

Abstract
Background/Aim: Glioblastoma (GBM) is one of the most lethal solid cancers due to its highly invasive nature. The malignant potential of GBM cells might be partially regulated by surrounding normal cells, such as oligodendrocytes or fibroblasts. The aim of this study was to examine the interaction between stromal cells and GBM cells. Materials and Methods: Two GBM cell lines were used. The effect of stromal cells, oligodendrocytes or fibroblasts, on the invasive ability of GBM cells was examined by wound-healing assay and invasion assay. Results: Oligodendrocytes, in contrast to fibroblasts, significantly increased the migration and invasive ability of GBM cells. Angiopoietin-2 levels were high in the conditioned medium obtained from oligodendrocytes. Angiopoietin-2 significantly increased the motility of GBM, and the motility-stimulating activity of the oligodendrocytes-derived conditioned medium was significantly decreased by anti-angiopoietin-2-neutralizing antibody. Conclusion: Glioma stromal cells, oligodendrocytes, might up-regulate the invasiveness of GBM cells via angiopoietin-2 signaling.
- Glioblastoma
- oligodendrocyte
- angiopoietin-2
- fibroblasts
- invasion
Diffuse glioma comprises the majority of primary brain tumors (1, 2). Recently, diffuse gliomas were classified as WHO grade II, III, and IV, based not only on their growth pattern and behavior, but also on genetic driver mutations in IDH1 and IDH2 genes (3). The current classification is based on the theory that primary glioblastoma (GBM) occurs de novo without the need of a less malignant precursor tumor, whereas secondary GBM arises from a lower-grade diffuse glioma (WHO grade II or III) (4). In spite of the standard multimodal treatments including surgical resection, chemotherapy, and radiotherapy, the median survival time of GBM patients is only around 14.6 months (1, 2). Surgical resection of diffuse gliomas is frequently non-curative because of their highly invasive nature (5). Therefore, a more thorough investigation of the invasive ability of GBM cells is of utmost importance for the discovery of novel therapies to improve the prognosis of patients with diffuse glioma.
Tumor development is not only determined by malignant tumor cells themselves, but also by the tumor stroma. Recently, it was reported that tumor progression depends on the interactions between tumor cells and its surrounding tumor stromal cells (6, 7). Oligodendrocytes, fibroblasts, and astrocytes are considered stroma cells for diffuse glioma, especially GBM (8, 9). However, the correlation between diffuse glioma cells and its surrounding tumor stroma cells has not been fully clarified. This study aimed to investigate the effect of stromal cells on the invasion of GBM cells and identify factors/signaling pathways that regulate migration and invasiveness of GBM. To the best of our knowledge, this is the first study to show that oligodendrocyte cells might up-regulate the invasive ability of GBM cells via angiopoietin-2 signaling.
Materials and Methods
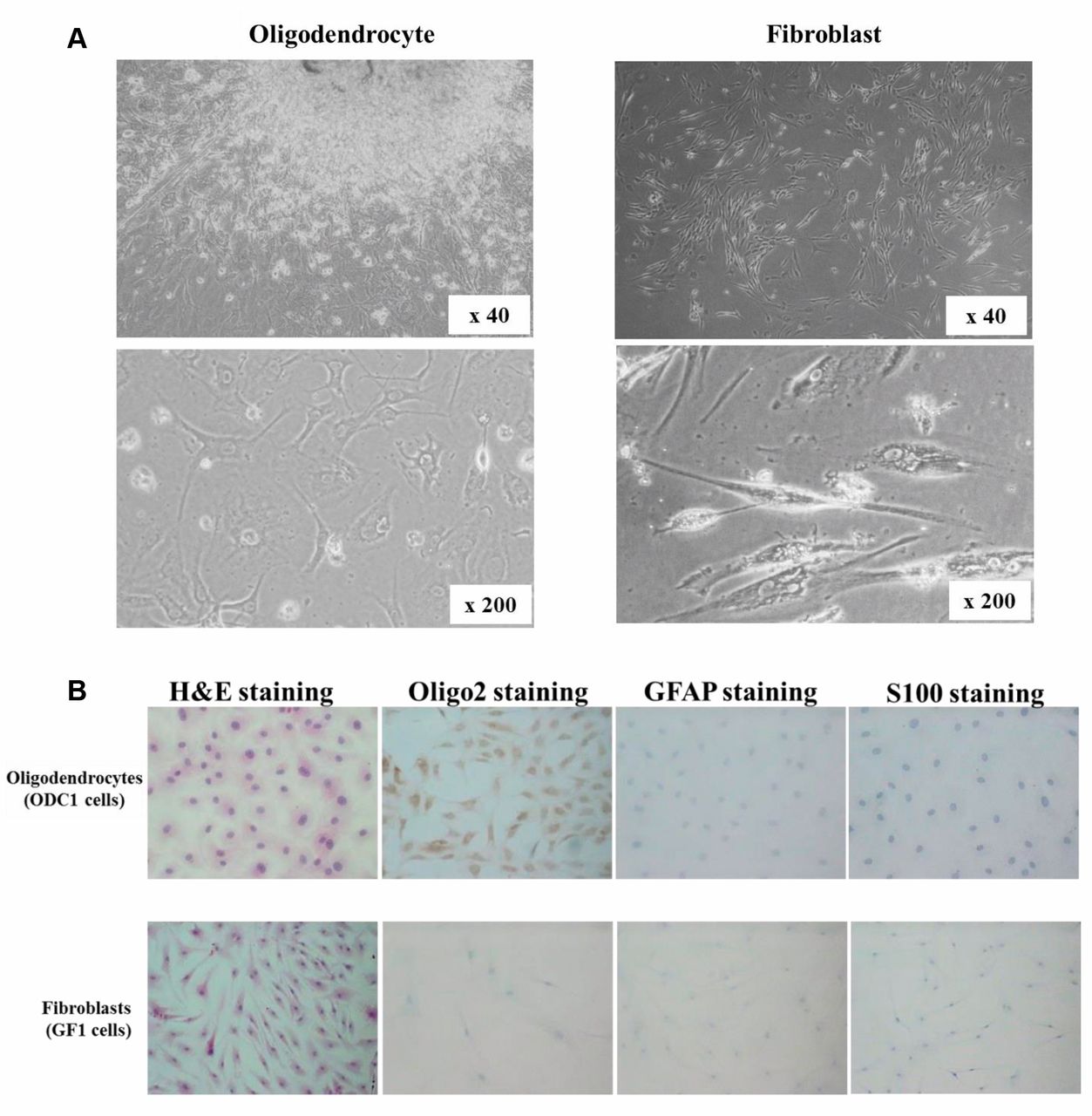
Cell culture and cell lines. Two glioblastoma (GBM) cell lines, T98G and U251 were purchased from the JCRB Cell Bank (Osaka, Japan). Two types of tumor stromal cells, oligodendrocytes and fibroblasts, were established from human glioma tissue in Osaka City University. Two oligodendrocyte cell lines, ODC1 and ODC2, were derived from tumoral tissues of patients with low-grade glioma (WHO Grade II), and one fibroblasts cell line, GF1, was derived from tumoral tissues of patients with GBM (Figure 1A). The presence of oligodendrocytes and fibroblasts was confirmed by immuno-histochemical staining. Oligodendrocyte cells, ODC1 and ODC2, were positive for the oligodendrocyte marker Olig2, but not GFAP or S100 which are markers for astrocytes. Fibroblasts, GF1 cells, were negatively-stained for Olig2, GFAP, and S100 (Figure 1B). The primary culture was prepared as previously reported (10). Briefly, each glioma specimen was excised under aseptic conditions, and minced with the help of forceps and scissors. The culture medium consisted of c Dulbecco's modified Eagle medium (DMEM; Wako, Osaka, Japan) supplemented with 10% fetal bovine serum (FBS; Nichirei Biosciences Inc., Tokyo, Japan), 100 IU/ml penicillin (Wako), 100 mg/ml streptomycin (Wako), and 0.5 mM sodium pyruvate (Wako). Cells were cultured in an atmosphere of 5% CO2 at 37°C. Oligodendrocytes and fibroblasts were used from the 3rd passage to the 10th passage in culture, mainly at the 5th passage.
Preparation of conditioned medium (CM). Two semi-confluent oligodendrocyte cultures, ODC1 and ODC2, and fibroblasts, GF1, grown in 100-mm plastic dishes containing DMEM with 10% FBS, were washed with phosphate-buffered saline (PBS; Wako) and incubated for additional 3 days in 5 ml of serum-free DMEM. The supernatant was collected and stored as conditioned media (CM) at −20°C until use. All experiments were performed in medium containing 1.5% FBS. As a control, DMEM was used instead of CM.
Compounds. Angiopoietin-2 was purchased for Novus Biologicals, Littleton, CO, USA), and anti-angiopoietin-2-neutralizing antibody was purchased from Cloud-Clone (TX, USA).
Wound-healing assay. GBM cells were cultured in 96-well plates (Essen ImageLock, Essen Instruments, Birmingham, UK). After the cells reached semi-confluence, a wound was created in the cell monolayer with the 96-well WoundMaker (Essen Bioscience, MI, USA). GBM cells were cultured in DMEM with 2% FBS for 48 h with or without CM or angiopoietin-2 at various concentrations. Scratched fields were imaged every 4 h and were monitored with Incucyte Live-Cell Imaging System and software (Essen Instruments).
Invasion assay. The in vitro invasiveness of GBM cells, T98G and U251, was measured by a two-chamber Matrigel invasion assay. The upper chambers (Falcon) which had an 8-μm pore membrane filter coated with 50 μg of Matrigel were placed in a 24-well culture plate (lower chamber). T98G or U251 cells (2.5×104 cells/500 μl/chamber) were seeded in the upper chamber, and 500 μl medium containing 250 μl CM from stromal cells and 250 μl glioblastoma culture medium with or without anti-angiopoietin-2-neutralizing antibody (500 ng/ml), or angiopoietin-2 (10 ng/ml) was added to the low chamber. After incubation for 24 h, T98G or U251 cells that invaded the matrigel and the membrane were stained by Diff-Quick (Sysmex, Kobe, Japan) and were manually counted under a microscope at ×200 magnification. Six randomly chosen fields were counted for each condition, and the mean of the six fields was calculated as the sample value.
Cell morphology. T98G and U251 cells were seeded and incubated for 7 days in the presence of 50% CM from stromal cells with 2% FBS. The morphology of the T98G and U251 cells was then observed under a light microscope.
Proteome profiler protein array. A total of 102 cytokines were measured using Proteome Profiler Human XL Cytokine Array Kit (R&D Systems), as previously reported (11). CM from oligodendrocytes or fibroblasts was mixed with a cocktail of biotinylated detection antibodies. After incubation with streptavidin-horseradish peroxidase, chemiluminescent detection reagents were added and the chemiluminescence was detected by LAS-4000 mini (GE Healthcare) and analyzed by Image Quant TL 7.0 (GE Healthcare).
Enzyme-linked immunosorbent assay (ELISA). The levels of angiopoietin-2 in the CM were quantified using an enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems) as per the manufacturer's protocol. Briefly, 200 μl of sample were added to each well coated with monoclonal antibody, and incubated for 1 h. After 4 washes, absorbance was measured on a microplate reader at 450 nm.
Proliferation assay. The proliferation of GBM cells, T98G and U251, was determined by Cell Counting Kit-8 (CCK-8; Dojindo) assay or MTT assay. Application of CCK-8 assay was as follows. A total of 1×103 GBM cells were seeded into two 96-well plates with culture medium exposed to 50% CM or anti-angiopoietin-2-neutralizing antibody at the concentration of 100 ng/ml (diluted with PBS).
In order to perform the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, cells were seeded at 1×103 per well in 96-well plates with 50% CM, angiopoietin-2, or anti- angiopoietin-2-neutralizing antibody, and incubated for 72 h. Subsequently, 100 μl of culture medium and 20 μl of MTT solution at the concentration of 2 mg/ml (diluted with PBS; Promega, Tokyo) were added to each well, and the absorbance at 570 nm was analyzed using a microplate reader (Bio-Rad 550; Bio-Rad, Tokyo).
Statistical analysis. Data are expressed as mean±SD, and differences in data were analyzed using the unpaired Student's t-test. p-Values <0.05 were considered significant.
Results
Effect of CM from stromal cells on the migration of glioblastoma cells. Figure 2A shows a representative phase-contrast image of the wound-healing assay. The migratory ability of GBM cells was increased by the CM from oligodendrocytes, but not by the CM from fibroblasts. The number of migrating T98G cells was significantly increased after the addition of CM from ODC1 and ODC2 cells for 24 h, as compared with the control. The number of migrating U251 cells was also significantly increased after the addition of CM from ODC1 and ODC2 cells for 12 h, as compared with the control. In contrast, the number of migrating T98G cells and U251 cells was not increased by the CM from GF1 fibroblasts (Figure 2B).
Effect of CM from stromal cells on the invasive ability of glioblastoma cells. Figure 3A shows a representative image of the invaded cancer cells. The CM from oligodendrocytes, ODC1 and ODC2, significantly increased the number of invading T98G and U251 cells. In contrast, CM from fibroblasts, GF1, did not affect the invasive ability of glioma cells (Figure 3B). On the other hand, the CM from oligodendrocytes or fibroblasts did not affect the proliferation of T98G or U251 cells. T98G and U251 cells did not show any morphologic changes following the addition of these CM from oligodendrocytes and fibroblasts (data not shown).

Primary culture and immunostaining of glioma stromal cells. (A) Representative picture of stromal cells. Oligodendrocytes, ODC1 and ODC2, were derived from tumor specimen of grade II glioma. Fibroblasts, GF1, were derived from tumor specimen of glioblastoma. (B) Immunohistochemical staining for Olig2 and GFAP. ODC1 were stained with Olig2, but not GFAP antibodies. GF1 were stained with neither Olig2 nor GFAP antibodies.
Cytokine production from stromal cells in glioma. A cytokine array kit, which detects the relative levels of 102 different cytokines, was used. As shown in Figure 4, the protein array indicated that many chemokines and cytokines were present in the CM from the glioma stromal cells, oligodendrocytes and fibroblasts. Among them, only the angiopoietin-2 was found at a higher level in the CM from oligodendrocytes, ODC1 and ODC2, compared with that from fibroblasts, GF1 (Figure 4A, arrow). In contrast, higher levels of chitinase 3-like 1, DPPIV, CXCL5, GM-CSF, IGFBP-3, IL-6, IL-8, IP-10, kallikrein 3, thrombospondin-1, and VEGF, were produced from fibroblasts, compared to oligodendrocytes (Figure 4A, arrowheads). The levels of angiopoietin-2 in the CM from oligodendrocytes and fibroblasts were examined by ELISA. Oligodendrocytes, ODC1 and ODC2, produced higher than 5 ng/ml of angiopoietin-2. In contrast, fibroblasts, GF1, produced lower angiopoietin-2 levels (Figure 4B).

The effect of conditioned medium (CM) from stromal cells on the migratory activity of glioblastoma cells (GBM cells). (A) Representative picture of the wound-healing assay. Migration of GBM cells was increased at 24 h and 48 h following the addition of CM from oligodendrocytes, but not CM from fibroblasts. (B) The time course of migratory activity of glioblastoma cells following the addition of CM from stromal cells. The migration of T98G cells was significantly increased 24 h after the treatment with CM from the oligodendrocyte cell lines, ODC1 and ODC2. The migration of U251 cells was significantly increased 12 h after the addition of CM from ODC1 and ODC2. In contrast, CM from fibroblast did not affect the migration of GBM cells. The mean of eight fields was calculated as the sample value. *p<0.05, **p<0.01.

The effect of CM from oligodendrocytesls on the invasive ability of glioblastoma cells. (A) Representative image of the invasion assay. Migration of GBM cells was increased by the addition of CM from oligodendrocytes, but not CM from fibroblasts. (B) Effect of CM from stromal cells on the invasive ability of glioblastoma cells. The invasive ability of GBM cells was significantly increased with CM from oligodendrocytes, ODC1, but not CM from fibroblasts, GF1.
Effect of angiopoietin-2 and anti-angiopoietin-2-neutralizing antibody on the invasive ability of glioma cells. To identify the motility-stimulating factor in the CM from oligodendrocytes the wound-healing assay was used. Angiopoietin-2 significantly increased the motility of T98G cells and U251 cells (Figure 5A). Furthermore, the role of this cytokine in the migration-stimulating activity of CM from oligodendrocyte cells was also examined, using an anti-angiopoietin-2-neutralizing antibody. The migration-stimulating activity was significantly (p<0.05) decreased by the anti-angiopoietin-2-neutralizing antibody (Figure 5B).

Cytokine analysis of the CM from stromal cells. (A) Profiles of cytokines in the CM from cultures of stromal cells. Antibodies to 102 cytokines were spotted on nitrocellulose membranes. Angiopoietin-2 was found at higher levels in the CM from oligodendrocytes, ODC1 and ODC2, compared to that from fibroblasts, GF1 (arrow). In contrast, chitinase 3-like 1, DPPIV, CXCL5, GM-CSF, IGFBP-3, IL-6, IL-8, IP-10, kallikrein 3, thrombospondin-1, and VEGF, were produced at higher levels from fibroblasts, compared to oligodendrocytes. (B) Production of angiopoietin-2 by oligodendrocytes and fibroblasts was assayed by ELISA. Oligodendrocytes, ODC1 and ODC2, produced more than 5 ng/ml of angiopoietin-2. Fibroblasts, GF1, produced lower levels of angiopoietin-2.
Discussion
In this study, the role of two types of stromal cells from glioma specimens, oligodendrocyte cells and fibroblasts, was assessed. The stromal cell types were identified by immunohistochemical staining. Oligodendrocyte cells, ODC1 and ODC2, were positive for Olig2, but not GFAP (12). Fibroblasts, GF1 cells, were stained with neither Olig2 nor GFAP. Interestingly, the two oligodendrocyte cell lines were derived from low-grade glioma tissues (WHO Grade II). The fibroblasts cell line, GF1, was derived from GBM tissues (WHO Grade IV). These findings suggest that oligodendrocytes might be abundant in low-grade glioma tissues and fibroblasts might be abundant in GBM tissues.
Using these glioma stromal cells, the effect of stromal cells on the motility and proliferation of GBM cells, T98G and U251 was examined. Our results demonstrated that CM from the oligodendrocyte cells, ODC1 and ODC2, significantly stimulated the migratory activity of GBM cells but not their proliferation. In contrast, the CM from fibroblast cells did not stimulate the motility or the proliferation of GBM cells. These findings suggest that oligodendrocytes might produce the motility-stimulating factor for GBM cells, but not for fibroblast cells. Thus, oligodendrocytes acting as tumor stromal cells might contribute to the development of the malignant phenotype of glioma cells. Secondary GBM develops through progression from lower-grade diffuse glioma (WHO grade II or III), whereas primary GBM occurs de novo without evidence of less malignant precursor lesions (4). The acquirement of the malignant phenotype of gliomas might be associated with the interaction between oligodendrocyte cells and low-grade glioma cells such as WHO Grade II.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of angiopoietin-2 and anti-angiopoietin-2-neutralizing antibody on the invasive ability of glioma cells. (A) Angiopietin-2 (500 nM) significantly increased the invasive ability of GBM cells, T98G cells (p<0.05) and U251 cells (p<0.01). (B) CM from oligodendrocyte, ODC1 and ODC2, significantly increased the migration of GBM cells, T98G and U251. The migration-stimulating activity of the CM from oligodendrocytes, ODC1 and ODC2, was significantly decreased after treatment with an anti-angiopoietin-2-neutralizing antibody (T98G cells, p<0.05 and U251 cells, p<0.05).
We next aimed to clarify a paracrine factor(s) produced by the oligodendrocyte cells that increases the invasive ability of GBM cells. Based on the hypothesis that the differences in cytokine production between oligodendrocyte cells and fibroblasts might be associated with the motility-stimulating activity of GBM cells, the levels of 102 cytokines were examined. The oligodendrocyte cells produced higher levels of angiopoietin-2 compared to fibroblasts. In fact, angiopoietin-2 increased the motility of GBM cells, T98G and U251. Our present findings suggest that angiopoietin-2 from oligodendrocytes might be a promising therapeutic target for GBM. In support, it has been reported that dual blockade of angiopoietin-2 prolongs survival of patients with glioblastoma (13, 14).
In contrast, among the 102 cytokines, 11 cytokines, such as chitinase 3-like 1, DPPIV, CXCL5, GM-CSF, IGFBP-3, IL-6, IL-8, IP-10, kallikrein 3, thrombospondin-1, and VEGF, were produced at higher levels by fibroblasts than by oligodendrocytes. Since the anti-angiopoietin-2-neutralizing antibody partially inhibited the motility-stimulatory activity of CM from oligodendrocyte cells, the cytokines produced by fibroblasts might be associated with inhibition of the invasive activity of GBM cells.
In conclusion, glioma stromal cells, oligodendrocytes, might up-regulate the invasiveness of GBM cells via angiopoietin-2 signaling pathway. Thus, angiopoietin-2 might be a promising target for the treatment of GBM.
Acknowledgements
The Authors would like to thank Megumi Yoshino (Department of Neurosurgery, Osaka City University Graduate School of Medicine), and Maiko Nagai (Osaka City University Graduate School of Medicine), for providing technical assistance. This study is partially funded by JSPS KAKENHI (Grant-in-Aid for Scientific Research B; Grant Numbers JP23390329).
Footnotes
Authors' Contributions
Study concept and design: T. Kawashima, M. Yashiro; Acquisition of data: T. Kawashima, H. Kasashima, Y. Terakawa, T. Uda, K Nakajo, R. Umaba, Y. Tanoue, S. Tamrakar; Analysis and interpretation of data: T. Kawashima, M. Yashiro, H. Kasashima, S. Tamrakar; Drafting of the manuscript: T. Kawashima, M. Yashiro, S. Tamrakar; Study supervision: M. Yashiro, K. Ohata.
This article is freely accessible online.
Conflicts of Interest
There are no financial or other interests present with regard to the submitted manuscript that might be construed as a conflict of interest.
- Received December 19, 2018.
- Revision received January 9, 2019.
- Accepted January 10, 2019.
- Copyright© 2019, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved