

Abstract
Background/Aim: This in vitro study analyzed the impact of heparins on expression of chemokines in human endometrial adenocarcinoma cell lines. Materials and Methods: Cell lines were incubated with unfractionated heparin (UFH), low molecular weight heparins (LMWH) and fondparinux under hypoxic and normoxic conditions. Chemokine (C-X-C motif) ligand 8 (CXCL8), CC-chemokine ligand 2 (CCL2) and CCL5 were detected by enzyme-linked immunosorbent assays and real-time reverse transcriptase-polymerase chain reaction and cell viability by fluorometric assay. Results: Different adenocarcinoma cell lines had distinct patterns of chemokine expression. UFH attenuated the secretion of CXCL8 and CCL2, and enhanced that of CCL5. The observed effects of heparin were in addition to the anti-coagulatory properties of heparin and dependent on molecular size and charge. Conclusion: UFH has selective modulating effects on the secretion of CXCL8, CCL2 and CCL5 in different endometrial adenocarcinoma cell lines. Molecular size and charge are relevant for these observed effects. By influencing the expression of these inflammatory mediators and thereby affecting the tumour microenvironment, heparins and related agents might play an essential role in the development of new therapeutic strategies.
- Heparin
- endometrial adenocarcinoma
- chemokines
- tumour environment
With an estimated incidence of 54,870 cases per year for 2015, endometrial adenocarcinoma is the most common gynaecological malignancy in the United States and in developed Western countries (1). The main hypothesis for endometrial carcinogenesis implicates an increase of mitogenic activity of endometrial cells triggered by excessive oestrogen exposure (2). However, there is accumulating evidence for other mechanisms involved in the development of endometrial cancer, mediated by inflammation and hypoxia (3, 4). Inflammation seems to influence carcinogenesis by altering the tumour microenvironment, especially the levels of cytokines and chemokines. The regular and cyclical episodes of inflammation in the human cycling endometrium can be enhanced by hormonal and genetic changes in the early development of endometrial adenocarcinoma, and thus the resulting exacerbated local inflammation contributes to the initiation and progression of endometrial adenocarcinoma (4).
The chemokines chemokine (C-X-C motif) ligand 8 (CXCL8), CC-chemokine ligand 2 (CCL2) and CCL5 are typical representatives of the tumour-supporting arm of inflammatory chemokines (4, 5). Due to its initiation of leucocyte infiltration and neovascularization, CXCL8 is a potent promotor of invasion and metastasis (6). CCL2 is also described as a very potent angiogenic chemokine. Together with CCL5, CCL2 seems to promote the migratory and invasion-related properties of tumour cells and thus might play an essential role in the metastatic process (7).
The anti-coagulatory activity of the classical anticoagulant unfractionated heparin (UFH) is based on its inhibition of factor Xa and thrombin (8). Being a mixture of polysulphated glycosaminoclycans (GAGs) with molecular weights ranging from 5 to 30 kDa, UFH has an average molecular weight of 13 kDa (9). Low-molecular-weight heparins (LMWHs) are commonly used in clinical practice. Derived from UFH by enzymatic or chemical depolymerisation, they primarily inhibit factor Xa (10). UFH can interact with many proteins containing positively charged amino acids due to their high charge density, which results in various biological activities in addition to anti-coagulatory activity (11, 12). Many in vitro studies showed several molecular and cellular actions of heparins, suggesting their beneficial effects, not only those already widely reported on inflammatory diseases but also on malignant diseases (12-14). Li et al. recently determined that pre-treatment with UFH significantly inhibited lipopolysaccharide (LPS)-stimulated CXCL8 production in human pulmonary microvascular endothelial cells (15). Heparin was also shown to inhibit CCL5-mediated migration and invasion of human hepatoma cells (16).
In contrast to these convincing data in vitro, there is much speculation on the potential benefit of heparin administration (mainly LMWH) in vivo with regard to the prolongation of survival for many patients with solid malignancies (17). Today there is no recommendation for the routine use of heparin (especially LMWH) in patients with cancer, mainly due to the fact that no sufficiently powered studies have been undertaken so far to support its routine use for improving the survival of patients with cancer (18).
Since cyclical episodes of inflammation in the human cycling endometrium may contribute to carcinogenesis of endometrial adenocarcinoma (4, 19), we examined the impact of heparin on secretion and production of chemokines in endometrial adenocarcinoma cells. Additionally, we characterized the effects we observed under normoxic and hypoxic conditions.
Materials and Methods
Cell culture and experimental conditions. Endometrial cancer cell lines (ECC-1, RL95-2, HEC-1A, KLE and AN3CA) were obtained from the American Type Culture Collection (Manassas, VA, USA). ECC-1 is a well-differentiated cell line of an endometrial adenocarcinoma; HEC-1 A and RL95-2 are moderately differentiated cell lines. KLE and AN3CA are poorly differentiated cell lines. ECC-1 cells were maintained in RPMI-1640 (Biochrom, Berlin, Germany), RL95-2 as well as KLE cells were maintained in phenol red-free Dulbecco's modified Eagle's medium: Nutrient Mixture F-12 (Gibco/Life Technologies, Karlsruhe, Germany). HEC-1 A cells were maintained in McCoy's 5A (Gibco) and AN3CA cells in MEM Earle's (Biochrom). All media were supplemented with 10% foetal bovine serum (Biochrom) and 50 μg/ml gentamycin (Ratiopharm, Ulm, Germany). For our experiments, cells were seeded in different microplates. For each cell line, a specific cell count was used: AN3CA 0.2×106/ml, ECC 1 0.2×106/ml, HEC-1 A 0.3×106/ml, KLE 0.15×106/ml and RL95-2 0.5×106/ml. The cells were incubated with the following agents: UFH (Sigma-Aldrich, St Louis, MO, USA); LMWHs dalteparin (FragminP®; Pharmacia, Berlin, Germany), enoxaparin (Clexane®; Sanofi-Aventis, Frankfurt, Germany), reviparin (Clivarin®; Abbot arzneimittel, Hannover, Germany) and fondaparinux (Arixtra®; Galaxo, Notre Dame de Bondeville, France). Cells were cultured under normoxic and hypoxic conditions. To create hypoxic conditions, cells were placed in a tissue culture incubator (MCO-18M Sanyo, Moriguchi, Japan) in an atmosphere of 1% O2 and 5% CO2 at 37°C.
For the detection of the stimulated chemokines at the protein level, different endometrial cell lines were incubated with 0.05-51.2 IU/ml UFH for 48 h. Thereafter, the protein levels of chemokines in the cell-culture supernatant of stimulated and untreated cells were determined by enzyme-linked immunosorbent assay (ELISA). For the detection of the mRNA of the specific chemokines, different endometrial cell lines were incubated with 0.05 IU/ml (KLE) and 3.2 IU/ml (other cell lines) UFH alone for 4 h. Thereafter, mRNA levels of chemokines in the cell-culture supernatant of stimulated cells were determined by semiquantitative real-time reverse transcriptase-polymerase chain reaction. Untreated cells served as controls.
Sequences of polymerase chain reaction primers used in this study.
For the experiments concerning molecular size and charge of the heparins, cells of KLE and RL95-2 cell lines were incubated with UFH, dalteparin, enoxaparin, reviparin and fondaparinux at 5 IU/ml (KLE) or 50 IU/ml (RL95-2) or at 2.5 μg/ml (KLE) or 250 μg/ml (RL95-2) for 48 h under normoxic and hypoxic conditions. The levels of chemokines in the cell-culture supernatants were then determined by means of ELISA.
The different dosages and incubation times of the mentioned agents are also indicated in detail in the corresponding figures.
Cell viability assay. The relative number of viable cells was measured using the CellTiter-Blue® assay (Promega, Madison, WI, USA) following the manufacturer's instructions. Fluorescence was recorded using a FLUOstar OPTIMA system (BMG Labtech, Offenburg, Germany).
Enzyme-linked immunosorbent assays (ELISA). Treated and untreated cell-culture supernatants were collected and analysed for CCL2, CXCL8 and CCL5 by commercially available ELISA kits (DY279 and DY278, by R&D Systems, Minneapolis, MN, USA and BMS204/3MST by Bender MedSystems GmbH Vienna, Austria). All assays were performed according to the manufacturer's instructions and as recently shown elsewhere (20).
Real-time reverse transcriptase-polymerase chain reaction (RT-PCR). Isolation of ribonucleic acid (RNA) using PeqGOLD TriFast™ reagent (PeqLab, Erlangen, Germany), reverse-transcription using the High Capacity complementary (c)DNA Reverse Transcription Kit from Applied Biosystems (Foster City, CA, USA) and semi-quantitative real-time PCR using Power SYBR® Green PCR-Master Mix (Applied Biosystems) were performed according to the manufacturers' instruction. The primers (Table I) (Life Technologies) were designed using Primer Express® Primer Design Software v2.0 (Applied Biosystems) with the resulting amplicons having an intron-overlapping sequence. PCR amplification was performed as recently described in (20).
Impact of unfractionated heparin on chemokine expression in the different endometrial carcinoma cell lines under normoxic conditions.
Statistical analysis. Each experiment was performed in triplicates or quadruplicates and repeated three times. Statistical analysis was carried out with one-way ANOVA, followed by Dunnett's and Bonferroni multiple comparison tests using GraphPad PRISM version 5 software (GraphPad, San Diego, CA, USA). The results are expressed as mean±standard error of the mean (SEM). Differences were considered to be significant when p<0.05.
Results
Secretion of the chemokines CXCL8, CCL2 and CCL5 is differentially influenced by UFH. To investigate whether UFH has an influence on CXCL8, CCL2 and CCL5 in human endometrial cancer cells, all cell lines were tested for their basal secretion of chemokines under hypoxic and normoxic conditions. HEC-1-A, RL95-2 and KLE cells exhibited basal secretion of all three chemokines under both normoxic and hypoxic conditions, whereas AN3CA and ECC-1 only basally secreted CCL5. ECC-1 cells secreted CCL5 only under normoxic conditions. Incubation with UFH, led to attenuation of CCL2 expression only in KLE cells, whilst in all other cell lines, no effect on CCL2 secretion was detected. The basal secretion of CXCL8 was enhanced in RL95-2 cells and attenuated in KLE cells after stimulation with UFH. CCL5 secretion, however, was enhanced after incubation with UFH in HEC-1A, RL95-2 and AN3-CA cells. These results are summarized in Table II.
In order to determine whether the observed effects of heparin were dose-dependent, cells were incubated with increasing dosages of UFH (0.05 IU/ml-51.2 IU/ml). CCL2 was only released by HEC-1-A, KLE and RL95-2 cells at measurable concentrations. After incubation with UFH, significant attenuation of CCL2 level under normoxic, as well as hypoxic (data not shown) conditions was observed in KLE cells: A dose of 0.05 U/ml UFH reduced the concentration of CCL2 by about 40% (Figure 1C).
CXCL8 was not detected in the supernatants of AN3CA and ECC-1 cells by ELISA, whereas HEC-1-A cells secreted this chemokine at measurable concentrations, although no influence of UFH was detected (data not shown). In KLE cells, secretion of CXCL8 was attenuated by all dosages of UFH under both normoxic and hypoxic conditions; UFH at 0.05 U/ml almost halved the concentration of CXCL8.
The secretion of CXCL8 in RL95-2 cells was influenced in a dose-dependent way by UFH: only under the influence of 0.8 to 6.4 U/ml of UFH was CXCL8 expression significantly increased in RL95-2 cells, no other dosage had any significant influence.
CCL5 was secreted by all five cell lines at detectable concentrations. The secretion of this chemokine by ECC-1 and KLE cells was not influenced by UFH (data not shown). However, stimulation of HEC-1-A, RL95-2 and AN3CA cells by UFH significantly increased the secretion of CCL5 in a dose-dependent manner. These results were observed under normoxic as well as hypoxic conditions (data not shown).
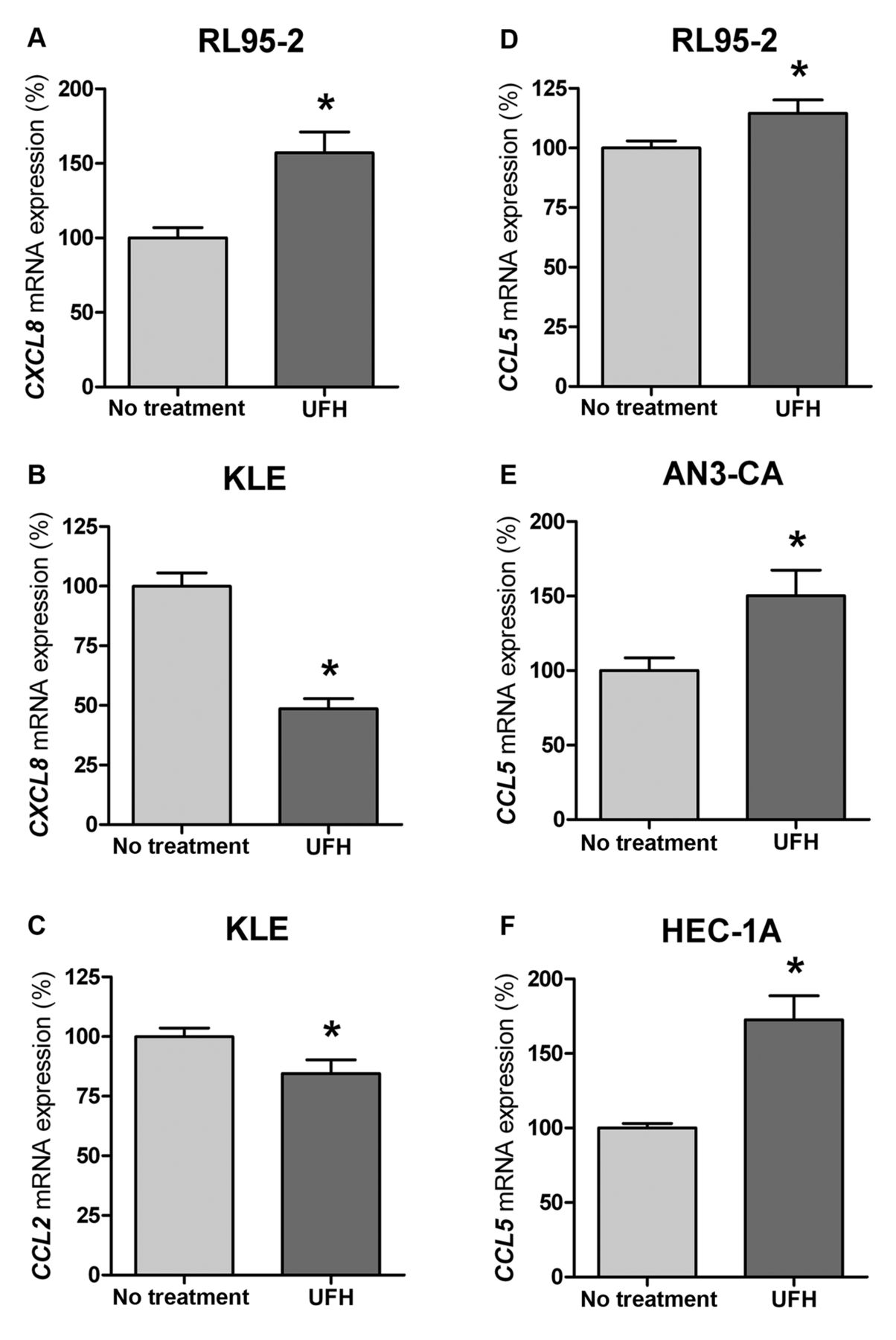
Secretion of chemokines CXCL8, CCL2 and CCL5 at the mRNA level is differentially influenced by UFH. In addition, the detected effects of UFH on chemokine secretion were based on changes at the mRNA level. In accordance with the results for protein expression (Figure 1), we observed that UFH exerted its effects (cell line- and dose-specific) on expression of the particular chemokine at the mRNA level (Figure 2). Similar results were obtained under normoxic as well as hypoxic conditions (data not shown).
Molecular size and charge are relevant for the effects of heparin on chemokine expression. To clarify the role of anti-coagulatory properties, molecular size and the amount of negative charge for the observed heparin-mediated effects, we tested LMWHs for different characteristics: In contrast to UFH, which has a molecular size of 5-30 kDa, that of dalteparin is 6 kDa, enoxaparin 3.8-5 kDa, reviparin 3.9 kDa and the pentasaccharide fondaparinux, a selective factor Xa inhibitor, only 1.7 kDa. The charge relative to size is high for UFH, dalteparin and enoxaparin, whereas for reviparin and fondaparinux, it is low. For these experiments, we evaluated the alterations in the concentration of CXCL8 in KLE cells and CCL5 in RL95-2 cells. We chose these two chemokines and cell lines because they exhibited the strongest effects of heparin (Figure 1). These cells were first incubated with the different LMWHs, dosed at the same anticoagulatory potency as UFH (0.5 IU/ml; Figure 3A and B) and (50IU/ml; Figure 3 C and D). In a second step, they were stimulated at LMHW dose adapted to the molecular size of UFH (2.5 μg/ml; Figure 4 A and B) and (250 μg/ml; Figure 4 C and D). Stimulation was performed under normoxic and hypoxic conditions.

Different endometrial cell lines were incubated with 0.05–51.2 IU/ml unfractionated heparin (UFH) for 48 h. Thereafter, the protein levels of chemokines chemokine (C-X-C motif) ligand 8 (CXCL8) (A, B), CC-chemokine ligand 2 (CCL2) (C) and CCL5 (D-F) in the cell-culture supernatant of stimulated and untreated cells were determined by enzyme-linked immunosorbent assays and related to the relative cell count of the cell viability assay. Results are standardized relative to the values for untreated cells (0 IU/ml UFH), which are set to 1 arbitrary unit (AU). Bars represent the mean±SEM; *p<0.05 compared to untreated cells.

Different endometrial cell lines were incubated with 0.05 IU/ml (B, C) and 3.2 IU/ml (A, D-F) unfractionated heparin (UFH) alone for 4 h. Thereafter, mRNA levels of chemokines chemokine (C-X-C motif) ligand 8 (CXCL8) (A, B), CC-chemokine ligand 2 (CCL2) (C) and CCL5 (D-F) in the cell culture supernatant of stimulated and untreated cells were determined by semiquantitative real-time reverse transcriptase-polymerase chain reaction. Results are standardized relative to the values of untreated cells, which are set to 100% expression. Bars represent the mean±SEM; *p<0.05 compared to untreated cells.

Cells of KLE (A, B) and RL95-2 (C, D) cell lines were incubated with unfractionated heparin (UFH), dalteparin, enoxaparin, reviparin and fondaparinux at 5 IU/ml (A, B) or 50 IU/ml (C, D) for 48 h under normoxic (A, C) and hypoxic (B, D) conditions. The levels of chemokines chemokine (C-X-C motif) ligand 8 (CXCL8) (A, B) and CC-chemokine ligand 5 (CCL5) (C, D) in the cell-culture supernatants were then determined by means of enzyme-linked immunosorbent assays. Results are standardized relative to the values of UFH-treated cells, which are set to 1 arbitrary unit. Bars represent the mean±SEM. *p<0.05 compared to no treatment; #p<0.05 compared to UFH.
As shown in Figure 3 B, when dosed at the same anti-coagulatory potency as UFH, comparable effects regarding the attenuation of CXCL8 concentration after stimulation with LMWH or fondaparinux under hypoxic conditions were detected. In addition, as shown in Figure 3A, under normoxic conditions, only UFH, dalteparin and reviparin had a reducing impact on CXCL8 concentration.
CCL5 concentration in culture supernatant from RL95-2 cells was increased after stimulation with all of the LMWHs. However, stimulation with fondaparinux had no detectable effect on CCL5 concentration. Similar results regarding CCL5 concentrations were obtained under normoxic and hypoxic conditions as shown in Figure 3C and D.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cells of KLE (A, B) and RL95-2 (C, D) cell lines were incubated with unfractionated heparin (UFH), dalteparin, enoxaparin, reviparin and fondaparinux at 2.5 μg/ml (A, B) or 250 μg/ml (C, D) for 48 h under normoxic (A, C) and hypoxic (B, D) conditions. The levels of chemokines chemokine (C-X-C motif) ligand 8 (CXCL8) (A, B) and CC-chemokine ligand 5 (CCL5) (C, D) in the cell-culture supernatants were then determined by means of enzyme-linked immunosorbent assays. Results are standardized relative to the values of UFH-treated cells, which are set to 1 arbitrary unit. Bars represent the mean±SEM. *p<0.05 compared to no treatment; #p<0.05 compared with UFH.
Using LMWHs and fondaparinux, adapted to the same molecular weight as that of UFH, comparable effects on the CXCL8 concentration under hypoxic conditions were found in stimulated KLE cells as in UFH-stimulated cells. However, under normoxic conditions, only dalteparin significantly reduced CXCL8 concentration similarly to UFH (Figure 4A and B).
An enhancement of CCL5 after stimulation of RL95-2 cells with LMWHs adapted to the same molecular weight as that of UFH was detected under hypoxic and normoxic conditions. Thereby the strongest impact was seen for UFH and dalteparin and these effects decreased with decreasing size of the molecules (enoxaparin, reviparin, fondaparinux) (Figure 4C and D).
Discussion
In the present study, we demonstrated selective modulating effects of heparin on the expression of the chemokines CXCL8, CCL2 and CCL5 in different human endometrial cancer cell line cells. The observed effects were detected at the mRNA as well as at the protein level and are beyond the anti-coagulant effects of heparin.
As demonstrated in animal studies and clinical trials, heparin has both anticoagulant properties and anti-inflammatory effects (21, 22). Whilst the benefit of heparin administered, for example, in cases of bronchial asthma and ulcerative colitis was caused by the competition for binding sites of L- and P-selectins, in other in vivo and in vitro studies, an effect of heparin on chemokine levels was also detected (23). Ding and co-workers observed in rat models of sepsis that pre-treatment of rats with UFH strongly reduced the levels of tumor necrosis factor-α (TNFα), and interleukin 1b and showed clearly that UFH can ameliorate inflammation (24). Li et al. detected that pre-treatment of human pulmonary microvascular endothelial cells with UFH significantly inhibited LPS-stimulated IL6 and CXCL8 production (15). Similar observations were recently reported by our group. We demonstrated that heparin plays an inhibitory role in TNFα-mediated effects in normal endometrial stromal cells as seen for CXCL8. Additionally, we detected that heparin enhanced the stimulation of CXCL5, CCL2, and CCL5 induced by TNFα (20). In the present study, we also observed selective modulating effects of heparin on chemokines, having either attenuating or enhancing effects depending on particular cell line and chemokine expression.
Many studies have underlined the importance of the tumour microenvironment and its soluble inflammatory mediators, including chemokines and cytokines. Such mediators affect all steps required for tumour growth and progression (7, 25). Chemokines CCL2, CCL5 and CXCL8 are believed to be typical representatives of the tumour-supporting group of inflammatory chemokines. Therefore, the effects observed in the present study of heparin on the level of secretion of these chemokines may be of anti-tumourigenic relevance (4, 5). Tumour cells are said to secrete more chemokines compared with cells of the original healthy tissue (4, 26, 27). In accordance with these reports, we observed that the well-differentiated cell line ECC-1, often used as a model for normal endometrial epithelium, secreted only CCL5 in measurable concentrations, in contrast to almost all the other cell lines studied that secreted CXCL8, CCL2 and CCL5 in measurable concentrations. The levels of chemokines secreted by the poorly differentiated KLE cell line were the highest of all these cell lines.
CXCL8, one of the most potent angiogenic chemokines (5), is secreted by three of the five examined cell lines: HEC-1A, RL 95-2 and KLE. The most poorly differentiated cell line KLE expressed the highest level of CXCL8 (data not shown). Interestingly, serum levels of CXCL8 were increased in about two-thirds of patients with advanced breast cancer and found to be associated with an accelerated clinical progression, a higher tumour load, and the presence of lymph node and liver metastases (28). In breast cancer, tumour-derived CXCL8 was also shown to induce blood-vessel formation in vivo, demonstrating the essential effect of CXCL8 in the context of tumour progression. However, little is known about the role of CXCL8 in endometrial carcinoma.
In the present study, the addition of UFH had no effect on the basal secretion of CXCL8 in HEC-1A cells, whereas RL95-2 cells exhibited a UFH-dose-dependent enhancement of the secretion of CXCL8, resulting in a bell-shaped response curve. In contrast, even minimal dosages of UFH in KLE cells sufficed to diminish the secretion of CXCL8 significantly. This diminishment was enhanced under hypoxic conditions.
Li et al. showed that UFH attenuates LPS-induced CXCL8 secretion via the phosphatidylinositol-3-kinase/protein kinase B/nuclear factor kappa B (NFκB) signaling pathway in human endothelial cells (15). Recently, our research group demonstrated an inhibitory impact of heparin on the TNFα-mediated effects on CXCL8 in human endometrial stromal cells by interference with NFκB (29). This mechanism might also be responsible for the attenuation of CXCL8 expression in KLE cells by heparin. The enhancement of CXCL8 in RL 95-2 cells after stimulation with UFH, however, was detected to have a bell-shaped response curve, thus leading to the hypothesis that the effects of heparin in these cells are mediated by forming complexes with interacting molecules (30, 31).
Expression of CCL2, an important chemokine for chemotaxis of monocytes and associated with tumour progression and metastasis, is attenuated in KLE cells after stimulation with UFH (32, 33). KLE, being the most poorly differentiated cell line, had the highest basal level of CCL2 secretion, that is in line with the observation that the expression of CCL2 and CCL5 is enhanced during breast cancer progression (26, 34).
In contrast to CXCL8 and CCL2, CCL5 is the only chemokine that is expressed in all five cell lines. Its secretion by AN3CA, HEC-1-A and RL95-2 cells after UFH stimulation was found increased at both protein and mRNA levels. An enhanced expression of CCL5 was also detected in different solid tumours, for example, in breast, prostate and endometrial cancer (4, 7, 35, 36). High levels of CCL5 are suspected to promote proliferation and invasion of carcinoma cells and seem to correlate with the stage of tumour progression in breast and cervical cancer (26, 34, 37, 38). In contrast, proliferation of the endometrial carcinoma cell lines HEC-1-A and ISHIKAWA were shown to depend on the dosage-level of CCL5 (35).
Different CCL5 functions were described to be dosage-dependent: at low concentrations, CCL5 mediates chemotaxis as a monomer or dimer; at higher concentrations, CCL5 acts as a multimer through interactions with cell surface GAGs. This complex of CCL5 and GAGs is oligosaccharide size-dependent and leads to amplification of intracellular signal transduction pathways (39). In accordance with these observations, we detected molecular size- and molecular charge-dependent effects of heparin on the expression of CCL5. Thus, we hypothesise that enhancement of CCL5 secretion after stimulation with UFH as a potent GAG might be a result of the complexation of heparin with CCL5.
In contrast to CCL2 and CXCL8, the role of CCL5 and its exact functions in tumour biology are still unclear (40). On the one hand, acting as a potent chemoattractant, it is able to favour tumour development, mainly by modulating the extracellular matrix and taking part in immune evasion mechanisms (41); on the other hand, CCL5 production is relevant to inducing a proper immune response to tumours (42). As we observed an enhancing effect of heparin on CCL5 expression, it is of great interest to examine the possible consequences of this effect in further studies.
In conclusion, UFH has a selective modulatory impact on the expression of chemokines CXCL8, CCL2 and CCL5 in human endometrial cancer cell lines. The observed effects of heparin were independent of its classical anti-coagulatory properties, but were influenced by molecular size and charge. By influencing the expression of soluble inflammatory mediators, heparin may affect the tumour microenvironment. Considering the crucial role of the tumour environment in tumour growth and progression, heparins and related agents might be of interest for the development of new antitumourigenic strategies.
Footnotes
This article is freely accessible online.
- Received January 26, 2016.
- Revision received March 15, 2016.
- Accepted March 17, 2016.
- Copyright© 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved