

Abstract
Background/Aim: Growing evidence suggests that vitamin D3 exerts anticancer effects. The present study aimed to evaluate 25-hydroxyvitamin D3 (25(OH)D3) as a potential endocrine factor regulating proliferation and vitamin D receptor expression in LNCaP prostate cancer cells. Materials and Methods: Cell counting after treatment was utilized to assess the effect of 25(OH)D3 on cell proliferation. Changes in mRNA expression of the vitamin D receptors, VDR and PDIA3, were evaluated using droplet digital polymerase chain reaction (ddPCR). Results: 25(OH)D3 inhibited cell proliferation in a dose- and time-dependent manner. The inhibitory effect of 25(OH)D3 on cell proliferation was potentiated after inhibition of CYP17B1 and CYP24 by genistein, preventing further metabolism of 25(OH)D3 to 1,25-dihydroxyvitamin D3 (1,25(OH)2D3) and 24,25-dihydroxyvitamin D3 (24,25(OH)2D3). Expression of PDIA3 and VDR mRNA increased after treatment with 25(OH)D3, whereas the ratio between PDIA3 and VDR mRNA remained unchanged. Conclusion: 25(OH)D3 has a direct inhibitory effect on cell proliferation, which is enhanced and accelerated when the metabolism of 25(OH)D3 to 1,25(OH)2D3 and 24,25(OH)2D3 was inhibited by the CYP17B1 and CYP24 inhibitor genistein. Furthermore, treatment with 25(OH)D3 increased receptor transcript expression, suggesting an increased VDR stability and sensibility of the treated cells.
- Prostate cancer
- vitamin D
- 25-hydroxyvitamin D3
- genistein
- VDR
- PDIA3
- cell proliferation
Prostate cancer is a disease of increasing significance worldwide and is the most common malignant non-skin-cancer and the third most significant cause of death for men in the United States (1). It is estimated that half of a million people are diagnosed every year with prostate cancer worldwide. Prostate cancer is most common in Europe, the USA, Canada, and Australia and less common in Asia (2). Although death rates have decreased among African Americans compared to men of Caucasian origin, death rates among African American men remain more than twice as high. One evident factor is that African Americans have lower levels of circulating 25-hydroxyvitamin D3 (25(OH)D3) and, vitamin D insufficiency is far more common in African Americans than other Americans, primarily because pigmentation reduces the production of vitamin D in the skin (3, 4). Other risk factors for prostate cancer are increasing age or living at northern latitudes. It has been observed that men living at northern latitudes tend to have reduced vitamin D formation and lower levels of vitamin D3 in the circulation (5, 6).
Although prostate cancer etiology is still largely unknown, there is growing evidence that vitamin D3 exerts anticancer effects (7-17). Most of the vitamin D is obtained from exposure to sunlight. The solar ultraviolet β photons have a wavelength of 290-315 nm, allowing skin penetration. Once in the skin, UV-light causes photolysis of 7-dehydrocholesterol to precholecalciferol, which undergoes a rearrangement of its double bonds and forms cholecalciferol (vitamin D3). Vitamin D3 exits into the circulation where the hormone binds to vitamin D-binding protein. Vitamin D3 is transported and hydroxylated to create the active hormone, first in the liver and subsequently in the kidney. The first hydroxylation in the liver forms 25(OH)D3 and the second hydroxylation of 25(OH)D3 in the kidney by the enzyme 25(OH)D3-1α-hydroxylase (CYP27B1) or by 25(OH)D3-24-hydroxylase (CYP24) forms 1,25-dihydroxyvitamin D3 (1,25(OH)2D3) or 24,25-dihydroxyvitamin D3 (24,25(OH)2D3) (18-20). 1,25(OH)2D3 plays a significant role in regulating expression of genes associated with cell proliferation, differentiation, apoptosis, calcium homeostasis, immune responses, and blood pressure control. 1,25(OH)2D3 inhibits the growth of the human prostate cancer cell line LNCaP through the vitamin D receptor (nVDR) localized in the cytosol and the nucleus. 24,25(OH)2D3 has been shown to have anti-proliferative effects in certain types of cancer, such as colon cancer. 24,25(OH)2D3 causes milder hypercalcemia than 1,25(OH)2D3 and therefore appears as a more attractive therapeutic agent. Consequently, it has been suggested that 24-hydroxylase may function as an oncogene (15-17, 21-23).
There is evidence that demonstrates a correlation between low 25(OH)D3 levels and aggressive progression of prostate cancer and an earlier appearance of prostate cancer (2, 4, 14, 24). Ultraviolet radiation has been shown to have a protective effect against prostate cancer (25, 26). The mechanism is still not revealed, but it is believed that increased levels of vitamin D3 lead to increased levels of 25(OH)D3, which is considered to be transformed by the prostatic CYP27B1 enzyme to the active metabolite 1,25(OH)2D3. This metabolite interacts with the nVDR receptor and induces cell-cycle arrest and apoptosis (14, 18, 27). However, Hendrickson et al. (28) showed that high expression of the VDR in prostate tumors correlates with a reduced risk of lethal prostate cancer. In addition, Munetsuna et al. showed that the analog 25(OH)-19-nor-D(3) directly binds to VDR as a ligand and transports VDR into the nucleus to induce transcription of the cyp24A1 gene. Furthermore, Munetsuna et al. also showed that siRNA knockdown of the cyp24A1 gene did not affect the anti-proliferative effects caused by 25(OH)-19-nor-D(3), suggesting 25(OH)D3 as a potential candidate for anticancer therapy (29).
The present study aimed to evaluate 25(OH)D3 as a potential endocrine factor regulating proliferation and vitamin D receptor expression in LNCaP prostate cancer cells.
Materials and Methods
In vitro cell culture. LNCaP cells, derived from a metastasis at the left supraclavicular lymph node of a 50 years old PCa patient (Sigma Aldrich, St. Louis, MO, USA) were cultured in RPMI-1640 complete medium (“media”, Sigma-Aldrich) supplemented with 10% fetal bovine serum (FBS; Signa-Aldrich) and 1% PEST (Sigma-Aldrich) and kept at a low passage number (4-10). The cells were sub-cultured with Trypsin-EDTA solution and maintained at 37°C in 5% CO2.
Cell proliferation assay. Cells were seeded in 100 μl of media into 96 multiscreen plates at an initial seeding density of 5000 cells/well and, after 24 h, were starved for one day with media without FBS. After starvation, old media was removed and new media, containing FBS and 10–7, 10–8, 10–9, 10–10 or 10–11 M of 25(OH)D3 alone or in combination with 50 μM of 25(OH)D3-1α-hydroxylase and 25(OH)D3-24-hydroxylase inhibitor (genistein), were added to the wells. Wells with cultured cells with exclusively media and with media and ethanol (<0.01%) were used as positive control and as vehicle control, respectively. Multiscreen plates were incubated for 72, 96, and 120 h after treatment, and all the wells were loaded with 5 μM of Calcein (Calcein-AM Invitrogen, Waltham, MA, USA). Finally, fluorescence was measured at an excitation wavelength of 485 nm and an emission wavelength of 520 nm using the FLUOstar Omega (BMG Labtech, Ortenberg, Germany) for up to 2 h.
RNA extraction and reverse transcription (RT-PCR). Six well plates were seeded with 50.000 cells/well and starved for one day with media without FBS before treatment. After starvation, cells were incubated for 72 h with media containing FBS and 10–7, 10–8, 10–9, or 10–10 M of 25(OH)D3 alone or in combination with 50 μM genistein. Total RNA was extracted from each well using the RNeasy Mini Kit, including genomic DNA digestion with DNase (Qiagen, Hilden, Germany). Samples were eluted in RNase-Free water and stored at –80°C. RNA (up to 2 μg) was reversed transcribed to cDNA according to the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). The obtained cDNA samples were used as a reaction template in the ddPCR absolute quantification experiment.
ddPCR absolute quantification experiment. Droplet digital polymerase chain reaction (ddPCR; QX200, Bio-Rad, Hercules, CA, USA) was performed according to the manufacturer’s instructions. A total of 50 cDNA samples were analyzed to quantify the amount of PDIA3 and VDR target in single-plex reactions. The ddPCR assays, containing two primers and a TaqMan hydrolysis probe for each target, were designed using the Primer3 Plus web interface. The amount of cDNA in each reaction was 10,000 copies corresponding to 33 ng of DNA. Each sample was partitioned into 15,000-25,000 droplets that were transferred to a 96-well-plate to perform the PCR amplification.
After PCR amplification, the plate was read by the QX200 Droplet Reader with the settings for absolute quantification experiments. First, the amplitude limit was set manually to 1000 for PDIA3 and 4000 for VDR. Next, the Poisson-corrected determination of absolute template concentration (copies/μl) was calculated using QuantaSoft™ Analysis Pro Software (v1.0.596, Bio-Rad). Finally, the ratio of mRNA expression between PDIA3 and VDR (PDIA3/VDR) was calculated manually from template concentrations.
Statistical analysis. Cell proliferation results were normalized and expressed as % of control (mean±SEM). Differences in cell proliferation between treatment and control groups were assessed by One-Way ANOVA followed by Dunnett’s post-hoc test.
The mRNA expression of VDR and PDIA3 (template concentrations) is presented as median values with interquartile range. The ratio of PDIA3/VDR mRNA expression is presented as median with interquartile range and the maximum and minimum values. Differences in mRNA expression between VDR and PDIA3 and the ratio of mRNA expression PDIA3/VDR among groups were assessed with the Mann-Whitney and Kolmogorov-Smirnov tests, respectively. Non-parametric tests were used when mRNA expression of VDR and PDIA3 did not follow a normal (Gaussian) distribution according to Anderson-Darling and D’Agostino & Pearson tests.
All statistical tests were performed with GraphPad Prism version 8.3 for Windows (GraphPad Software, La Jolla, CA, USA). Levels of significance were set to *p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001.
Results
An inhibitory effect on the proliferation (approximately 15%) of LNCaP cells was observed for the two highest concentrations of 25(OH)D3 (10–8 M and 10–7 M) after 72 h incubation (p<0.05, Table I). Contrary, increased proliferation of LNCaP cells was observed at the three lowest concentrations of 25(OH)D3 (10-11, 10-10, and 10-9 M) after 96 h incubation (p<0.05, Table I). Incubation with 25(OH)D3 for 120 h showed a decrease in cell proliferation for all concentrations tested (p<0.01, Table I).
Proliferation of LNCaP cells incubated with 25(OH)D3 and the CYP27B1 and CYP24 inhibitor genistein (50 μM) for 72, 96, or 120 h.
After 72 h incubation, LNCaP cell proliferation was inhibited by 25(OH)D3 in combination with the CYP27B1 and CYP24 inhibitor genistein, except for incubation with 25(OH)D3 at concentrations of 10–9 M and 10–7 M (p<0.01, Table I). All studied concentrations of 25(OH)D3 together with genistein showed decreased cell proliferation after 96 h incubation even though not all reached statistical significance; the more potent inhibition of cell proliferation (65%-90%) was observed after 120 h (p<0.0001, Table I). Incubation with genistein exclusively demonstrated an inhibitory effect on LNCaP cell proliferation after 72 h but not at longer incubation times (96 and 120 h; Table I; Figure 1).

Proliferation of LNCaP cells incubated with 25(OH)D3 and genistein for A) 72 h, B) 96 h, and C) 120 h. LNCaP cells were stimulated with five concentrations of 25(OH)D3 (10–11, 10–10, 10–9, 10–8, or 10–7 M) alone or in combination with genistein (50 μM). Data are expressed as % of the control (mean±SEM). The stars indicate the degree of statistical significance between paired groups in the graph (*p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001). n.s.; Not significant.
The CYP17B1 and CYP24 inhibitor genistein combined with 25(OH)D3 demonstrated both dose- and time-dependent inhibition of LNCaP cell proliferation. In addition, the anti-proliferative effect was higher when genistein was administrated together with 25(OH)D3 compared to effects of 25(OH)D3 alone (Figure 1).
Expression of VDR and PDIA3 mRNA increased after 72 h of treatment with 25(OH)D3 (p<0.05, Table II). Further, treatment of cells with a combination of 25(OH)D3 and the inhibitor genistein increased expression of both VDR and PDIA3 mRNA (p<0.05, Table II). Finally, incubation with exclusively genistein increased the mRNA expression of VDR and PDIA3 in LNCaP cells (p<0.05, Table II).
mRNA expression of VDR and PDIA3 (copies/μl) in LNCaP cells stimulated with four concentrations of 25(OH)D3 (10–10, 10–9, 10–8 or 10–7 M) alone or in combination with genistein (50 μM), for 72 h.
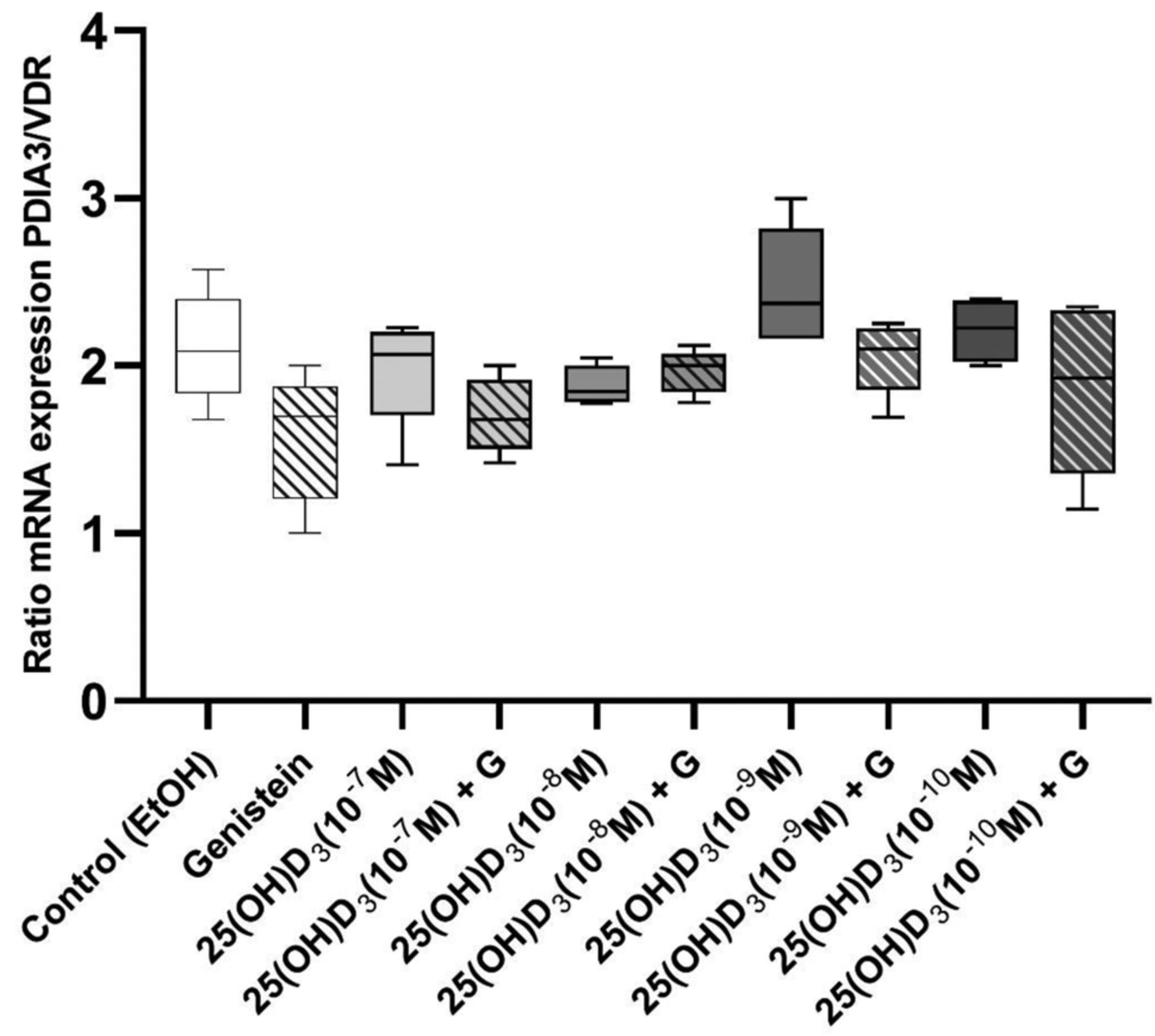
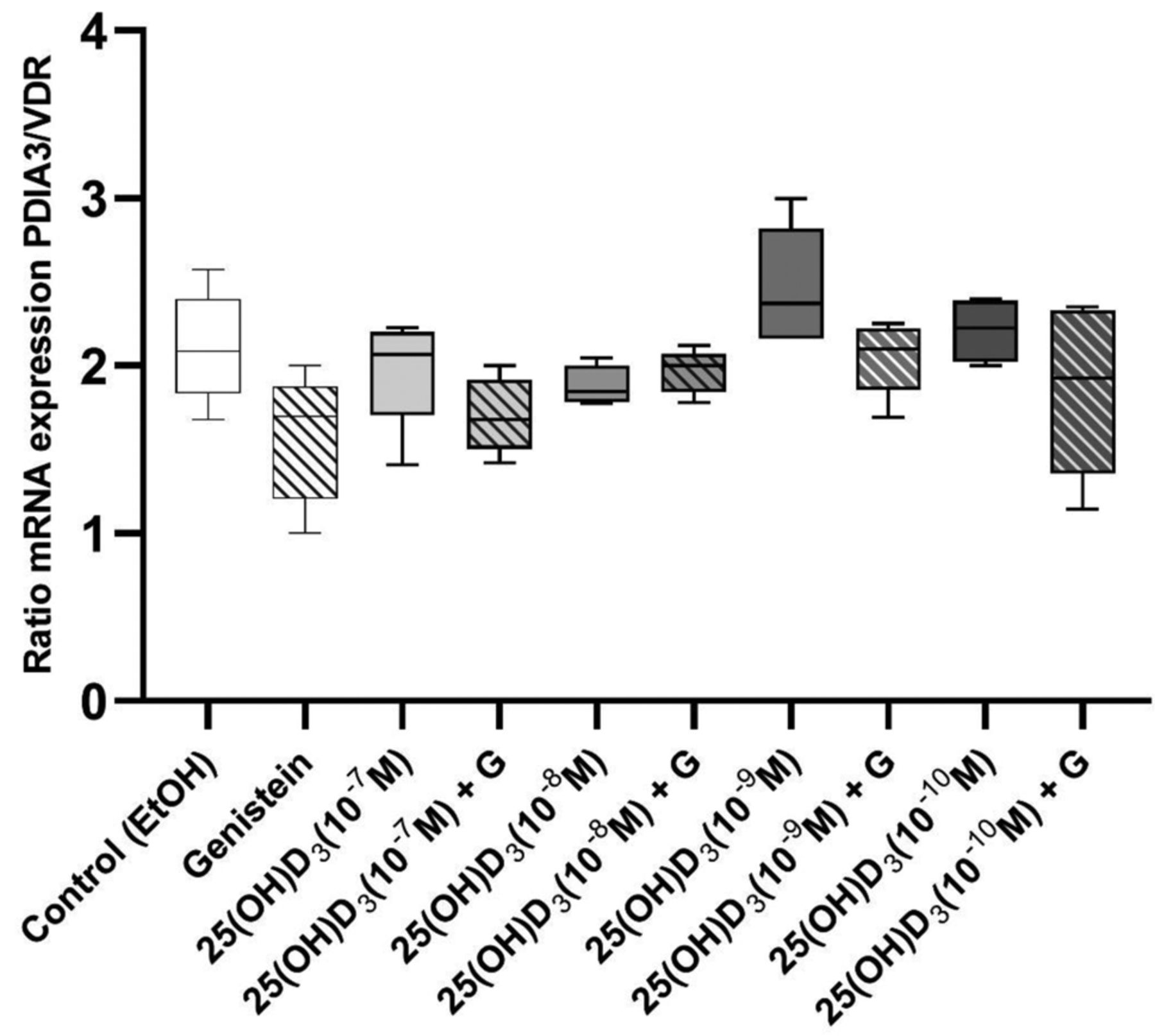
The ratio of PDIA3/VDR mRNA expression stayed stable and did not change after 72 h of treatment with 25(OH)D3 exclusively or in combination with genistein (Figure 2). In general, mRNA expression of PDIA3 was two-fold higher compared to expression of VDR in LNCaP cells (Figure 2).
{kind=link}
{kind=link}
Ratio of mRNA expression between PDIA3 and VDR (PDIA3/VDR) in LNCaP cells stimulated with 25(OH)D3 alone or in combination with genistein for 72 h. Data are presented as median with interquartile range and maximum and minimum values. LNCaP cells were stimulated with four concentrations of 25(OH)D3 (10–10, 10–9, 10–8, or 10–7 M) alone or in combination with genistein (50 μM). Differences in the ratio of mRNA expression PDIA3/VDR among groups were assessed with the Kolmogorov-Smirnov test. A non-parametric test was used when mRNA expression of VDR and PDIA3 did not follow a normal (Gaussian) distribution according to Anderson-Darling and D’Agostino & Pearson tests.
Discussion
The use of vitamin D analogs, which do not give rise to unwanted side effects, combined with other hormones or chemo-preventing agents might be beneficial in the future to treat prostate cancer and other forms of cancer However, the therapeutic use of 1,25(OH)2D3 is limited because elevations in serum 1,25(OH)2D3 can cause dangerous elevations in serum calcium levels. Our group has investigated the effects of the vitamin D pro-hormone, 25(OH)D3, on prostate cancer cell proliferation, which is known to have lower calcemic effects (30, 31). Our data strengthen the hypothesis that 25(OH)D3 has an inhibitory effect on the proliferation of LNCaP cells, enhances the stability of VDR and PDIA3, and exerts the inhibitory effects through a new and yet undescribed cellular signaling pathway. The data in the present study supports the hypothesis that 25(OH)D3 itself has an inhibiting effect on LNCaP proliferation, that initially takes place before the metabolic pathway is activated and the enzymes CYP27B1 and CYP24 convert 25(OH)D3 to 1,25(OH)2D3 and 24,25(OH)2D3. 1α,25(OH)2D3, is considered the active metabolite of vitamin D, and studies have shown an evident inhibitory effect on prostate cancer cell proliferation. The problem with using 1,25(OH)2D3 is that it causes hypercalcemia; thus, 25(OH)D3 may be an alternative treatment with fewer side effects (4, 18, 29, 30, 32).
Analyses that show a relationship between latitude or sunlight exposure and prostate cancer support the hypothesis above, as 25(OH)D3 tends to vary in relation to UV light exposure. 1,25(OH)2D3 concentrations in the circulation do not vary until 25(OH)D3 is distinctly deficient, suggesting that 1,25(OH)2D3 as well as 25(OH)D3 are essential factors regulating prostate cancer cell development. In support of the hypothesis that 25(OH)D3 affects prostate cancer, African American men, which have the highest incident of prostate cancer in the world, have lower circulating levels of 25(OH)D3 and higher levels of 1,25(OH)2D3, compared to men with European heritage (3, 6).
When LNCaP prostate cancer cells were treated with the 25(OH)D3-1α-hydroxylase and 25(OH)D3-24-hydroxylase inhibitor genistein in this study, cell proliferation was inhibited at all concentrations, suggesting that genistein potentiates the inhibitory effects of 25(OH)D3 on cell proliferation. Genistein is used to block the activity of 25-hydroxyvitamin-D3-1α-hydroxylase and 25(OH)D3-24-hydroxylase, which is necessary to determine if the inhibitory effect is due to 25(OH)D3 or its metabolites 1,25(OH)2D3 and 24,25(OH)2D3. After 72 h, treatment with genistein alone showed increased inhibition of cell proliferation compared to the control group. This result suggests that genistein initially has an inhibitory effect that diminishes at the two most extended incubation times of 96- and 120-h. This hypothesis is supported by a study by Rao et al., 2004. In this report, the inhibitory effect of genistein is tested both separately and with 1,25(OH)2D3 in LNCaP cells. In both cases, inhibitory effects were observed, and when the two of them were combined, this effect was strongly enhanced. Furthermore, Rao and his group observed that genistein and 1,25(OH)2D3 mediated their effect through the nVDR receptor and p21, which is essential for cell cycle arrest in G0-G1. Genistein was able to enhance the stability of the nVDR receptor and increase the levels of p21. They were also able to demonstrate the requirement of p21 in genistein-mediated growth inhibition. Furthermore, genistein has been shown to be a potential inhibitor in prostate cancer cells other than LNCaP. Edward Kyle et al., confirmed genistein’s ability to inhibit prostate cancer cell growth by demonstrating that genistein induced cell apoptosis and that it had a unique effect on cellular protein-tyrosine kinase activity (33, 34). The present study confirms the previous results from Rao et al., where treatment with 25(OH)D3 increased receptor transcript levels, suggesting increased VDR stability and sensibility of the treated cells.
One might ask whether the diminishing effect of genistein on cell proliferation after 96- and 120-h treatment may be because the LNCaP cells have metabolized genistein in such a way that the inhibitor no longer influences them. Genistein could give a better inhibitory effect on LNCaP cell proliferation at the first hours of treatment. On the contrary, 25(OH)D3 has an enhanced inhibitory effect at the longest time periods of 96 and 120 h. LNCaP cells treated for 96 and 120 h with 25(OH)D3 and genistein, showed inhibition of cell proliferation at all 25(OH)D3 concentrations administered. The decreased cell proliferation may be due to the blockage of 25(OH)D3-1α-hydroxylase and 25(OH)D3-24-hydroxylase, which leads to a significantly strengthened effect on inhibiting cell proliferation. We suggest, based on our findings, that 25(OH)D3 has a direct inhibitory effect on cell proliferation, which is enhanced and accelerated when the CYP17B1 and CYP24 inhibitor genistein inhibits the metabolism of 25(OH)D3 to 1,25(OH)2D3 and 24,25(OH)2D3. These results do not agree with those of Ju-Yu Hsu et al. (32). They demonstrated inhibitory effects of 25(OH)D3 on the growth of normal prostate cancer cells, some inhibition in primary prostate cancer cells, but no inhibition in LNCaP cells. They reported that due to reduced levels of 1α-hydroxylase and its activity in this particular cell line, the LNCaP cells were resistant to treatment with 25(OH)D3 and mean that 1α-hydroxylase is an essential component of growth inhibition by 25(OH)D3. They indicated a strong correlation between the growth-inhibitory activity and 1α-hydroxylase activity; however, they did not clarify how long the cell line was treated with 25(OH)D3. Therefore, it can be assumed that the lack of inhibitory effect on cell growth may be due to the short time-period of treatment (32). However, more recent studies by Susa et al. (30) and Munetsuma et al. (31) have shown that 25(OH)D3 has the same target gene expression profiles as 1,25(OH)2D3 in LNCaP prostate cancer cells, suggesting regulation of prostate cancer cell biology via interaction with VDR, as well as an anti-proliferative action in PZ-HPV-7 prostate cells, supporting the present study.
The present study suggests that inhibition of CYP27B1 and CYP24 possibly increase the clinical outcome of individuals diagnosed with prostate cancer. Meta analyses on risk factors linked to prostate cancer incidence have shown that circulating levels of 25(OH)D3 have U-shaped correlations, where low and high levels stimulate whereas intermediate levels inhibit prostate cancer progression (35). It is generally accepted that the higher the dose of 1,25(OH)2D3 used, the greater the anticancer effects will be achieved (8, 36, 37). When used therapeutically at higher than physiologic doses, the major side effect of 1,25(OH)2D3 is the risk of inducing hypercalcemia and renal stones (38). There is also a theoretical risk of causing vascular calcification and cardiovascular disease, as shown by Mizobuchi et al. when studying the effects of vitamin D in chronic kidney disease (39). The present study put forward the hypothesis that 25(OH)D3 may constitute a putative therapeutic agent. From a clinical point of view, 25(OH)D3 is highly interesting since 25(OH)D3 does not cause hypercalcemia in contrast to its metabolite 1,25(OH)2D3. Therefore, 25(OH)D3 may be used even as a prodrug for prostate cancer treatment. Because of its less calcemic activity, the dosage could be increased compared to the active metabolite 1,25(OH)2D3 (32). To further evaluate the impact of different vitamin D3 metabolites on prostate cancer, future studies should focus on limiting further metabolism of 25(OH)D3 to 1,25(OH)2D3 as well as maintaining circulating 25(OH)D3 levels within close limits.
Footnotes
This article is freely accessible online.
Authors’ Contributions
D Larsson: Principal Investigator, Conceptualization, Investigation, Methodology, Analysis, Visualisation, Writing-Reviewing and Editing; MA Díaz Cruz: Investigation, Methodology, Analysis, Visualisation; Writing; S Karlsson: Investigation, Methodology, Analysis, Visualisation, Writing; M Faresjö: Investigation, Methodology, Analysis, Visualisation, Writing; A Papadopoulou Khamou: Investigation, Methodology, Analysis.
Conflicts of Interest
The Authors declare no conflicts of interest in relation to this study.
- Received August 18, 2021.
- Revision received September 1, 2021.
- Accepted September 2, 2021.
- Copyright © 2021 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.