

Abstract
Background/Aim: To determine if long-chain non-coding RNA (lncRNA) MIR4435-2HG (MIR4435) expression is associated with pre-malignant colon polyps and colon cancer. Materials and Methods: Children’s colonic-polyp specimens were sequenced for MIR4435 expression. LncRNA MIR4435 expression data in colorectal cancer and normal intestinal tissues were retrieved from The Cancer Genome Atlas (TCGA). The proliferation, adhesion, and invasion ability of human colon-cancer cell line HCT116 with or without MIR4435 knockdown was analyzed. The expression of Smad4, desmoplakin, and β-catenin genes was detected by western blotting in HCT116 cells. Results: MIR4435 expression correlated with the size of intestinal polyps in children. Expression of MIR4435 was up-regulated in colorectal cancer. MIR4435 knockdown in HCT116 cells inhibited their proliferation, adhesion, and invasion ability. Smad4 and desmoplakin were up-regulated and β-catenin was down-regulated in HCT116 cells by MIR4435 knockdown. Conclusion: MIR4435 expression correlated with the size of intestinal polyps in children and with the proliferation, adhesion, and invasion ability of colon-cancer cells and was upregulated in colon cancer.
- Long chain non-coding RNA
- MIR4435-2HG
- colon polyps
- colon cancer
Long-chain non-coding RNAs (lncRNAs) are more than 200 nucleotides long (1). LncRNAs are involved in chromatin remodeling, gene transcription, protein translation and stability (2, 3). Recently, studies have found that abnormal expression or function of lncRNA is associated with the occurrence and development of many major human diseases, such as cancer (4), kidney disease (5), cardiovascular diseases (6), inflammatory bowel disease (IBD) (7) and diabetes (8).
LncRNA MIR4435-2HG (MIR4435) (also known as lncRNA-AWPPH, MIR4435-1HG, linc00978, MORRBID and AGD2) was originally recognized as a predictive biomarker for lung cancer and colon cancer (9, 10). It was also recently found to act as an oncogene in some cancers such as prostate cancer (9-13).
Juvenile polyps are the most common histologic subtype of polyps in pediatrics. Although pediatric intestinal polyps are generally considered benign, they have also been reported to lead to malignant transformation (14). Some colorectal adenomas and adenocarcinomas in adulthood have been shown to originate from juvenile polyps (15, 16). Based on the relationship between children’s intestinal polyps and adult colon cancer, as well as the respective pathological characteristics of the two diseases, in the present report, we investigated role of MIR4435 in juvenile intestinal polyps, colon cancer and colon-cancer cells.
Materials and Methods
Patient specimens and cell culture. Sixty-five colonic polyps specimens were obtained from patients at the Second Affiliated Hospital of Nanjing Medical University with confirmatory diagnoses based on histopathology. Informed written consent prior to operation was obtained from the parents of the children. The human colon-cancer cell line HCT116 was purchased from the Shanghai Cell Bank of Chinese Academy of Sciences (Shanghai, PR China). HCT116 cells were cultured in RPMI 1640 medium (GIBCO, Grand Island, NY, USA) containing 10% fetal bovine serum (FBS) (Lifesciences, Shanghai, PR China), one hundred U/ml penicillin (Solarbio, Beijing, PR China) and 100 μg/ml streptomycin (Yuanye Bio-Technology, Shanghai, PR China), and uncubated at 37°C with 5% CO2. The study was approved by the Ethics Committee of The Second Affiliated Hospital of Nanjing Medical University.
LncRNA sequencing. The sequencing of the patient’s lncRNA specimens was done by Shanghai Major Biological Co. The screening criteria for lncRNAs were: 1) differential expression >2, 2) p-value ≤0.05, 3) lncRNAs with known gene sequences, 4) target genes predicted to be cis-regulated by the of Majorbio Cloud Platform.
Data collection & processing. The data containing expression of MIR4435 in 570 colorectal cancer and 42 normal intestinal mucosal tissues were retrieved from TCGA (https://portal.gdc.cancer.gov). Gene-expression profiling and interactive analyses (GEPIA, http://gepia.cancer-pku.cn) were used for analyzing RNA sequencing data of tumors and normal tissues.
RNA extraction and quantitative reverse transcription PCR (RT-qPCR). Total RNA was extracted from patient specimens and cells using the TRIzol reagent (Life Technologies). cDNA was synthesized using reverse transcriptase from VazymE using the following conditions: 15 min at 50°C and 2 min at 85°C, before stopping the reaction at 4°C. The PCR reaction conditions were as following: 5 min at 95°C, followed by 5 s at 95°C, and 31 s at 60°C. The primers for MIR4435 and the endogenous control GAPDH were synthesized by Sangon Biotech Co., Ltd (Shanghai). Sequences of primers were as following: MIR4435 forward, 5′-TGGGAATGGAGGGAAATA-3′ and reverse, 5′-CGGGCAAC AGGTAGAGGT-3′; GAPDH forward, 5′-AAGGTGAAGGTC GGAGTCA-3′ and reverse, 5′-AATGAAGGGGTCATTGATG-3′. The relative expression levels of MIR4435 were normalized to the GAPDH endogenous control and expressed as 2-ΔΔCq.
RNA-interference-vector transfection. For silencing of MIR4435 expression, three RNA interference vectors (shMIR4435-1, shMIR4435-2, and shMIR4435-3) targeting the MIR4435 gene were used to transfect HCT116 cells. An empty vector (shCtrl) was used as negative control. The transfection was performed according to standard procedures. Eighteen h post-transfection, the medium was replaced with RPMI 1640 medium containing 10% FBS. Cells were then observed under fluorescence microscopy 48 h after transfection. The transfected cells with the highest green fluorescent protein (GFP) expression from the RNA interference vectors were selected and treated with medium containing 1 μg/ml of puromycin (Solarbio). Stable GFP expression was observed for more than 10 passages. Images were obtained under fluorescence microscopy. Transfection efficiency of the RNA interference vector in HCT116 cells was confirmed based on down-regulation of MIR4435 expression by qPCR.
Cell-proliferation assay. Cell proliferation was measured using the Cell Counting Kit-8 (CCK-8); (Dojindo Molecular Technologies, Inc., Rockville, MO, USA). HCT116 cells in logarithmic growth phase were seeded in 96-well plates with 5×103/200 μl. Cells were cultured at 37°C in 5% CO2. After 24 h, 48 h and 72 h culture, 10 μl of CCK-8 solution was added to each well and the culture was continued for 4 h. The absorbance at 450 nm was measured and the inhibition rate was calculated.
Cell-adhesion assay. A cell adhesion kit (BestBio, Shanghai, PR China) was used for the cell-adhesion assay. One hundred μl of coating solution was added to each well of 96-well plates and incubated overnight at 4°C. The coating solution was then removed, and the plate was placed in a fume hood for a few minutes to complete drying, followed by 3 washes. Next, 5×104 untransfected and transfected cells were plated in the coated wells and incubated at 37°C with 5% CO2 for 60 min. The medium then was removed, and the wells were washed 3 times before adding 10 μl cell-staining solution and incubation at 37°C for 3-4 h. Colorimetric quantification of the dye was performed at 450 nm using an ELISA plate reader.
Wound-healing assay. A total of 1.5×105 untransfected and transfected cells were plated in 12-well plates for 24 h. The cell layers were then scratched with a sterile pipette tip to make a wound in the monolayer. Cell debris were washed out, and fresh medium was replenished. The cells started migrating from the edge of the wound and repopulated the gap area. Cells were then observed for ‘wound closure’ after 24 h under a light microscope. Image J Software (NIH, Bethesda, MD, USA) was used for analysis and quantification of the healing rate. The wound-area closure was measured in three-independent wound sites per group. The wound-healing rate was calculated as follows: (0 h wound area − 24 h wound area)/0 h wound area ×100%.
Cell invasion assay. The upper-chamber surface of the bottom membrane of the transwell was coated with 50 mg/l Matrigel (Solarbio) (1:4 dilution), air-dried at room temperature and placed under UV light for 30 min. Cells from each experimental group, in the logarithmic-growth phase, were inoculated in the chambers with 10,000 cells per well. Serum-free medium (Lonza, Basel, Switzerland) (200 μl) was added to the upper chamber and complete medium (600 μl) containing 20% FBS was added to the lower layer. After 24 h incubation, cells were washed with PBS 2 times, and fixed with ethanol for 20 min. The cells remaining on the upper membrane were removed with cotton wool, whereas the cells that had invaded through the membrane were stained with crystal violet. After staining, cells on the back of the transwell membrane were imaged and counted using an inverted microscope.
Western blotting. Total proteins from tissues or cells were lysed with RIPA lysis buffer (Beyotime, Haimen, Jiangsu, PR China) containing phenylmethanesulfonyl fluoride (Beyotime) and a protease inhibitor cocktail (Beyotime). Proteins (20 μg) were separated by SDS-PAGE and transferred to a PVDF membrane (Millipore, Boston, MA, USA). The membranes were incubated with the diluted primary antibodies specific for desmoplakin, β-catenin, E-cadherin, and Smad (Santa Cruz Biotechnology, Dallas, TX, USA) at 4°C overnight followed by an a horse-radish peroxidase (HRP)-conjugated secondary antibody (Santa Cruz Biotechnology, Shanghai) after blocking with 5% bovine serum albumin (Yuanye Bio-Technology) for 1 h. Anti-GAPDH antibody (Santa Cruz Biotechnology) was used as an internal control. Blots were measured with an ECL chromogenic substrate (Beyotime). Protein-expression levels were measured by densitometry analysis of the developed bands.
Statistical analyses. All data are expressed as mean±standard deviation (SD). The t-test was used to analyze the statistical significance of the two groups. The statistical significance of multiple groups was analyzed by ANOVA followed by Bonferroni correction. p≤0.05 was considered significant.
Results
Correlation of MIR4435 expression with size of intestinal polyps in children. Sixty-five colonic polyp specimens from children were sequenced for MIR4435. The median expression level of MIR4435 in children’s intestinal-polyp tissues was calculated. The association of MIR4435 expression with clinicopathological parameters was analyzed to investigate the clinical relevance of MIR4435 expression in children’s intestinal polyps (Table I). It was found that the expression level was related to the size of intestinal polyps. Polyps greater than 1 cm had significantly higher expression of MIR4435 compared to polyps smaller than 1 cm (p<0.05). There was no significant correlation with the patients’ age, sex, and polyp distribution location (p>0.05).
Correlation between lncRNA MIR4435 expression and clinicopathological characteristics in children’s intestinal polyps.
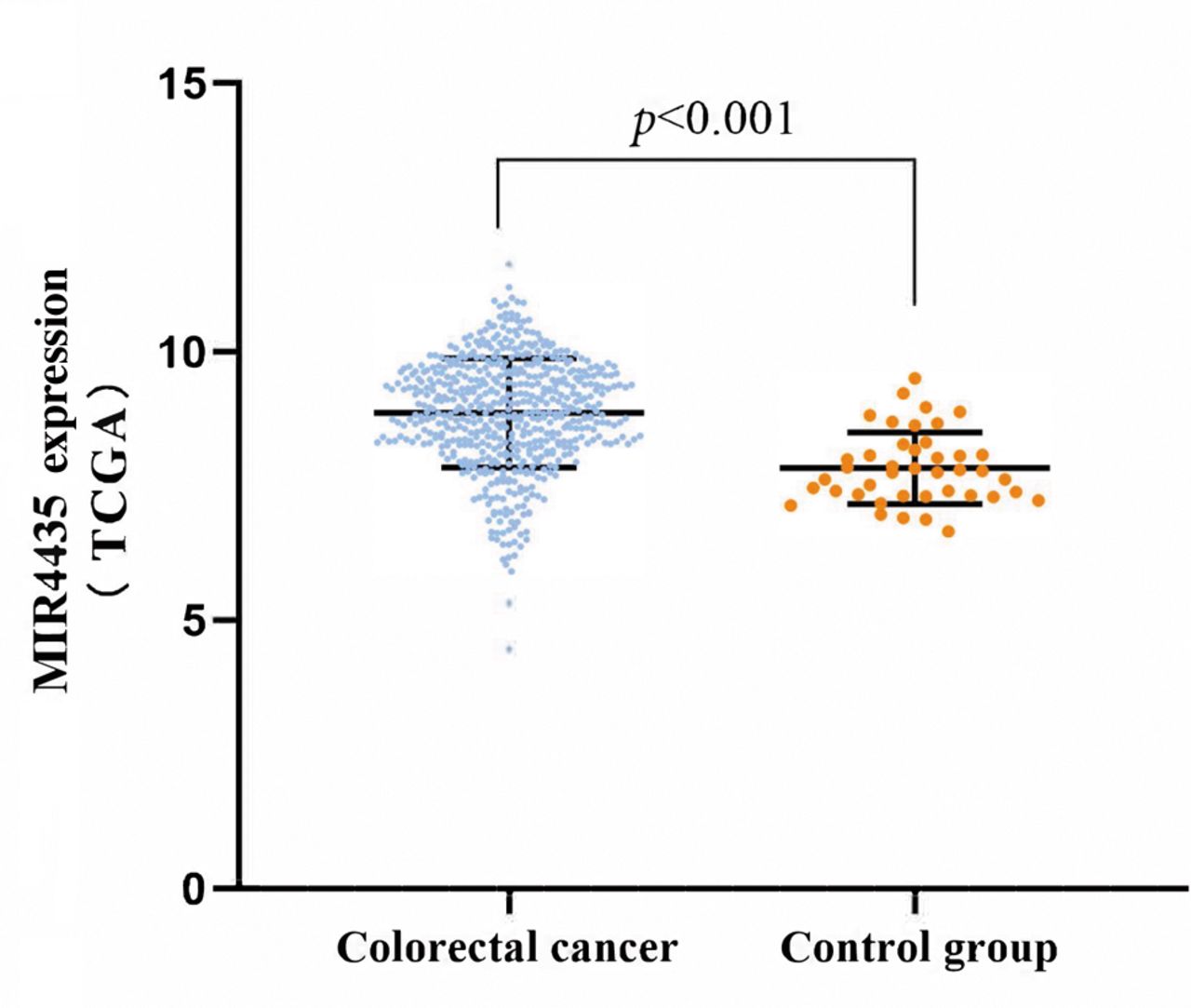
Expression of MIR4435 is up-regulated in colorectal cancer. A data set containing 570 colorectal cancer tissues and 42 normal intestinal mucosal tissues was downloaded from TCGA. The analysis showed that MIR4435-expression levels were significantly up-regulated in colorectal cancer tissues (p<0.001) (Figure 1).

MIR4435 expression in normal intestinal-mucosal and colorectal-cancer tissues. A data set containing 570 colorectal-cancer and 42 normal intestinal-mucosal tissues downloaded from TCGA was analyzed for MIR4435 expression.
Effect of MIR4435 knockdown on proliferation of colon-cancer cells. To investigate the role of MIR4435 in the malignancy of colon cancer, the lncRNA was knocked down in colon cancer cells by RNA interference. As shown in Figure 2, MIR4435 expression was significantly down-regulated by more than 70% in shMIR4435-2 transfected HCT116 cells as compared to shCtrl-vector-control cells (**p<0.01, ***p<0.001).

Knockdown of MIR4435 in colon cancer cells. MIR4435 was knocked down in HCT116 colon-cancer cells by RNA interference. MIR4435 mRNA expression in HCT116 cells and HCT116 cells transfected with each of the 3 shMIR4435 vectors or shCtrl control vector. **p<0.01, ***p<0.001; ns: not significant.
The CCK-8 assay was used to assess the potential effects of MIR4435 silencing on cancer-cell growth. Cell proliferation was monitored for 72 h. As shown in Figure 3, cell proliferation was significantly reduced by MIR4435 silencing in the transfected HCT116-shMIR4435 cells as compared to the untransfected and vector control cells (*p<0.05).

Effect of MIR4435 knockdown on proliferation of colon-cancer cells. Cell proliferation at 24 h, 48 h and 72 h was significantly reduced by MIR4435 silencing in the transfected HCT-116-shMIR4435 cells. *p<0.05; ns: no significance.
Effect of MIR4435 knockdown on adhesion of colon-cancer cells. To understand how MIR4435 knockdown affects cell motility in 2D, we assessed the effect of its knockdown on the adhesion of colon-cancer cells to collagen. Figure 4 reveals that the transfected HCT116-shMIR4435-2HG cells have a lower adhesion rate than the untransfected cells (*p<0.05), suggesting that MIR4435 knockdown decreased their adhesion ability.

Effect of MIR4435 knockdown on adhesion of colon-cancer cells. Cell adhesion was significantly reduced by MIR4435 silencing in the transfected HCT116-shMIR4435 cells. *p<0.05; ns: no significance.
Effect of MIR4435 knockdown on invasion capability of colon-cancer cells. Cell-invasive ability was examined by transwell and wound-healing assays. As shown in Figure 5A, cell migration to the lower chamber in the transwell was significantly decreased by MIR4435 knockdown in the transfected HCT116-shMIR4435 cells, compared to the untransfected and vector-control cells (**p<0.01).

Effect of lncRNA MIR4435 knockdown on invasion and migration capability of colon-cancer cells. (A) The cell-invasive ability of HCT-116 cells was examined by a transwell assay. Cell invasion was significantly reduced by lncRNA MIR4435 silencing in transfected HCT-116-shMIR4435 cells. **p<0.01. (B) Cell migration ability was examined by the wound-healing assay. Cell migration was significantly reduced by MIR4435 silencing in the transfected HCT-116-shMIR4435 cells. **p<0.01; ns: no significance.
Additionally, in the wound-healing assay, the transfected HCT116-shMIR4435 cells healed the wound significantly slower than the untransfected and vector control cells (**p<0.01) (Figure 5B). These results suggest that MIR4435 knockdown may play a role in the migration and invasion abilities of colon cancer cells.
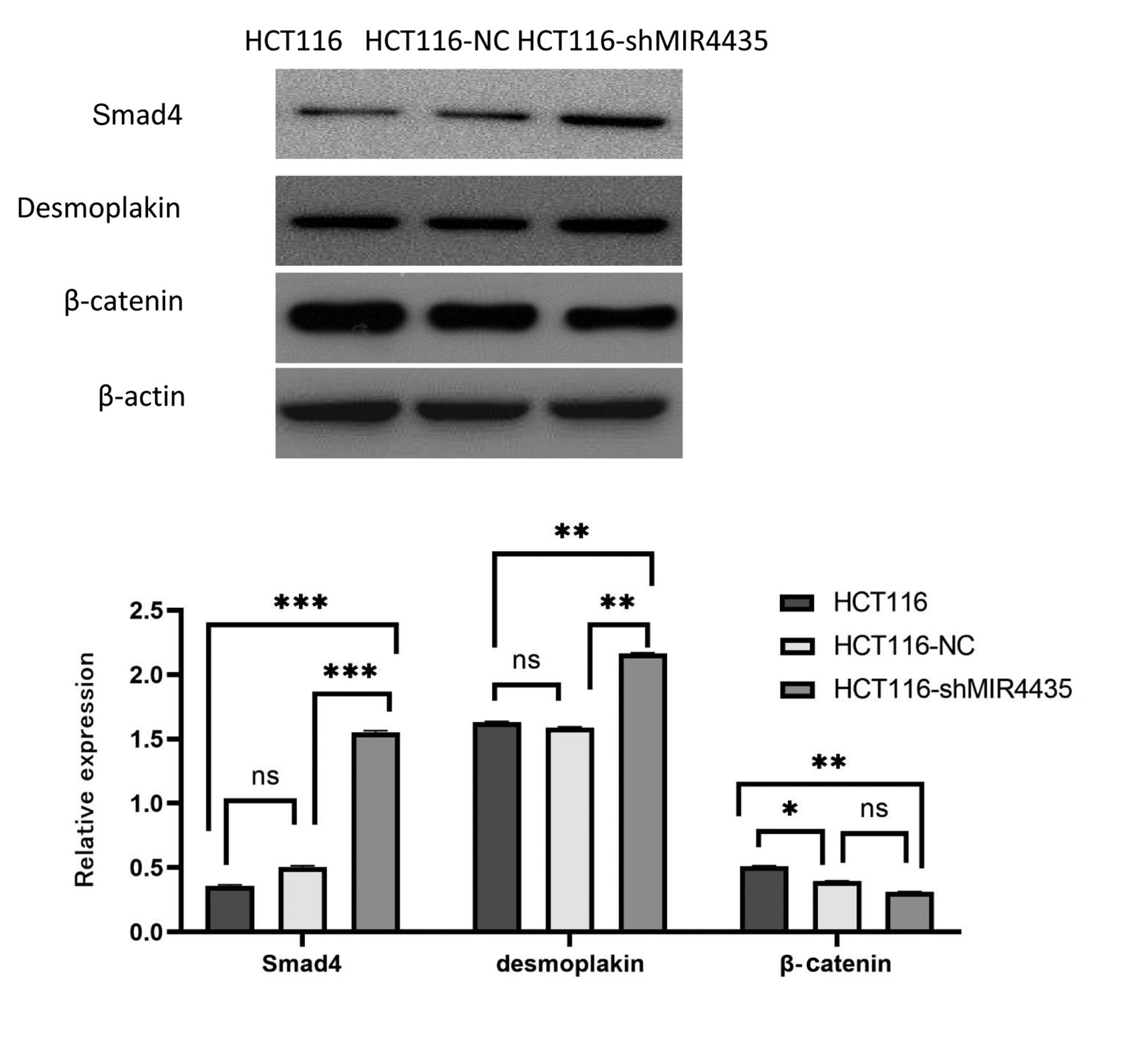
Effect of MIR4435 knockdown on gene expression of colon-cancer cells. To understand the molecular mechanisms underlying the effect of MIR4435 knockdown on the metastasis of colon cancer, Smad4, desmoplakin and β-catenin were analyzed by western blotting. Compared to the untransfected and vector control cells, Smad4 and desmoplakin were up-regulated and β-catenin expression was down-regulated in the transfected HCT116-shMIR4435 cells (*p<0.05, **p<0.01, ***p<0.001) (Figure 6).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of MIR4435 knockdown on gene expression of colon-cancer cells. Smad4 and desmoplakin were up-regulated and β-catenin expression was down-regulated in the transfected HCT-116-shMIR4435 cells. *p<0.05, **p<0.01, ***p<0.001; ns: no significance.
Discussion
Accumulated evidence has indicated that lncRNAs might play important roles in several human diseases (17). The dysregulation of HOTAIR, one of the most well-known lncRNAs, may lead to abnormal expression of gene products related to progression, invasion, and metastasis of many different cancers (18). In addition, the lncRNA MALAT1 is related to the microvascular dysfunction in STZ-induced diabetic rats and type 2 diabetic nephropathy mice (19). Knockout of MALAT1 can affect p38 MAPK signal transduction and interfere with the proliferation of retinal- endothelial cells, suggesting that MALAT1 is a potential target for anti-angiogenic therapy in diabetic retinopathy (20).
However, the association of lncRNA expression with children’s intestinal polyps was poorly understood. In the present study, we first report that expression of MIR4435 is related to the size of children’s intestinal polyps.
MIR4435, which has been reported to have aberrant expression in some malignant tumors, may regulate tumorigenesis and progression (10, 13). We analyzed the data from the TCGA database and found that MIR4435 was upregulated in colorectal cancer compared with normal intestinal mucosa. We then showed that the proliferation, adhesion, migration, and invasion ability in human colon-cancer cell line HCT116 was decreased after MIR4435 knockdown. Some recent reports have found that MIR4435 is associated with colorectal-cancer proliferation, metastasis and cisplatinum resistance, which is in agreement with our results (21, 22). These results suggest that MIR4435 acts as an oncogene in colon cancer, from the colon-polyp stage. MIR4435 may become a new target gene for the treatment of children’s intestinal polyps and colon cancer.
LncRNAs may regulate intracellular signal transmission by interacting with RNA-binding proteins (23, 24). Wang et al. (25) reported that desmoplakin is an MIR4435-interaction protein and negatively correlates with the expression of MIR4435. Desmoplakin and β-catenin are highly homologous and compete with each other to connect with E-cadherin and regulate the Wnt/β-catenin pathway (26). Smad4 is a classic tumor-suppressor protein encoding a common intracellular mediator of the TGF-β superfamily and is mutated or deleted in certain cancers (27, 28).
The present study showed that MIR4435 knockdown increased the expression of Smad4 and desmoplakin and decreased β-catenin expression. Based on the present results, we could speculate that MIR4435 may promote the transformation of intestinal cancer cells, possibly by affecting Wnt/β-catenin signal transduction and decreasing expression of tumor suppressor genes such as Smad4.
Acknowledgements
This work was supported by the Nanjing Municipal Health Science and Technology Development Special Fund (grant number 2018C03022, 2018) and Ili Prefecture Science and Technology Development Fund Project (grant number YZ2021B004, 2021).
Footnotes
Authors’ Contributions
Danrong Zhu, Guozhong Ji, Weisong Sheng and Rong Zhang designed the study. Danrong Zhu, Piao Guo and Xin Yu Jin carried out experiments. Danrong Zhu and Piao Guo analyzed the data and wrote the manuscript. Zhijian Yang and Robert M. Hoffman revised the manuscript.
Conflicts of Interest
None of the Authors have any conflicts of interest in regard to this study.
- Received May 28, 2022.
- Revision received August 7, 2022.
- Accepted August 26, 2022.
- Copyright © 2022 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).