell viabilities were calculated according to the manufacturer’s instructions.
Tumor cell lysis activities of OAds in the presence of human serum. Three batches of human adult serum isolated from healthy donors were purchased from KAC Co. Ltd (Kyoto, Japan). MIA PaCa-2 and PANC-1 cells were seeded on a 96-well plate at 2×104 cells/well. On the following day, OAds were pre-incubated with human serum for 30 min at room temperature. Cells were then infected with pre-incubated OAds at 500 VP/cell. Cell viabilities were determined after a 5-day incubation by the WST-8 assay as described above.
Determination of Ad genome copy numbers in tumor cells. Pancreatic cancer cells were seeded on a 12-well plate at a density of 1-2×105 cells/well. On the following day, cells were infected with OAds at 100 VP/cell. Total DNA was isolated using DNAzol (Molecular Research Center, Cincinnati, OH, USA) 24 and 72 h after infection. The Ad genome copy numbers were quantified by real-time PCR analysis using the primers for the Ad5 E4 and Ad35 E1A genes, a StepOnePlus System (Thermo Fisher Scientific), and THUNDERBIRD NEXT SYBR qPCR Mix reagents (TOYOBO, Osaka, Japan). The sequences of the primers are previously described (27).
Treatment of subcutaneous pancreatic cancers by OAd administration. BxPC-3 cell suspension (3×106 cells per mouse) was mixed with Matrigel (Corning, Corning, NY, USA) at 1 to 2 volume ratios and subcutaneously injected into the right flank of 5-week-old female BALB/c nu/nu mice (Nippon SLC, Hamamatsu, Japan). Mice were randomly divided into three groups when the tumors grew to approximately 5 to 8 mm in diameter. PBS, OAd5, and OAd35 were intratumorally and intravenously injected into each group at doses of 1×109 VP/mouse (intratumoral administration) and 1×1011 VP/mouse (intravenous administration) on days 0 and 3. The tumor sizes were measured every 3 days using a vernier caliper. The tumor volumes were calculated from the following formula: tumor volume (mm3) = a × b2 × 3.14 × 6−1, where a is the longest dimension, and b is the shortest (39). The in vivo animal experiments were approved by the Animal Experiment Committee of Osaka University.
Statistical analyses. The one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparisons test, and the two-way ANOVA with Bonferroni’s multiple comparisons test were performed using GraphPad Prism (GraphPad Software, San Diego, CA, USA). Data are presented as means±SD or SE.
Results
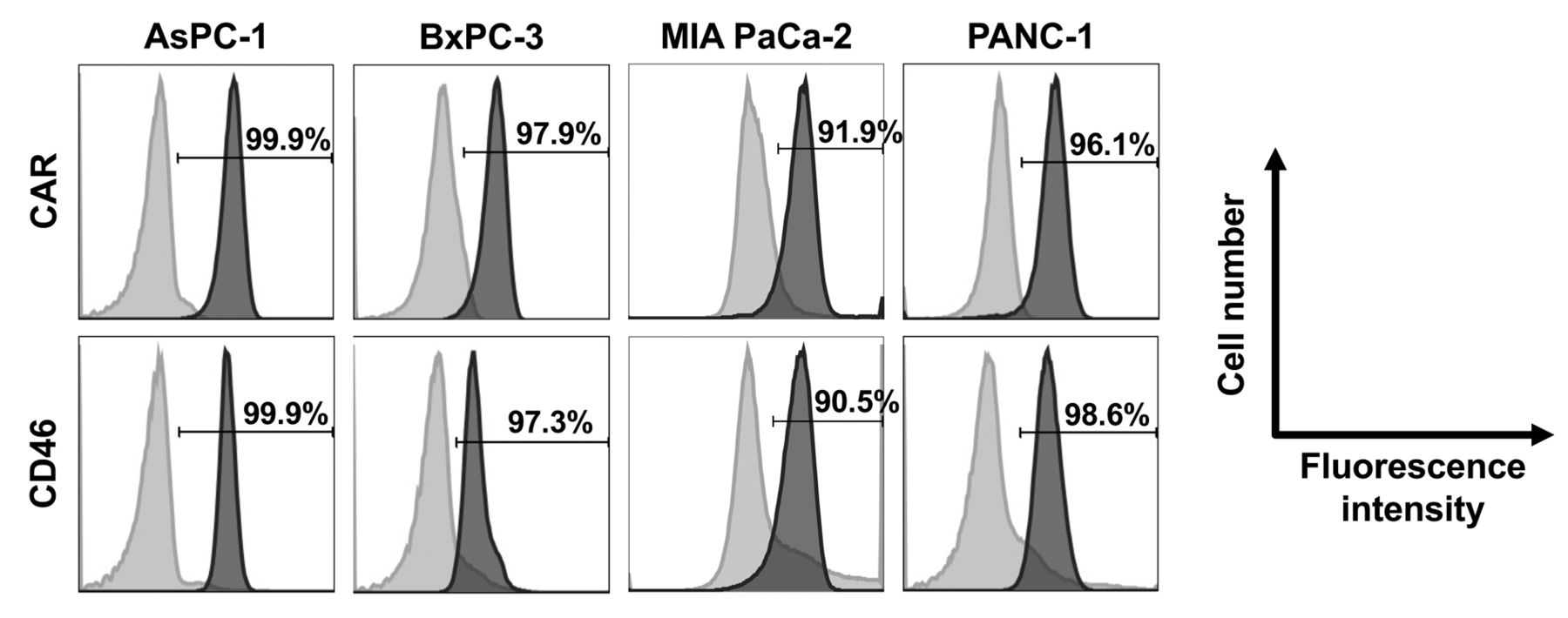
Expression of CAR and CD46 on human pancreatic cancer cell lines. Flow cytometric analysis was performed to examine the expression levels of the infection receptors for OAd5 and OAd35, i.e., CAR and CD46, respectively, on the human pancreatic cancer cell lines (Figure 1). More than 90% of all pancreatic cancer cell lines examined efficiently expressed CAR and CD46 on their cell surfaces. These data suggested that the four human pancreatic cancer cell lines used in the present study were susceptible to both OAd5 and OAd35.

The expression levels of CAR and CD46 on human pancreatic cancer cell lines. AsPC-1, BxPC-3, MIA PaCa-2, and PANC-1 cells were labeled with mouse anti-CAR monoclonal antibody and PE-conjugated goat anti-mouse IgG secondary antibody or PE-conjugated mouse anti-CD46 monoclonal antibody. The Y-axis indicates cell numbers (% of max); the X-axis indicates fluorescence intensity. Light gray histogram: isotype control; gray histogram: anti-CAR or anti-CD46 antibody.
In vitro tumor cell lysis activities of OAd35. To examine the in vitro tumor cell lysis activities of OAds, cell viabilities were determined by crystal violet staining following a 5-day incubation after virus infection (Figure 2A). In AsPC-1 and BxPC-3 cells, OAd35 showed more efficient tumor cell lysis activities than OAd5. Although OAd5 did not fully lyse BxPC-3 cells even at 1,000 VP/cell, almost all tumor cells were lysed by OAd35 at 10 VP/cell in BxPC-3 cells. OAd5-F35 also showed higher levels of tumor cell killing in BxPC-3 cells than OAd5, but less efficiently killed BxPC-3 cells compared with OAd35 (Figure 2B). OAd35 killed almost all tumor cells at more than 300 VP/cell in MIA PaCa-2 and PANC-1 cells, although OAd5 mediated more efficient tumor cell lysis activities than OAd35.

In vitro tumor cell lysis activities of OAd35. (A) The cell viabilities of human pancreatic cancer cell lines were assessed by crystal violet staining assay. AsPC-1, BxPC-3, MIA PaCa-2, and PANC-1 cells were infected with OAd5 or OAd35 at the indicated VPs/cell. After a 5-day incubation, tumor cells were stained with crystal violet. (B) BxPC-3 cells were infected with OAd5, OAd5-F35 or OAd35 at the indicated VPs/cell. After a 5-day incubation, tumor cells were stained with crystal violet. (C) Viabilities of pancreatic cancer cells were evaluated by WST-8 assay. AsPC-1, BxPC-3, MIA PaCa-2, and PANC-1 cells were infected with OAd5 or OAd35 at 500 VP/cell. Cell viabilities were determined on days 1, 3, and 5. The viability in the mock-infected group was normalized to 100%. The dotted line with a circle is OAd5; the solid line with a square is OAd35. These data are expressed as the means±SD (n=4).
The tumor cell lysis activities of OAds were also assessed by WST-8 assay (Figure 2C). More than 95% of the pancreatic cancer cells were efficiently killed following a 5-day incubation after OAd35 infection. More than 60% of BxPC-3 cells survived after a 5-day incubation with OAd5. In contrast, the viability of BxPC-3 cells was 41% on day 1 following OAd35 infection. In MIA PaCa-2 and PANC-1 cells, OAd35 achieved less than 10% tumor cell viabilities on day 5, although OAd5 more efficiently killed MIA PaCa-2 and PANC-1 cells than OAd35. These data demonstrated that OAd35 exhibited efficient tumor cell lysis activities in four types of human pancreatic cancer cell lines, although the tumor cell lysis activity levels of OAd35 differed among the four cell lines.
Effects of pre-existing anti-Ad neutralizing antibodies on in vitro tumor cell lysis activities of OAd35. Next, to examine the impacts of pre-existing anti-Ad neutralizing antibodies on the in vitro tumor cell lysis activities of OAds, OAds were pre-incubated with human serum before infection of MIA PaCa-2 (Figure 3A) and PANC-1 cells (Figure 3B). The viabilities of OAd5-infected tumor cells were largely elevated in the presence of all human serum batches tested. On the other hand, OAd35 mediated efficient tumor cell lysis even in the presence of human serum. The tumor cell viabilities were comparable following infection with OAd35 in the presence and absence of all human serum samples examined. These results indicated that OAd35 mediated more efficient tumor cell lysis activities than OAd5 in the presence of human serum.

In vitro tumor cell lysis activities of OAd35 in the presence of human serum. Viabilities of (A) MIA PaCa-2 and (B) PANC-1 cells were assessed by WST-8 assay. OAd5 or OAd35 were pre-incubated with human serum (#1, #2, #3), followed by addition to the cells at 500 VP/cell. The dilution ratios of human serum were 1 to 250, 1 to 50, and 1 to 10. Cell viabilities were measured following a 5-day incubation after virus infection. The viability in the mock-infected group was normalized to 100%. The data were statistically analyzed using one-way-ANOVA with Dunnett’s post hoc test. *p<0.05, ***p<0.001, n.s.: not significant; p>0.05. [vs. Human serum (−)]. These data are expressed as the means±SD (n=4).
Replication of the OAd35 genome in pancreatic cancer cells. To examine the OAd35 genome replication levels in the pancreatic cancer cells, total DNA containing the OAd genome was isolated from the tumor cells following 24 and 72 h infection with OAds (Figure 4). In AsPC-1 and BxPC-3 cells, although OAd35 showed higher in vitro tumor cell lysis activities than OAd5 (Figure 2), the genome copy numbers of OAd35 were comparable to those of OAd5 at both time points. In MIA PaCa-2 and PANC-1 cells, which OAd5 more effectively killed than OAd35 (Figure 2), the OAd5 genome copy numbers were more than 10-fold higher than the OAd35 genome copy numbers at 24 and 72 h after infection. However, the OAd35 genome copy numbers were more efficiently increased than the OAd5 genome copy numbers from 24 to 72 h in MIA PaCa-2 and PANC-1 cells. The genome copy numbers of OAd35 and OAd5 increased by 319-fold and 9.6-fold, respectively, from 24 to 72 h in MIA PaCa-2 cells. These data indicated that the OAd35 genome was efficiently replicated in pancreatic cancer cells.
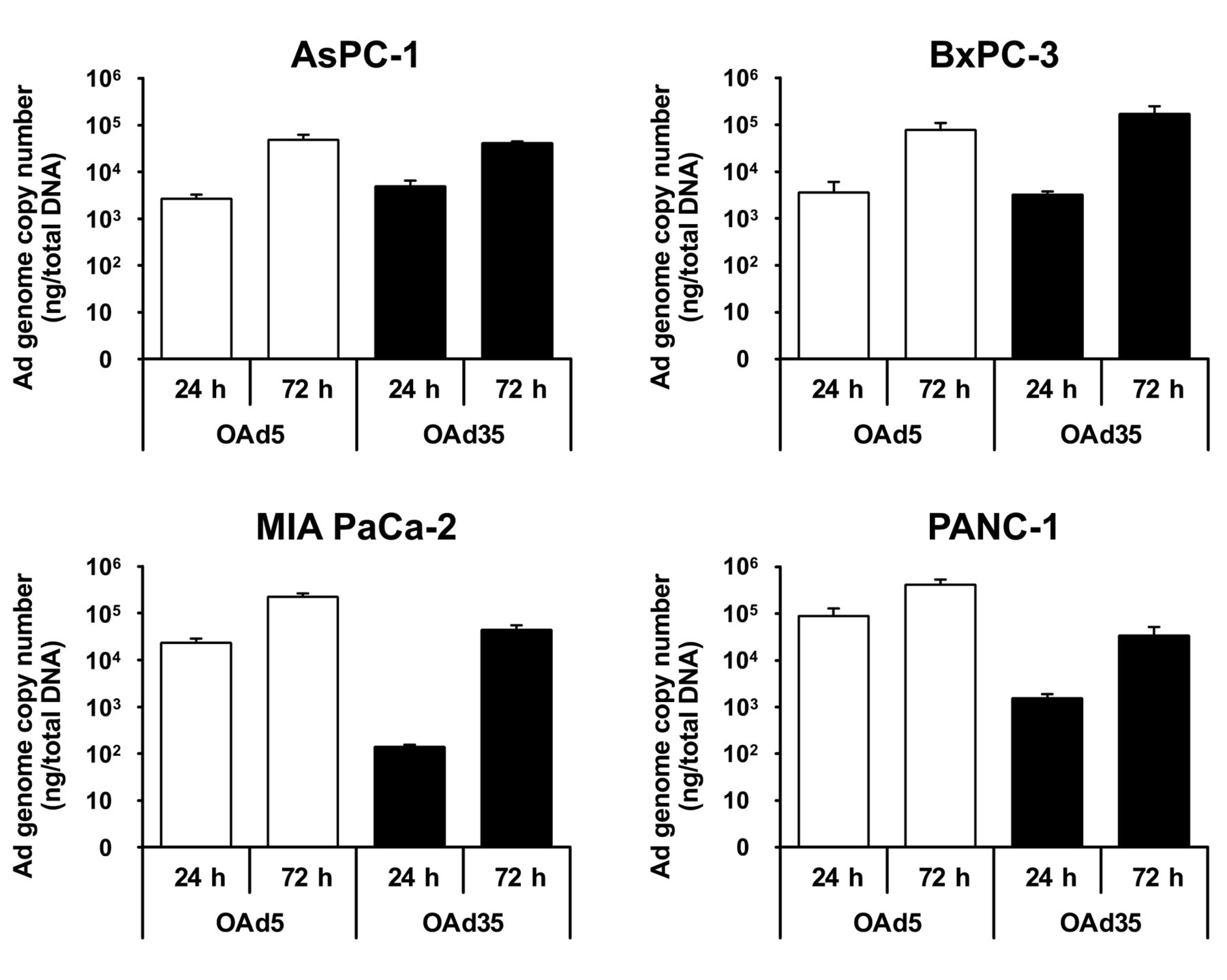
Replication of the OAd35 genome in pancreatic cancer cells. AsPC-1, BxPC-3, MIA PaCa-2, and PANC-1 cells were infected with OAd5 or OAd35 at 100 VP/cell. Total DNA was recovered at 24 h and 72 h after infection, followed by real-time PCR analysis of viral genome copy numbers. These data are expressed as the means±SD (n=3).
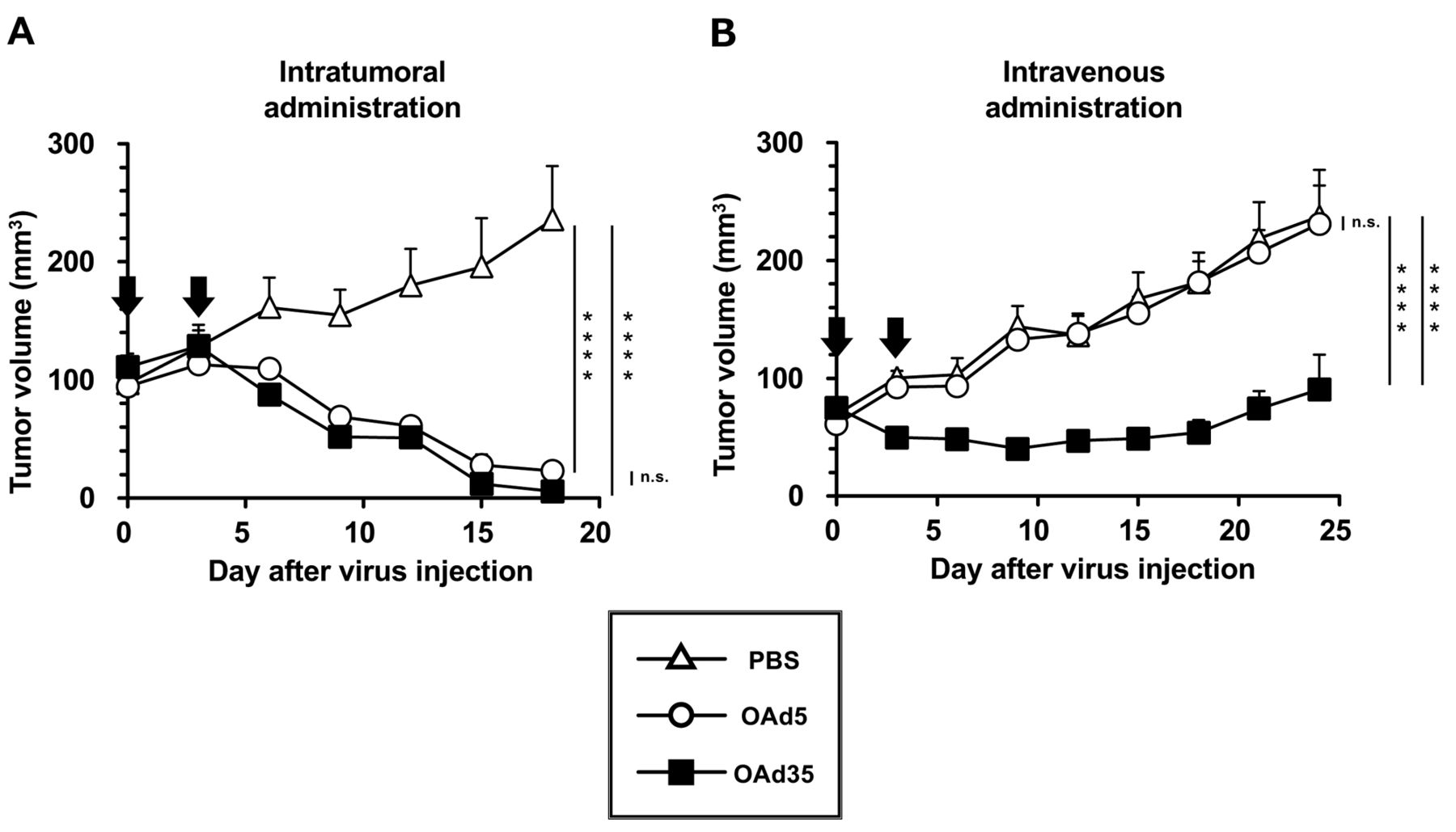
In vivo antitumor effects of locally and systemically administered OAd35. Next, to examine the in vivo antitumor effects of OAds, OAds were intratumorally and intravenously administered to subcutaneous BxPC-3 tumor-bearing mice. Intratumoral administration of OAd5 and OAd35 significantly suppressed the growth of the subcutaneous BxPC-3 tumors (Figure 5A). There were no statistically significant differences in the tumor growth suppression levels of OAd5 and OAd35 following intratumoral administration. Intravenous administration of OAd35 resulted in significant suppression of the growth of subcutaneous BxPC-3 tumors (Figure 5B). However, OAd5 did not appear to suppress tumor growth following intravenous administration. These results indicated that OAd35 mediated efficient tumor growth suppression in pancreatic cancers via local and systemic administration.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
In vivo antitumor effects of intratumorally and intravenously administered OAd35. OAd5 or OAd35 were (A) intratumorally and (B) intravenously injected into BxPC-3 tumor-bearing mice at a dose of 1.0×109 and 1.0×1011/mouse, respectively. Arrows indicate the day points of OAd injection (days 0 and 3). The data were statistically analyzed by two-way ANOVA with Bonferroni’s multiple comparisons post hoc test. ****p<0.0001, n.s.: not significant; p>0.05. Tumor volume is expressed as the mean tumor volume±SE (N=5-6).
Discussion
This study examined the therapeutic efficacy of OAd35 on pancreatic cancers. Despite the advancements in preventive diagnosis, chemotherapy, and combination therapy for pancreatic cancer therapy, the five-year survival rate is still among the lowest of all cancers over the previous decade. Various types of OVs have been tested as potential novel therapies for pancreatic cancers (40, 41). However, there are several problems in OV-mediated pancreatic cancer therapy, such as insufficient tumor cell lysis activities and poor access to distant metastatic tumor cells. In this study, OAd35 efficiently killed human pancreatic cancer cells even in the presence of human serum, suggesting that OAd35 is a promising remedy for pancreatic cancers (Figure 2 and Figure 3). Pancreatic cancers are an ideal target for OAd35 because pancreatic cancer cells highly express CD46 (Figure 1) (30, 33), which is a primary receptor for OAd35. Pancreatic cancers in the clinical samples have also been shown to highly express CD46 (31, 32).
In this study, not only intratumoral administration but also intravenous administration of OAd35 resulted in efficient regression on subcutaneous BxPC-3 tumors. In the case of OAd5, however, statistically significant growth suppression of BxPC-3 tumors was not observed following intravenous administration, despite the efficient tumor growth suppression of OAd5 following intratumoral administration (Figure 5). Since Ad5 possesses high hepatotropism, conventionally OAd5 has been administered mainly via local injection in preclinical and clinical settings (16, 42-44). More than 90% of the injected dose of replication-incompetent Ad5 vector accumulated in the liver following intravenous administration in mice (45). An intravenously administered Ad35 vector was also found to accumulate in the liver, but most of the hepatically accumulated Ad35 vector was taken up by Kupffer cells (16). OAd35, which escaped from the uptake by liver Kupffer cells, might be distributed to the subcutaneous tumors. Alternatively, OAd35 might activate innate immunity following intravenous administration, resulting in immune-mediated antitumor effects. Previous studies demonstrated that Ad35 activated innate immunity in immune cells, including dendritic cells (46, 47). Since distant metastasis is involved in pancreatic cancer-related death, the finding that systemically administered OAd35 mediates efficient antitumor effects holds promise for the treatment of pancreatic cancer.
A fiber-substituted OAd5 containing the Ad35 fiber protein, OAd5-F35, mediated higher levels of tumor cell killing in BxPC-3 cells than OAd5, but killed BxPC-3 cells less efficiently than OAd35 (Figure 2B). These results indicated that other factors except for CD46 expression levels on BxPC-3 cells led to the difference in tumor cell lysis activities of OAd35 and OAd5-F35 in BxPC-3 cells. Intracellular trafficking pathways and subsequent cellular responses might differ between OAd35 and OAd5-F35. Previously, we demonstrated that innate immune responses following transduction with a replication-incompetent Ad35 vector and a fiber-substituted Ad5 vector containing the Ad35 fiber protein were different, although both Ad vectors showed comparable levels of transduction efficacies in mouse dendritic cells (48).
OAd35 killed AsPC-1 and BxPC-3 cells more efficiently than conventional OAd5 (Figure 2), although the genome copy numbers of OAd35 in these cells at 24 and 72 h following infection were comparable to those of OAd5 (Figure 5). These results suggested that the mechanism of tumor cell killing following infection would be somewhat different between OAd5 and OAd35. However, it remains unclear why OAd35 showed higher levels of tumor cell killing in AsPC-1 and BxPC-3 cells than OAd5. Further examination is necessary to elucidate the mechanism underlying the efficient tumor cell killing activity of OAd35 in pancreatic tumor cells.
In conclusion, OAd35 showed more efficient in vitro tumor cell lysis activities against four pancreatic cancer cell lines than conventional OAd5. Pre-incubation with human serum significantly attenuated the tumor cell lysis activities of OAd5. On the other hand, OAd35 efficiently lysed the tumor cells in the presence and absence of human serum. While systemic administration of OAd5 did not show significant tumor growth suppression, OAd35 significantly suppressed subcutaneous BxPC-3 tumor growth. These results suggested that OAd35 would be suitable as an alternative anticancer agent for pancreatic cancers.
Acknowledgements
The Authors thank Toshiyoshi Fujiwara (Okayama University Graduate School of Medicine, Dentistry and Pharmaceutical Sciences, Okayama, Japan) and Kazunori Aoki (National Cancer Center Research Institute, Tokyo, Japan) for their support. This study was supported by grants-in-aid for Scientific Research (A) (20H00664) from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan and the Platform Project for Supporting Drug Discovery and Life Science Research [Basis for Supporting Innovative Drug Discovery and Life Science Research (BINDS)] from the Japanese Agency for Medical Research and Development (AMED) under grant numbers JP22ama121052 and JP22ama121054, and Grant from Oncolys Biopharma, Inc. (Tokyo, Japan). R. Ono is a Research Fellow of the Japan Society for the Promotion of Science (22J13377).
Footnotes
Authors’ Contributions
R. Ono and F. S. conceptualized the experiments. R. Ono, K. T., S.T., and R. Onishi performed the experiments and analyzed the data. R. Ono and F. S. wrote the manuscript, and H. M. reviewed and edited the manuscript. F. S. and H.M. supervised the project. R. Ono, F. S., and H. M. acquired the funding.
Conflicts of Interest
F.S. and H.M. have the potential to receive patent royalties from Oncolys BioPharm, Inc. The other Authors declare no conflicts of interest in relation to this study.
- Received November 20, 2022.
- Revision received December 5, 2022.
- Accepted December 6, 2022.
- Copyright © 2023 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).