

Abstract
Background/Aim: The perioperative phase is supposed to be a period with high vulnerability for cancer dissemination. Acetaminophen and metamizole are common analgesics administered during this phase. We investigated the effect of acetaminophen, metamizole and 4-methylaminoantipyrine (MAA) on proliferation and apoptosis of colon carcinoma cell lines (SW 480 and HT 29). Materials and Methods: Proliferation was detected by cell proliferation ELISA BrdU, and apoptosis by Annexin V staining. Cytochrome c and caspase 3, 8 and 9 expression levels were detected by western blot. Results: Acetaminophen, metamizole or MAA caused slight changes in proliferation. Acetaminophen, metamizole or the combination increased apoptosis in both cell lines. All agents decreased caspase 3 and 8 expression in SW480. Acetaminophen decreased caspase 9 expression in both cell lines. Conclusion: In clinically relevant doses, acetaminophen and/or metamizole induce apoptosis in both colon cancer cell lines. Both mitochondrial and death receptor pathways might be involved in acetaminophen-induced apoptosis.
- Cancer
- acetaminophen
- metamizole
- cell proliferation
- apoptosis
For a variety of solid tumors, surgery is the first-line treatment. However, evidence is growing that the perioperative phase is a period with high vulnerability for cancer dissemination (1). The fatal combination of perioperative immunosuppression, release of cancer cells by surgical trauma and enhanced levels of growth factors increase the risk for cancer recurrence during the perioperative period. Traumatization of tumor vessels and surgical manipulation enable cancer cells to enter the circulation (2). The excessive release of growth factors and inbalance between pro- and antiangiogenetic factors during the wound healing process can foster the growth of disseminated cancer cells and micrometastases (2). Also impairment of anticancer surveillance can be detected during the first postoperative days (3, 4). Anesthetic and analgesic agents are applied during this high-risk phase for cancer progression. For this reason, it is under discussion, if the selection of a special anesthesia technique or pain therapy regime can influence the oncological outcome of cancer patients (3-5).
The combination of analgesic agents with different mechanisms of action is a recommended concept in perioperative pain management. Acetaminophen, metamizole and NSAIDs are commonly used non-opioid analgesics (6). Results of several studies indicate that NSAIDs reveal anticancer potency by inhibiting cell proliferation, inducing apoptosis and disturbing tumor stroma interaction processes such as angiogenesis and cell migration (7-9). However, if acetaminophen and metamizole exert similar antineoplastic efficiency as NSAIDs has not been well elucidated yet.
The aim of this study was to investigate the effects of acetaminophen, metamizole and its active metabolite 4-methylaminoantipyrine (MAA), on cell proliferation and apoptosis rate of SW 480 and HT29 colon carcinoma cell lines in vitro.
Materials and Methods
Reagents. Commercially available acetaminophen (Merck, Darmstadt, Germany), metamizole (Fluka, Buchs, Switzerland), MAA and staurosporine (both Sigma-Aldrich, Steinheim, Germany) were used for this study. A stock solution was prepared by dissolving the agents in standard growth medium, final concentrations were achieved by diluting the stock solution with standard growth medium. All solutions were prepared freshly prior to use.
Cell lines. HT 29 and SW 480 colon carcinoma cell lines were purchased from the German Collection of Microorganism and Cell Culture (DSMZ). Standard growth media (RPMI 1640, Pan Biotech, Aidenbach, Germany) for SW 480 and DMEM (Sigma-Aldrich, Steinheim, Germany) for HT 29 contained 10% Fetal calf serum (FCS) (Sigma Aldrich, Steinheim, Germany) and 2 mM L-Glutamine (Sigma Aldrich, Steinheim, Germany) supplemented with 5% penicillin plus streptomycin (Sigma Aldrich, Steinheim, Germany). SW 480 culture media additionally contained 2 mM sodium pyruvate (Appli Chem, Darmstadt, Germany). Cells were maintained in monolayer culture and were cultured in a humidified CO2 atmosphere (5%) at 37°C. Experiments were performed when cells reached ~80% confluence.
Cell proliferation. The cell proliferation ELISA BrdU (Roche applied science, Mannheim, Germany) was used for the detection of cell proliferation rate. In brief, cells (SW 480 5×103, HT 29 (3×103) were seeded in 96-well plates (Costar, Bodenheim, Germany) and incubated overnight to allow attachment. Cells were incubated for 48h with acetaminophen (0-1000 μM), metamizole (0-500 μM) or MAA (0-500 μM) and for 16 h with BrdU labeling solution. After cell fixation and DNA denaturation, cells were labeled with Anti-BrdU-POD solution for 90 min. Cells were washed, incubated with TMB substrate for 15 min and immune complexes were measured by detecting absorbance at 405 and 490 nm. All tests were performed in duplicates, eight wells per treatment group were used and tests were repeated at least twice.
Apoptosis analysis. SW 480 (8×105) or HT 29 (3,5×105) cells were seeded in T 25 cm2 cell culture flasks (BD Falcon, Heidelberg, Germany). After 24h attachment time, cells were incubated with the test agents (250 μM or 1000 μM) for 3-48 h. One μM staurosporine (Sigma Aldrich, Steinheim, Germany) was used for positive control. Floating cells were preserved by decanting supernatant. Adherent cells were rinsed with PBS (37°C) (Sigma Aldrich Steinheim, Germany) and detached by standard trypsinization. FITC Annexin V Apoptosis detection kit (BD Bioscience, Franklin Lakes, NJ, USA) was used according to the manufacturer's protocol. In brief, floating and harvested cells were mixed, washed twice with cold PBS and resuspended in binding buffer at a final concentration of 106 cells/ml. 5 μl of FITC Annexin and 5 μl of PI were added to 100 μl of the cell suspension containing 105 cells. After mixing gently, cell suspension was incubated at room temperature in the dark for 15 min. After adding 400 μl of binding buffer, cells were analyzed by flow cytometry using FACS Calibur (BD Bioscience, Franklin Lakes, NJ, USA) and Cellquest™ Pro software (BD Bioscience, Franklin Lakes, NJ, USA). All tests were performed in duplicates and repeated at least twice. For data analysis Fowjo v10.0.7 (Ashland, OR, USA) was applied.
Western blot analysis. Antibodies: For immunoblotting, membranes were probed with antibodies against caspase 3, caspase 8, caspase 9, cytochrome c (Cell signaling, Leiden, the Netherlands), and ß-actin (Sigma-Aldrich, Steinheim, Germany).
Subcellular fractionation and immunoblotting: HT 29 and SW 480 cells were either incubated with 1000 μM of metamizole, MAA, acetaminophen or a combination of the reagents for 24 h and 48 h. Standard growth medium was used for negative control, staurosporine (1 μM) for positive control. Cells were washed twice with cold DPBS and collected by centrifugation at 4000 rpm at 4°C for 10 min. Lysates were then resuspended in RIPAE-Buffer (5 ml Triton X100, 190 mg EDTA, 0.5 g SDS, 2.5 g Deoxycolid Acid, 500 ml DPBS, proteinase inhibitors) for 15min and centrifuged at 13.000 rpm for 30 min. Supernatants were transferred to new cups and incubated on ice. 30 μg of total lysates were analyzed by SDS-PAGE and blotted onto nitrocellulose. After protein extraction and gel transfer, membranes were washed in TBS washing buffer and incubated with peroxidase-conjugated secondary antibodies. Immunoreactive proteins were visualized by means of an enhanced chemiluminescence detection system (Western Blotting Detection Reagent, GE Healthcare, Freiburg, Germany).
Statistical analysis. Results are expressed as mean±SD. For comparison between mean values the non-parametric Mann-Whitney-U-test was used. Differences were considered statistically significant at p<0.05. IBM SPSS Statistics (V 21; IBM New York, NY, USA) and Excel 2010 (Microsoft, Redmond, WA, USA) packages were employed for statistical analysis.
Results
Cell proliferation. After incubation with 1,000 μM acetaminophen cell proliferation was significantly reduced in SW 480 cells (Figure 1A). Lower concentrations of acetaminophen did not affect SW 480 cell growth. In HT 29 cells, only 1 μM acetaminophen caused a slight but significant inhibition of cell proliferation. 1 μM metamizole slightly but significantly enhanced cell proliferation rate in SW 480 cell by 5%±16% but inhibited growth of HT 29 cells by 13%±1.8% (Figure 1B). MAA only revealed antiproliferative activity at 500 μM in HT29 cells (Figure 1C). All other concentrations of MAA and metamizole did not change proliferation rate (Figure 1).
Apoptosis analysis. Incubation with 250 μM of the test agents: Incubation with 250 μM metamizole enhanced the number of apoptotic cells after 24 h compared to the untreated control in HT 29 (14,3 %±3,5% to 17,0%±2,6%) and SW 480 (17,6%±3.4% to 21,8%±1.6%) cell lines. The amount of 250 μM MAA did not affect the apoptosis rate in both cell lines, 250 μM acetaminophen only induced apoptosis in SW 480 colon carcinoma cells after 16 h and 24 h treatment time compared to untreated control. The combination of 250 μM acetaminophen and metamizole significantly enhanced apoptotic cells in both cell lines after 16 and 24h (HT 29 14,3%±3,5% to 17,9%±4,7% and SW 480 17,6%±3,4% to 22,2%±2,3%) (data not shown).
Incubation with 1 mM of the test agents: In SW 480 cell line a significant induction of apoptosis was detected after 24 h and 48 h incubation with 1 mM acetaminophen, metamizole as well as the combination of 1 mM acetaminophen with 1 mM metamizole. However, treatment with MAA for 24 and 48 h did not affect the rate of apoptosis (Figure 2A). In HT 29 colon cancer cells a significant enhancement of apoptosis was caused by 1mM acetaminophen or metamizole after 24 h and 48 h treatment time. Also, an incubation with MAA or the combination of acetaminophen and metamizole induced apoptosis after 24 h (Figure 2B). The positive control staurosporine increased apoptosis rate in both cell lines (data not shown).
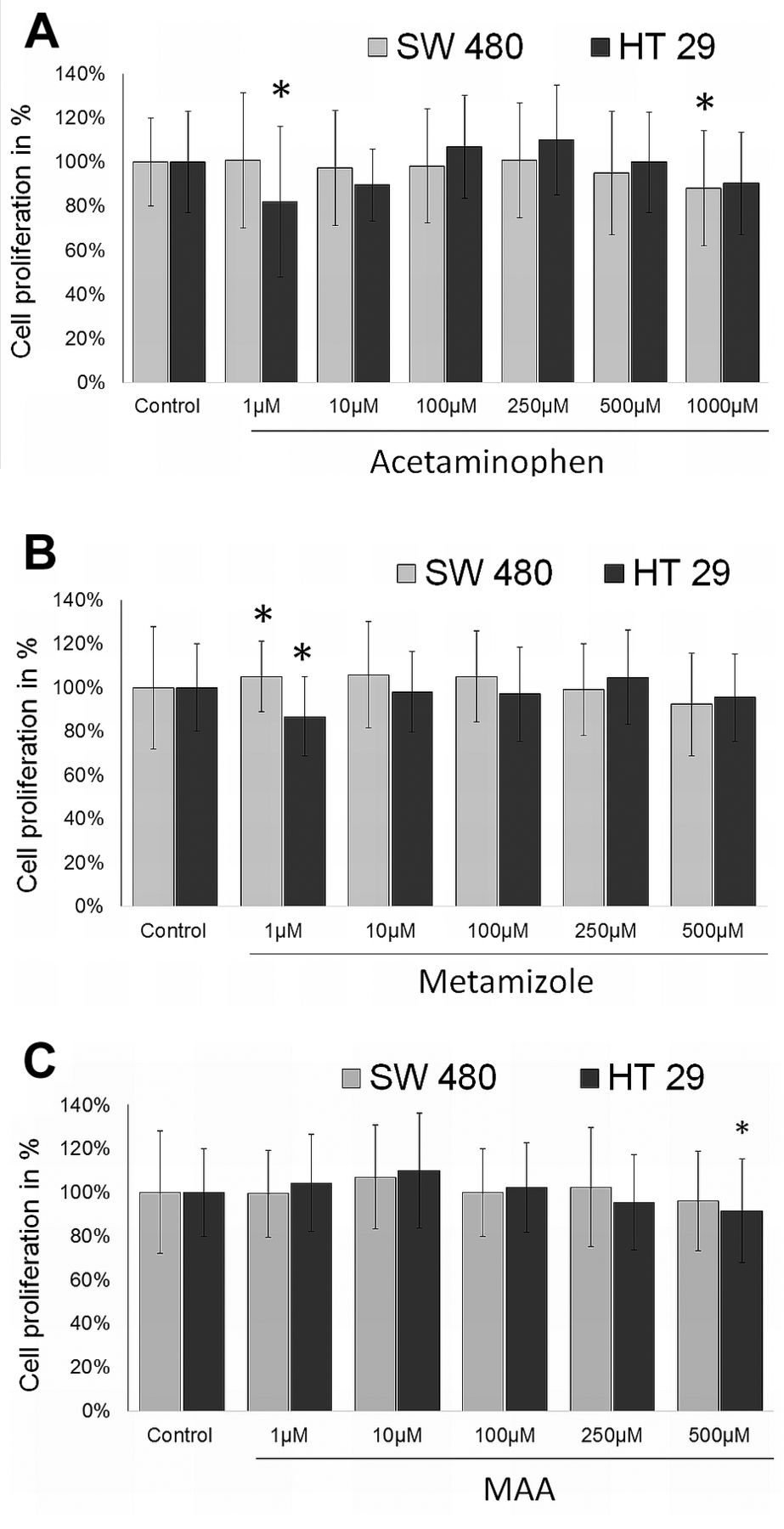
Cell proliferation. Effect of acetaminophen (A), metamizole (B) and MAA (C) on SW480 and HT29 colon carcinoma cell proliferation after 48h incubation with 0-1000 μM. Proliferation rate was detected by BrdU uptake. *Statistical significance at p<0.05 compared to untreated control.
Western blot. Western blot analysis was performed after 24 h and 48 h treatment with the test compounds or vehicle. In SW 480 cells a reduction of caspase 3 full length and caspase 8 full length expression was seen after treatment with all test agents. Also, a decrease in procaspase 9 expression occurred after treatment with metamizole, acetaminophen and the combination of acetaminophen and metamizole (Figure 3A). Also in HT29 colon cancer cells all test agents caused a reduction in caspase 3 full length expression level. Caspase 8 full length expression decreased after incubation with acetaminophen and procaspase 9 expression decreased after therapy with acetaminophen, MAA or the combination of both agents (Figure 3B).

Apoptosis analysis. Effect of 1000 μM acetaminophen (AC), metamizole (M), MAA or the combination of metamizole + acetaminophen (M-AC) or MAA and acetaminophen (MAA-AC) on apoptosis rate in SW 480 (A) and HT29 (B) colon cancer cell lines after 24 and 48h incubation time. *Statistical significance at p<0.05 compared to untreated control.
Discussion
Acetaminophen and metamizole are well established non-opioid analgesic agents, which are often used for multimodal postoperative pain therapy. However, the mechanism of action remains unknown. For both agents, interaction with the cannabinoid and opioid system as well as an inhibiton of the cyclooxygenase are discussed (10, 11). The pyrazolone derivate metamizole is rapidly hydrolysed to its active metabolite 4-methylaminoantipyrine (MAA). After oral administration of 1g metamizole maximal plasma concentration of 17.3±7.5 mg/l was measured within 1-2h. Peak plasma concentrations of MAA of 62.2±15.9 mg/l (~210-350 μM) were obtained after i.v. injection of 1g metamizole (12).

Western blot analysis. Western blot analysis for caspase 3 full length, caspase 8 full length, procaspase 9, cytochrome c and ß-actin. SW480 (A) and HT 29 (B) cells were treated with 1mM of metamizole, acetaminophen, MAA or the combination of metamizole and acetaminophen or MAA and acetaminophen for 24 and 48h. Standard growth medium was used for negative control.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Apoptosis signaling pathway. Extrinsic pathway: activation of caspase 8 after ligand binding to death receptor. Intrinsic (mitochondrial) pathway: activation of caspase 9 after release of cytochrome c.
In a pharmacological study, patients undergoing heart surgery received 1 g acetaminophen every 6h either intravenously (i.v.) or rectally. Peak plasma concentrations after the first i.v. administration was 95±36 μM, after the 4th i.v. dose concentrations of 210±84 μM were measured, the concentration after the 4th rectal dose was 69±44 μM (13).
One important characteristic of solid cancer cells is the unlimited cell proliferation. While in healthy tissue cell growth is regulated by a variety of factors, tumor cells develop several mechanisms for evading growth control (14). Results of preclinical studies show that several analgesic agents such as opioids and NSAIDs can affect cancer cell proliferation rate (8, 15, 16). In this study, we detected only slight effects on cell proliferation by acetaminophen, metamizole and MAA. Metamizole inhibited A549 lung cancer cell proliferation in a dose dependent manner while no induction of apoptotic cell death was detected (17). Clinical concentrations of acetaminophen inhibited cell growth in an epithelial ovarian tumor cell line in vitro (18). Vad et al. reported a selective toxicity of acetaminophen towards melanoma cell lines, while no toxic effects were detected in other tumor entities such as colon and pancreas cancer cell lines (19). However, stimulation of cell proliferation by acetaminophen was observed in lung carcinoma cells (20), kidney fibroblasts (21) and estrogen responsive breast cancer cells (22). Interestingly, co-incubation with acetaminophen enhanced sensitivity to chemotherapeutic agents and ionizing radiation (23-25).
Apoptosis is a highly selective process, which plays a crucial role in physiological as well as pathological processes. The programmed cell death can be initiated by two pathways: the intrinsic (or mitochondrial) pathway and the extrinsic (or death receptor) pathway (Figure 4) (26). The extrinsic pathway is started by binding of a ligand to the death receptor. Consecutively, procaspase 8 is cleaved into its activated form. Internal stimuli like hypoxia and irreparable genetic damage can trigger the intrinsic pathway. Due to an increased mitochondrial permeability, the pro-apoptotic molecule cytochrome c is released into the cytoplasm. This leads to the activation of caspase 9. The mitochondrial pathway is controlled by proteins of the Bcl 2 family, which are located at the mitochondrial membrane. The sensible balance between pro-apoptotic (Bax, Bad and Bak) and antiapoptotic proteins (Bcl-2, Bcl-xl) is often altered in cancer cells. With the activation of the effector caspase 3 by the initiator caspases 8 and 9 the common pathway of apoptosis is initiated.
In this study we show an induction of apoptotic cell death in clinically relevant doses of metamizole and acetaminophen alone and in combination in HT 29 and SW 480 colon cancer cell lines. To elucidate the underlying signaling pathway, drug concentrations were increased to 1 mM. In this concentration, a significant induction of apoptosis occurred in SW 480 and HT 29 cells after 24 and 48 h incubation with metamizole and/ or acetaminophen. As a slight decrease of caspase 9 full length expression level after 48h incubation with acetaminophen was detected in both cell lines, the mitochondrial pathway might be involved in acetaminophen induced apoptosis. The coincidental reduction of caspase 8 full length expression level suggests also an involvement of the death receptor signaling pathway.
Posadas et al. showed a potentiation of the staurosporine induced cell death by 1 mM acetaminophen in the human neuroblastoma cell line SH-SY5Y. However, incubation with acetaminophen alone did not cause caspase 3 activation nor cytochrome c release (27). In contrast, higher concentrations (2 mM) of acetaminophen induced apoptosis via the mitochondrial pathway (28). Several studies detected an induction of apoptosis via the mitochondrial pathway in different hepatoma cell lines (21, 29, 30), as well as in neuroblastoma (28). Metamizole attenuated ischemic brain injury and inhibited neuronal cell death via the inhibition of the mitochondrial apoptotic pathway in vivo and in vitro. (31) In three human leukemia cell lines, metamizole revealed cytoprotective effects in low concentrations (<300 μM), while in higher concentrations induction of apoptosis was detected (32). Garcia Martinez et al. detected no induction of apoptosis by 75 μM metamizole or MAA (33). At concentrations of 10 and 100 μM metamizole induced apoptosis in osteoblast like cells and at 1,000 μM necrotic cell death was observed (34).
Conclusion
Acetaminophen, metamizole and MAA only caused slight changes in cell proliferation in both colon carcinoma cell lines investigated. Results of apoptosis analysis as well as western blot analysis indicated an induction of apoptotic cell death by metamizole and acetaminophen in both cell lines. Due to consumption of procaspase 3, 8 and 9, the induction of apoptosis by acetaminophen seems to follow both an extrinsic and intrinsic signaling pathway. Further studies are warranted to investigate if the combination of acetaminophen and metamizole leads to an augmentation of anticancer effects.
Acknowledgements
The Authors would like to thank Renate Lange, Sigrid Bamberger, Regina Lindner, Marion Schindler, Ruth Spaeth, Theresa Rost and Daniel Potschka (all Department of Anesthesiology, University of Regensburg, Germany) for their excellent technical support. The Authors would also like to thank the Regensburger Forschungsfoerderung in der Medizin (ReForM) program of the University of Regensburg for financial support.
Footnotes
Conflicts of Interest
None.
- Received November 13, 2017.
- Revision received November 27, 2017.
- Accepted November 28, 2017.
- Copyright© 2018, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved