

Abstract
Background/Aim: Reports on over-expression of the epidermal growth factor receptor (EGFR) in bladder cancer and its function in tumorigenesis have suggested to target this antigen. Materials and Methods: We generated the targeted toxin EGF-PE40 consisting of the human epidermal growth factor (EGF) as the binding domain and PE40, a truncated version of Pseudomonas Exotoxin A, as the toxin domain. EGF-PE40 was tested on EGFR-expressing bladder cancer cells in view of binding via flow cytometry, and cytotoxicity via WST viability assay. Induction of apoptosis was examined by western blot. Results: The targeted toxin specifically triggered cytotoxicity in the bladder cancer cells with 50% inhibitory concentration (IC50) values in the low nanomolar or picomolar range, and was about 1,250- to 1,500-fold more cytotoxic than the EGFR inhibitor erlotinib. Cytotoxicity of EGF-PE40 was based on the induction of apoptosis. Conclusion: EGF-PE40 represents a promising candidate for the future treatment of bladder cancer.
- Bladder cancer
- epidermal growth factor
- epidermal growth factor receptor
- targeted toxin
- Pseudomonas Exotoxin A.
Bladder cancer (BC) is the 10th most common cancer worldwide. Approximately 573,000 new cases and 213,000 deaths from this tumor are expected every year, with highest incidence rates in Europe and North America (1). About 70% of BC cases are non-muscle invasive at the time of diagnosis. They are treated by transurethral resection of the tumor (TURBT) followed by adjuvant intravesical treatment with Bacillus Calmette-Guerin (BCG) or mitomycin (2). However, in about 70% of patients the tumor recurs and in 10-20% of these cases it progresses to muscle-invasive BC (3). Only 50% of patients with muscle-invasive BC survive 5 years despite undergoing radical cystectomy (4). For patients with metastatic disease, cisplatin-based chemotherapy is the standard of care, conferring a median overall survival of 13-16 months (5, 6). Despite therapeutic progress in recent years, e.g. with immune checkpoint inhibitors (7, 8), BC is still characterized by a high recurrence rate and a complex and expensive treatment with numerous local and systemic side effects (9). Therefore, new targeted treatment options are urgently needed.
In recent years, EGFR has aroused great interest as new target for the treatment of BC. EGFR was mainly detected in the membrane of BC cells and is only sparsely distributed in the cytoplasm (10). EFGR expression was detected in 55-58% of bladder transitional cell carcinoma samples (10, 11) and in 75% of invasive BC tissues (12). In contrast, only 10% of non-cancer bladder mucous membrane tissues samples were found to be EGFR positive (10).
EGFR signaling has been shown to regulate cell proliferation, apoptosis, angiogenesis, invasion, and tumor metastasis in preclinical models of transitional cell carcinoma (TCC) of the bladder (13). EGFR expression was positively correlated with clinical stage, pathologic grade, and recurrence of BC, and negatively correlated with prognosis and survival (10, 14).
Targeting EGFR with the monoclonal antibody Cetuximab, however, yielded disappointing results in patients with BC, failed to prevent disease progression and developed drug resistance and toxicities (15, 16). Preliminary data from a phase II clinical trial examining the effectiveness of neoadjuvant EGFR inhibition in muscle-invasive BC (clinical stage T2) by the inhibitor erlotinib suggested that it might have single-agent activity (17).
Targeted toxins might represent alternatives in the immunotherapy of BC. They consist of a ligand as binding domain and a toxin of bacterial, fungal, plant, or animal origin as active domain (18). In general, the ligand domain binds to target antigens on the surface of cancer cells and, after internalization, the toxin domain can unleash its enzymatic activity and trigger cancer cell death (19). One prominent toxin for the construction of targeted toxins is Pseudomonas Exotoxin A (PE). PE is a virulence factor from the bacterium Pseudomonas aeruginosa (20). PE is a 638 aa protein with several functional domains. Domain Ia (aa 1-252) binds to CD91, a receptor, which is omnipresent on the surface of eukaryotic cells. Domain II (aa 253-364) includes a furin cleavable motif and enables the toxin to cross cellular membranes. Domains Ib (aa 365-404) and III (aa 405-613) have ADP-ribosyl transferase activity and a KDEL-like motif at the C-terminus. PE binds to CD91 on host cells and is internalized into endosomes, where it is cleaved by furin. The C-terminal part of the protein, consisting of domain II, Ib, and III is 40 kDa in size and is therefore also called PE40. PE40 can bind via its KDEL-motif to KDEL-receptors of the Golgi apparatus. It is then transported retrogradely to the ER and secreted into the cytosol. On the ribosomes, it specifically ADP-ribosylates the eukaryotic elongation factor 2 (eEF-2). ADP-ribosylation of eEF-2 leads to an inhibition of protein biosynthesis and to apoptosis of the target cells [rev. in (21)].
In the present study, we generated and analyzed the new targeted toxin EGF-PE40, consisting of the human EGF ligand as the binding domain and PE40 as the toxin domain, in view of binding, cytotoxicity, and induction of apoptosis on EGFR expressing BC cells.
Materials and Methods
Cell lines and reagents. Human urinary BC cell lines T24 (transitional cell carcinoma) and HT-1376 (grade 3, carcinoma) were purchased from the American Type Culture Collection (Manassas, VA, USA) and cultured in Dulbecco’s modified Eagle medium (Gibco, Invitrogen, Karlsruhe, Germany) with 10% fetal calf serum (Biochrom, Berlin, Germany) and penicillin/streptomycin (100 U/ml, 100 mg/l) at 37˚C and 5% CO2. Verification of the cell lines was performed by short tandem repeat analysis (CLS GmbH, Eppelheim, Germany). CHO cell line was cultured in Ham’s F12 nutrient mix medium (Gibco, Invitrogen) with 10% fetal calf serum (Biochrom) and penicillin/streptomycin (100 U/ml, 100 mg/l) at 37˚C and 5% CO2 as control. Erlotinib HCl (Selleck Chemicals Llc, Houston, TX, USA) was dissolved as 4 mg/ml stock solution in DMSO and stored at –80˚C until use.
Generation and purification of EGF and EGF-PE40. The ligand EGF and the targeted toxin EGF-PE40 were cloned into the expression vector pHOG21, periplasmatically expressed in E. coli XL-1 blue bacteria, and purified by immobilized affinity chromatography (IMAC) as described before (22). Purified EGF and EGFR were detected by SDS-PAGE and protein content was determined by Nanodrop analysis (Thermo Fisher Scientific, Waltham, MA, USA).
Western blot analyses. To determine EGFR expression in the target cells, cells were lysed in RIPA buffer as described previously (22) and western blot analysis was done on whole cell lysates (50 μg/lane) using EGFR rabbit polyclonal antibody (pAb) (Santa Cruz Biotechnology, Dallas, TX, USA) and HRP-labeled goat anti-rabbit pAb (Dako Denmark A/S, Glostrup, Denmark). β-Actin was used as a loading control and detected by HRP-conjugated β-Actin rabbit pAb (Cell Signaling Technology Europe, Leiden, the Netherlands). Expression of EGF and EGF-PE40, containing a c-myc tag was verified by western blot using the HRP-labeled anti-cmyc antibody (Roche laboratories) or an anti-PE rabbit pAb (Sigma) and HRP-labeled goat anti-rabbit pAb (Dako Denmark A/S, Glostrup, Denmark). To detect apoptosis, cell lysates before and after 48 h incubation with 2 nM EGF-PE40 were blotted and poly-ADP-ribose-polymerase (PARP) cleavage was detected by rabbit anti-PARP pAb (Cell Signaling Technology Europe) and HRP-labeled goat anti-rabbit pAb (Dako). Visualization of the protein bands was carried out after developing the membranes with an enhanced chemiluminescence (ECL) system (Clarity™ Western ECL Substrate, Bio-Rad Laboratories, Inc., Hercules, CA, USA), using the ChemiDoc™ MP Imaging System and the software Image Lab™.
Flow cytometry. Binding of EGF and EGF-PE40 on the target cells was examined by flow cytometry as described before (23). Bound EGF and targeted toxins were detected via rabbit anti-His-tag mAb (Cell Signaling Technology Europe, Leiden, The Netherlands) and goat anti-rabbit IgG-PE (Southern Biotech, Birmingham, AL, USA). Stained cells were analyzed with a FACS Calibur flow cytometer and the software CellQuest Pro (BD Biosciences, Heidelberg, Germany).
WST-1 cell viability assay. Cell viability after targeted toxin or erlotinib treatment was quantified by the colorimetric WST-1-Assay (Roche Diagnostics, Mannheim, Germany) according to the standard protocol. 50% inhibitory concentrations (IC50) values of WST-1 data were calculated for each cell line using GraphPad Prism 7 software by non-linear regression [(inhibitor) vs. normalized response].
Results
Firstly, expression of the 170 kDa protein EGFR on the BC cell lines T24 and HT-1376 was verified by western blot analysis. The control cell line CHO was found to be human EGFR negative (Figure 1A). After periplasmatic expression in E. coli XL-1 blue bacteria and purification via IMAC, the ligand EGF (8.4 kDa) and the targeted toxin EGF-PE40 (49.2 kDa) were obtained in high purity of >90% as determined by SDS-PAGE (Figure 1B). Expression of the proteins was verified by western blot analysis (Figure 1C and D). Binding of EGF and EGF-PE40 was demonstrated by flow cytometry on T24 and HT-1376 cells. No binding with both constructs was seen on EGFR-negative CHO cells (Figure 1E). This proved that addition of the PE40 domain did not impair the specificity of EGF.
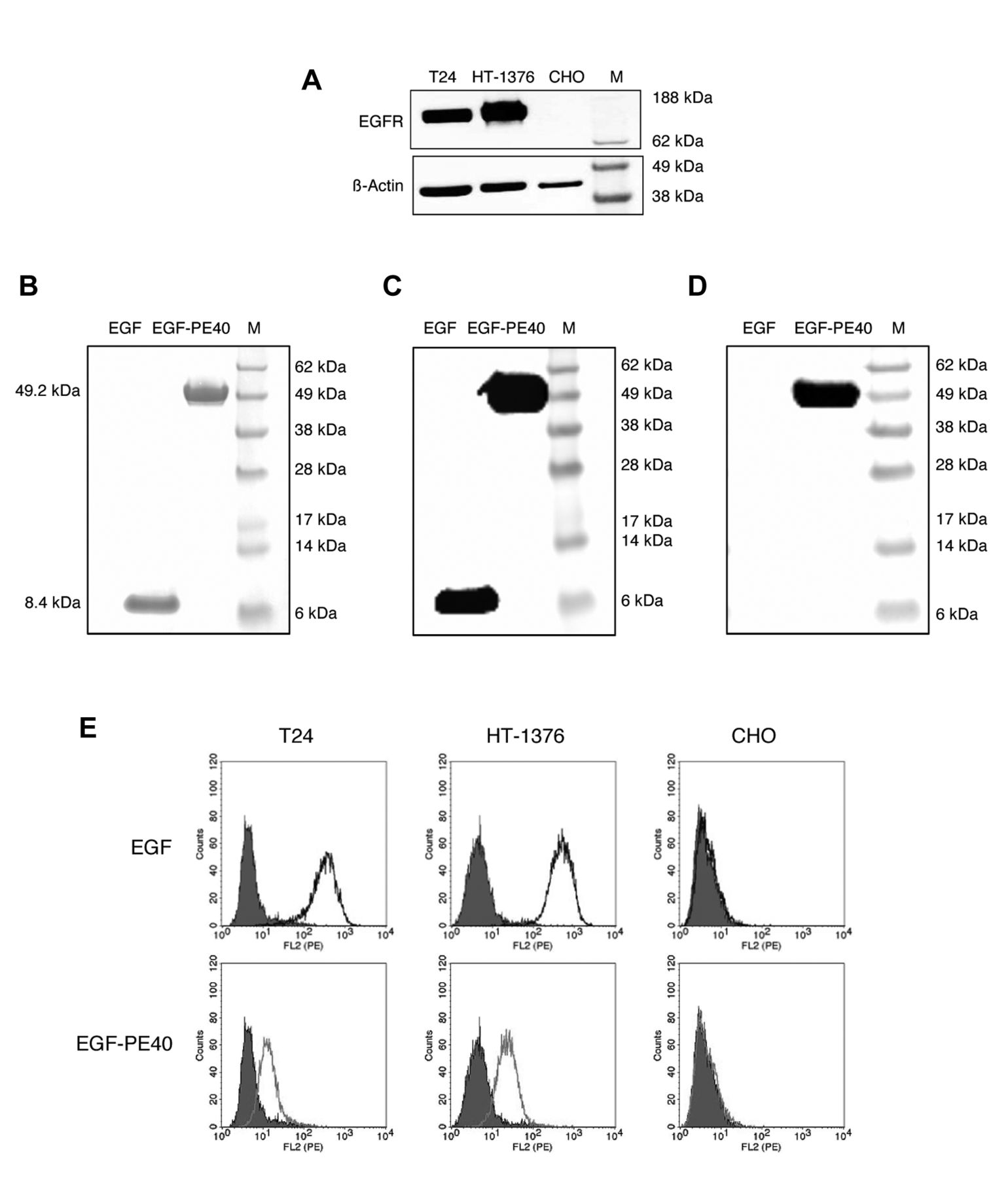
Generation and characterization of the EGF ligand and the anti-EGFR targeted toxin EGF-PE40 on bladder cancer (BC) cells. (A) EGFR expression in BC cells as shown by western blot analysis. EGFR negative CHO cells served as control. (B) SDS-PAGE of the ligand EGF and the targeted toxin EGF-PE40 after periplasmatic expression in E. coli and purification via IMAC. (C) Western blot analysis of EGF and EGF-PE40 using detection antibodies against the c-myc tag or (D) against the PE40 domain. (E) Binding of EGF and EGF-PE40 to EGFR-positive T24 and HT-1376 BC cells as analysed by flow cytometry with secondary detection antibody alone in grey. CHO cells served as controls.
The targeted toxin elicited a high and specific cytotoxicity in T24 and HT-1376 cells with IC50 values of 2.009 nM and 0.281 nM after 48 h, respectively. In CHO cells, no cytotoxicity was detected up to a maximal targeted toxin concentration of 10 nM (Figure 2A). PARP cleavage demonstrated that the targeted toxin induced apoptosis in the BC cells (Figure 2B). With the EGFR inhibitor erlotinib, IC50 values of >2.5 μM and 0.42 μM were calculated on T24 and HT-1376 cells, respectively (Figure 2C). Thus, EGF-PE40 showed an about 1,250 to 1,500-fold enhanced cytotoxicity compared to erlotinib.

{kind=link}
{kind=link}
Cytotoxicity of EGF-PE40 and the EGFR inhibitor erlotinib in bladder cancer cells. (A) Cytotoxicity of EGF-PE40 in T24 and HT-1376 bladder cancer (BC) cells as measured by WST-1 tests. CHO cells were used as control. (B) Induction of apoptosis, marked by cleavage of poly-ADP-Ribose-Polymerase (PARP), in BC cells after incubation with 2 nM EGF-PE40 as shown by western blot analysis. (C) Cytotoxicity of the EGFR inhibitor erlotinib on T24 and HAT-1376 cells as measured by WST-1 tests. Mean±SD of three independent experiments. M: Marker.
Discussion
Our study demonstrates that EGF can be used for the effective targeting of EGFR on the surface of BC cells. The targeted toxin EGF-PE40 elicited a specific and concentration-dependent killing of EGFR-expressing BC cells based on the induction of apoptosis. Compared to the inhibitor erlotinib, which shows stoichiometric one to one binding with the tyrosine kinase moiety of EGFR, EGF-PE40 caused a 1,250- to 1,500-fold increased cytotoxicity in T24 and HT-1376 cells, respectively. This underlines the high potential of biologically active targeted toxins in the fight against BC.
Our study conforms to other studies in which targeted toxins against EGFR were produced and tested on BC cells. Sarosdy et al. developed the targeted toxin TP-40, consisting of TGF-α as the binding domain and PE40, which showed a high sensitivity in murine and human BC cells as well as primary tumors, including well-differentiated tumors, from BC patients (24). In a recent study, a targeted toxin was published consisting of the EFG ligand and Anthrax toxin from Bacillus anthracis. The immunotoxin was internalized within few minutes into BC cells of human, mouse or dog origin and killed them with LD50 values of <1 nM. Moreover, the immunotoxin induced an about 30% average tumor reduction after one intravesical application cycle in dogs with BC (25).
Future clinical application of EGF-PE40 in patients with BC must be considered not only in terms of specificity and efficacy, but also against the background of possible side effects. These include a possible immunogenicity of the targeted toxin leading to neutralization by anti-drug antibodies (ADA) and evocation of allergic reactions as well as on target/off tumor toxicities due to EGFR expression on various normal tissues. Immunogenicity of the EGF ligand is not expected, since EGF is of human origin. Immunogenicity of the PE40 domain can be reduced by de-immunization, i.e. elimination of immunodominant B- and T cell epitopes, or combination with immune modulating drugs [rev. in (26)]. Instillation of EGF-PE40 into the bladder could reduce systemic distribution and diminish or avoid on target/off tumor activities. Moreover, combination therapies with drugs that act synergistically with EGF-PE40 could reduce side effects. In studies with PE40 based immunotoxins targeting the prostate specific membrane antigen (PSMA) on prostate cancer cells, we showed synergisms with docetaxel or with the BH3 mimetic ABT-737, which led to an enhanced down-regulation of anti-apoptotic Bcl-2 proteins (23, 27, 28). Future experiments will show whether the effectivity of EGF-PE40 against BC cells can be increased in combination with chemotherapy or BH3 mimetics.
Taken together, the targeted toxin EGF-PE40 represents a promising candidate for the specific treatment of BC and merits additional research activities to be developed further towards the clinic.
Acknowledgements
This work was supported by a grant of the German Research Foundation (Grant No. WO 2178/2-1 to P.W.).
Footnotes
This article is freely accessible online.
Authors’ Contributions
P.W.: project administration; A.P.M., P.W.: study design; A.P.M., A.F., S.S.S., I.K., I.W.: performance of experiments, data collection; A.P.M., I.W., F.F.D., C.G., P.W.: writing, editing and reviewing of the manuscript. All Authors have approved the contents of the article.
Conflicts of Interest
The Authors declare no conflicts of interest regarding this study.
- Received June 15, 2021.
- Revision received June 25, 2021.
- Accepted June 28, 2021.
- Copyright © 2021 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.