

Abstract
Background: Canine melanoma is the most common type of tumor in dogs. We investigated the effects of canine interferon-beta (cIFN-β)-overexpressing adipose tissue-derived mesenchymal stem cells (cATMSCs) on apoptosis and proliferation of canine melanoma cells. Materials and Methods: Expression of IFN-β in cATMSCs was confirmed using reverse transcription-polymerase chain reaction and enzyme linked immunosorbent assays. Flow cytometry was performed for cell-cycle analysis and apoptotic cell quantification of LMeC (melanoma) cells. Protein expression of cyclin D1, procaspase-3, activated caspase-3, and Bcl-2 homologous antagonist killer (Bak) was evaluated by western blot analysis. Results: Decreased proportions of cells in S- and G0/G1 phases were observed in parallel with decreased cyclin D1 expression in LMeC cells treated with cIFN-β-cATMSC-conditioned media. Protein expression of active forms of caspase 3 and Bak increased in response to treatment with cIFN-β-cATMSC-conditioned media. Conclusion: IFN-β overexpression by cATMSCs was associated with pro-apoptotic and growth-inhibitory effects on canine melanoma cells. The antitumor effects of these cells have therapeutic potential for the treatment of canine melanoma.
- Melanoma
- mesenchymal stem cells
- interferon β
- apoptosis
- cell cycle
Melanoma is the most common form of tumor in dogs that arises from melanocytes or melanoblasts. Canine melanomas most commonly occur on the skin, in the mouth, and on the digits (toes) or in association with the nail bed. Although the majority of skin melanomas are benign in dogs, oral or mucosal melanomas are considered extremely malignant tumors, with a high degree of local invasiveness and high metastatic propensity (1). Melanomas in dogs have diverse biological features based on a variety of factors, including anatomical localization, size, stage, and histological parameters (2). It is estimated that 80-100% of cases result in poor outcomes due to local invasion or metastasis to other organs, such as lymph nodes and lungs (1). Surgery, chemotherapy, and to a lesser extent radiotherapy, are widely adopted treatments. However, tumors at advanced stages are often resistant to these treatments. A combination therapy consisting of dacarbazine, carmustine, cisplatine, and tamoxifen citrate (BCDT) resulted in a 50% response rate in melanoma patients (3, 4). For this reason, novel therapies have been explored for the treatment of canine melanoma, such as immunotherapy and gene therapy, as well as conventional biological response modifiers (5-9).
Interferons (IFNs) are a group of signaling proteins (10) produced by host cells in response to pathogens such as viruses and bacteria, as well as tumor cells. Type I IFNs, comprising IFN-α and IFN-β, are a family of cytokines known for anti-viral, immunomodulatory, and anti-proliferative effects (11, 12). IFN-β is an important factor in the growth of melanoma because homozygous deletions of the class I IFN gene cluster have been demonstrated in melanomas (13, 14). Moreover, IFN-β has greater anti-proliferative and anti-invasive effects on melanoma cells compared with IFN-α (15). For this reason, Yoshida et al. (16) suggested that IFN-β gene transfer could induce apoptosis in glioma, melanoma, and renal cell carcinoma, which are known IFN-resistant tumors.
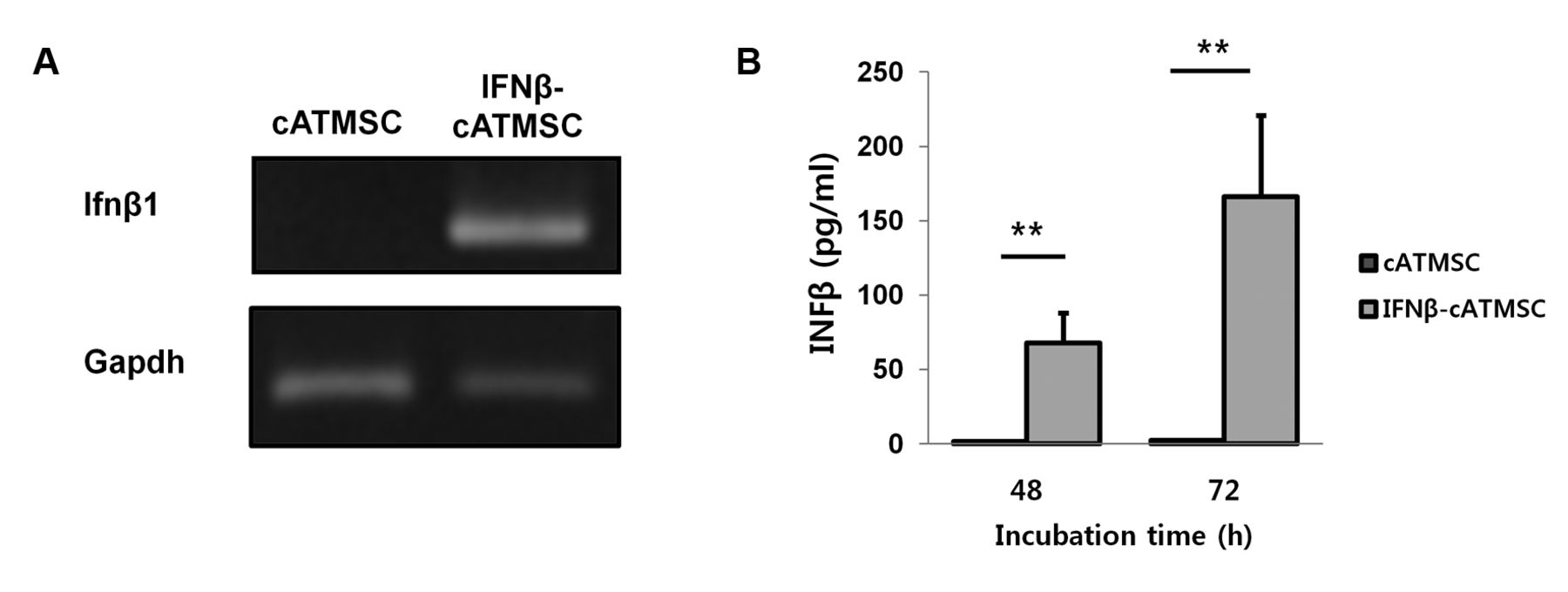
Analysis of interferon-β (IFN-β) expression in IFN-β-overexpressing canine adipose tissue mesenchymal stem cells (IFN-β-cATMSCs) by reverse transcription polymerase chain reaction (RT-PCR) and enzyme linked immunosorbent assays (ELISA). A: RT-PCR results showed significantly increased expression of IFN-β mRNA (Infb1) in IFN-β-cATMSCs after 48 h of incubation compared to control cATMSCs. B: IFN-β protein in IFN-β-cATMSCs after 48 and 72 h was quantified using ELISA. Expression of IFN-β protein was markedly increased in IFN-β-cATMSCs compared with control cATMSCs. Results shown are representative of three independent experiments with similar results. Data are expressed as the mean+standard deviation (SD). **p<0.01
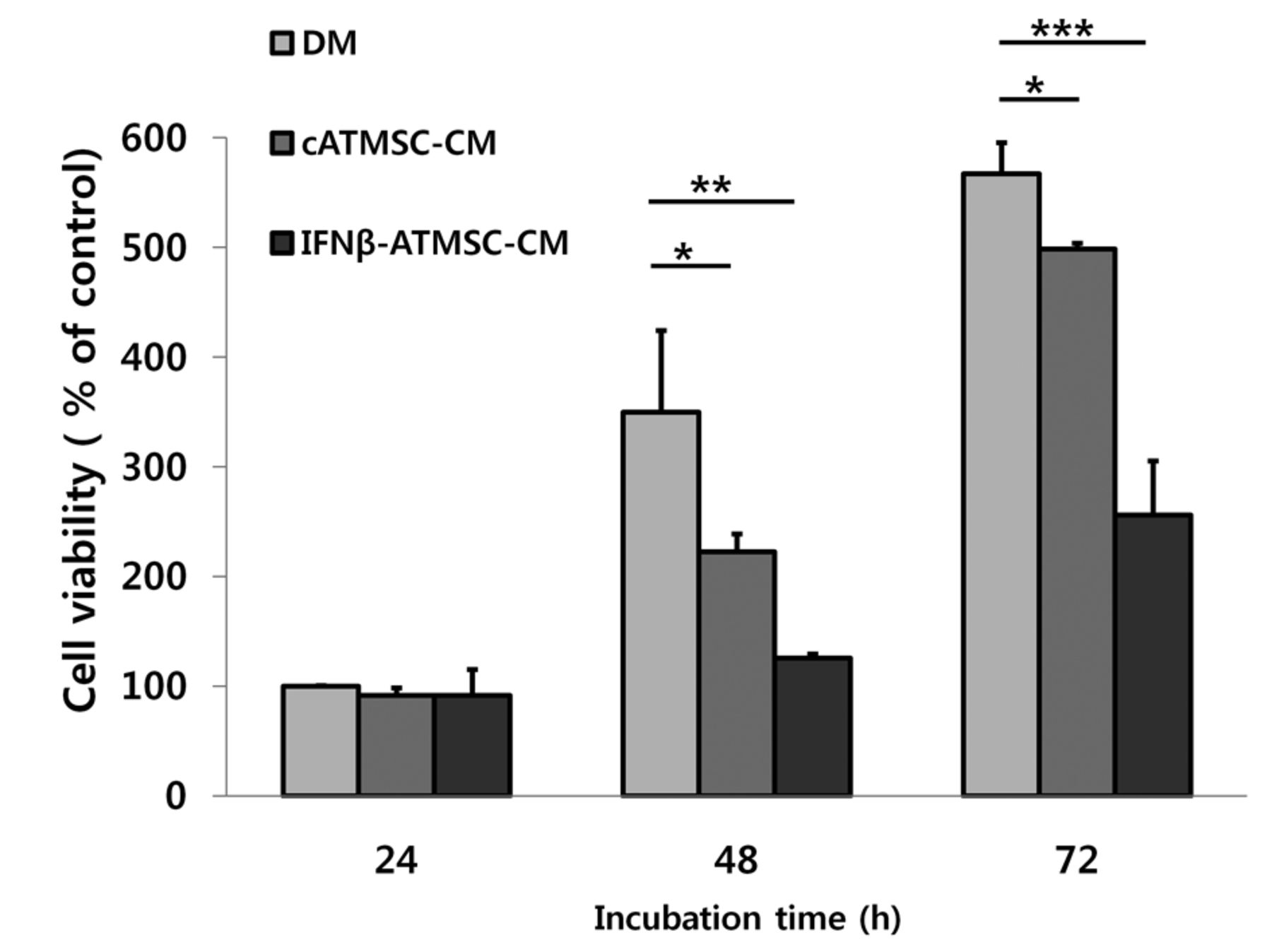
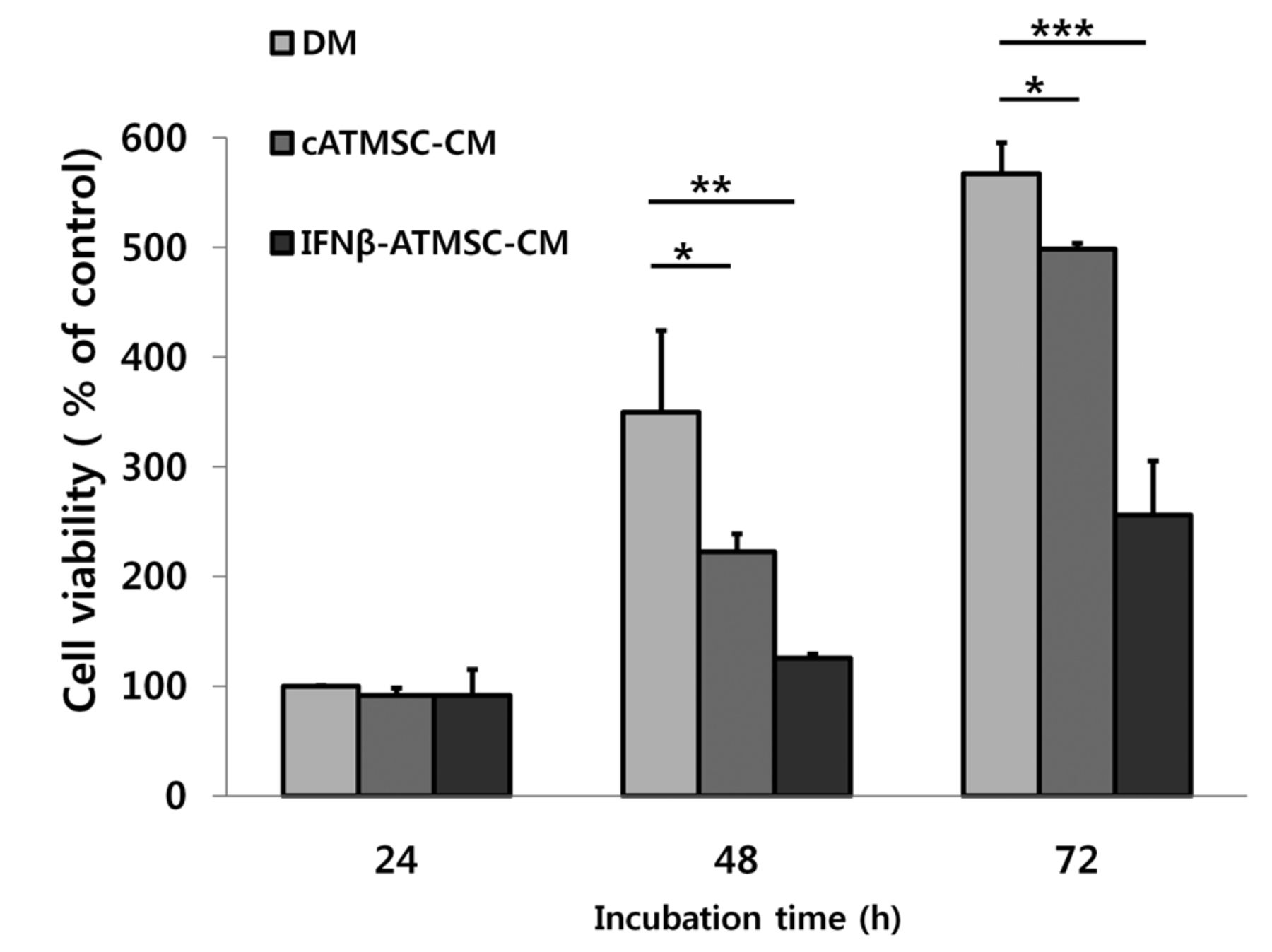
Inhibitory effect of conditioned media (CM) from canine adipose tissue mesenchymal stem cells (cATMSCs) on the growth of LMeC canine melanoma cells. LMeC cells were cultured in three types of conditioned media: defined media (DM), cATMSC-CM, and interferon-β-overexpressing cATMSCs (IFN-β-cATMSCs)-CM. Cell proliferation was assessed by 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay after 24, 48, and 72 h incubation. The CM from IFN-β-cATMSCs showed a greater inhibition of LMeC proliferation compared to that from cATMSCs. Data are expressed as the mean+SD. *p<0.05, **p<0.01 and ***p<0.001.
Mesenchymal stem cells (MSCs) have recently been used as a platform for targeted delivery of anticancer agents due to their unique characteristics and ability to accumulate at tumor sites (4, 17, 18). In addition, treatment of melanoma by xenogeneic application of human MSCs carrying anti-tumor genes was found to be safe and effective (19). We evaluated the effects of canine IFN-β-overexpressing adipose tissue-derived MSCs (IFN-β-cATMSCs) on apoptosis and growth of canine melanoma cells.
Materials and Methods
Cell culture. The LMeC canine malignant melanoma cell line was used in this study (20, 21). The cell line was kindly provided by Professor Nobuo Sasaki at the University of Tokyo. Cells were plated in high-glucose Dulbecco's modified Eagle's medium (DMEM; HyClone, Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS; HyClone), 50 U/ml penicillin, and 50 μg/ml streptomycin (HyClone).
Canine ATMSCs were obtained from canine subcutaneous fat tissue, as previously described (22, 23). Briefly, adipose tissue was collected from subcutaneous fat depots of beagle dogs using standard surgical procedures. Adipose tissue was digested overnight at 37°C with collagenase type IA (1 mg/ml; Sigma-Aldrich, St Louis, MO, USA) and then washed in phosphate-buffered saline (PBS; HyClone). Following centrifugation, the pellet was filtered through a 100 μm nylon mesh and incubated overnight in DMEM supplemented with 10%, heat-inactivated FBS at 37°C in a humidified atmosphere of 5% CO2. After 24 h, non-adherent cells were removed by washing with PBS.
Before their use in experiments, MSCs were identified based on the following cell surface markers: CD44+, CD73+, CD90+, CD31−, CD34−, and CD105−. Canine ATMSCs were maintained with low-glucose DMEM containing 10% FBS, 50 U/ml penicillin, and 50 μg/ml streptomycin. The medium was changed at 48 h intervals until the cells became confluent. When cells were >90% confluent, they were banked or serially subcultured under standard conditions.
Engineering of canine IFN-β into cATMSC. The lentiviral vector carrying the gene encoding canine IFN-β was generated as previously described (24). Briefly, the target gene amplified by PCR from canine thymus cDNA was cloned into the pLenti6/V5-D-TOPO® vector using the pLenti/V5 Directional TOPO Cloning Kit (Invitrogen, Carlsbad, CA, USA). The resulting lentiviral vector carrying Ifnb1 was sequenced to verify the correct reading frame and DNA sequence. Lentiviral particles were amplified in 293FT cells using the ViraPower™ Lentiviral Expression System (Invitrogen) according to the manufacturer's protocol.
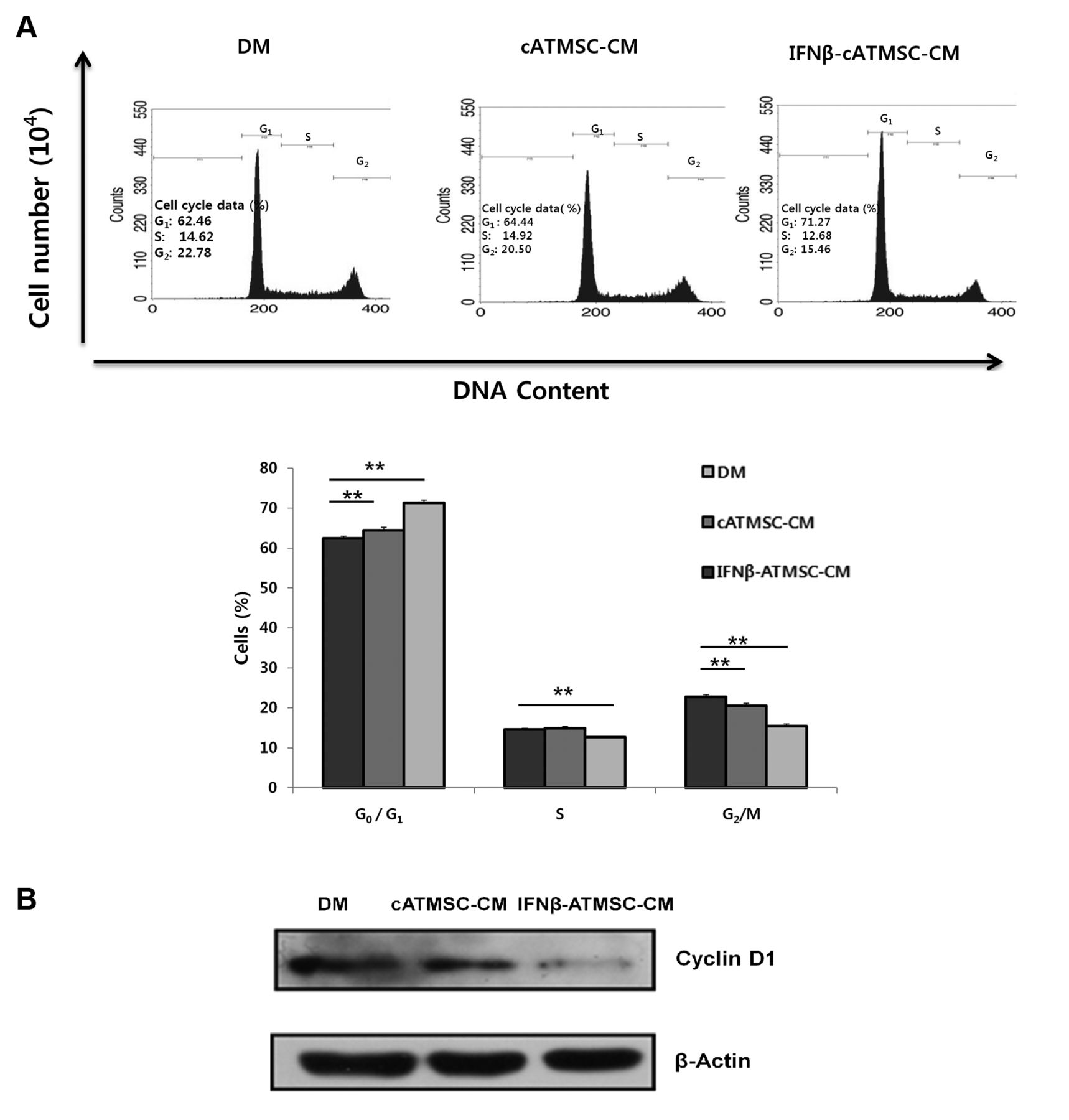
Cell-cycle arrest of LMeC melanoma cells incubated in conditioned media (CM) from canine adipose tissue mesenchymal stem cells (cATMSCs) and interferon β overexpressing cATMSCs (IFN-β-cATMSCs) for 24 h. A: The proportion G0/G1 phase LMeC cells cultured in cATMSC-CM or IFN-β-cATMSC-CM was higher than that of controls cultured in defined media (DM). LMeC cells cultured in CM from IFN-β-cATMSCs exhibited a decreased proportion of cells in S phase and G2/M phase compared to controls. The x-axis indicates DNA content and the y-axis indicates cell number. Data are expressed as the mean+SD. **p<0.01. B: Expression of cyclin D1 was decreased in the LMeC cells incubated with IFN-β-cATMSC-CM compared with expression in the DM and cATMSC-CM groups.
For transduction, the viral supernatant was added to cATMSCs at a multiplicity of infection of 5 with 6 μg/ml Polybrene (Sigma-Aldrich). After 16 h, the medium was replaced with fresh DMEM supplemented with 10% FBS. After an additional 24 h, cells were cultivated in selection medium containing 5 μg/ml blasticidin (Invitrogen) for 5 days. The cells were prepared routinely and used for in vitro and in vivo studies as low-passage cultures (passages 4-6). Successful transduction of the IFN-β-overexpressing cATMSCs (IFN-β-cATMSCs) was confirmed by reverse transcription-polymerase chain reaction (RT-PCR) and enzyme-linked immunosorbent assay (ELISA).
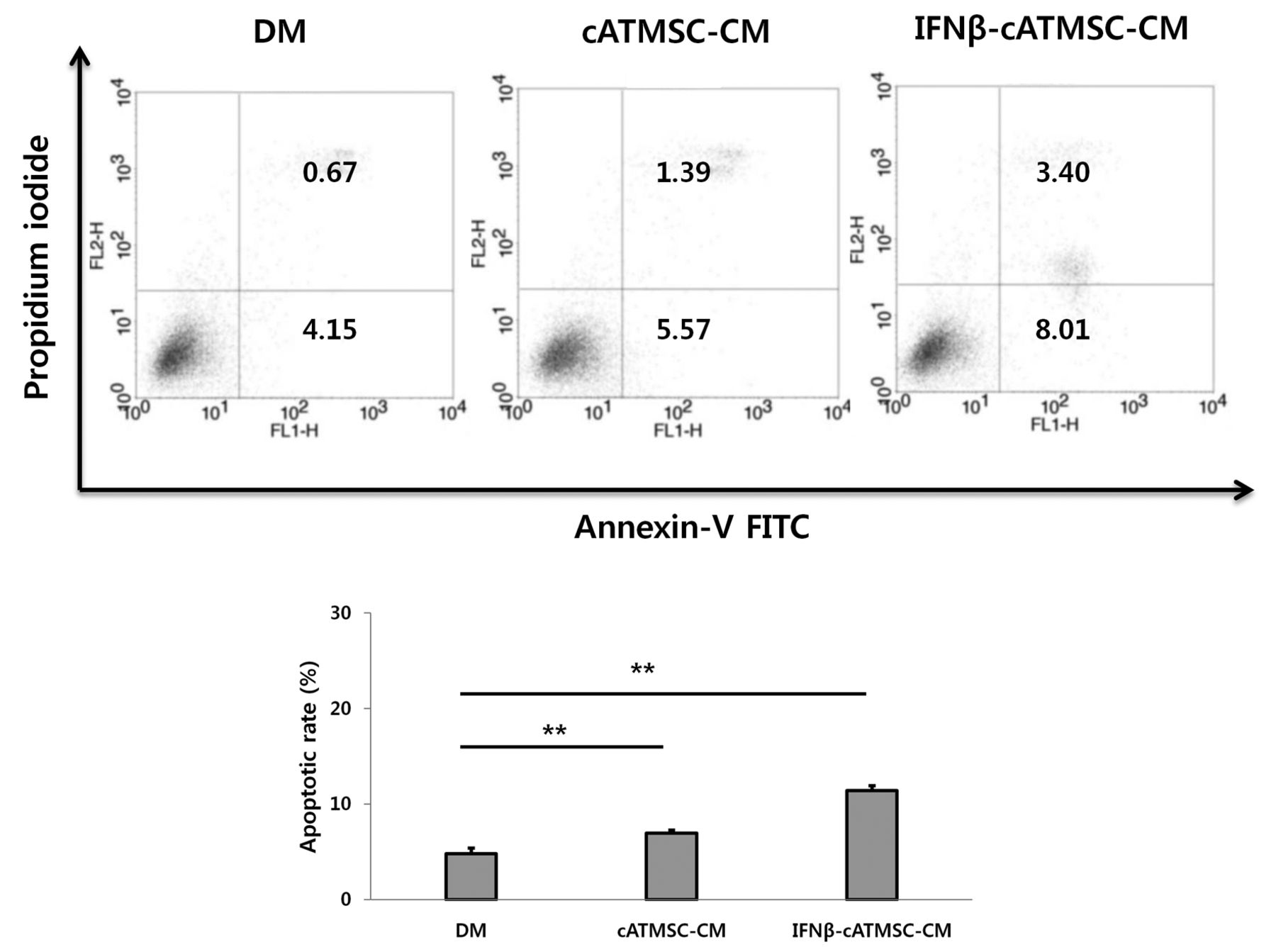
Quantification of apoptotic cells. The percentages of annexin V+/ Propidium iodide (PI)+ (apoptotic) and annexin V+/PI− (early apoptotic) cells cultured in defined media (DM), conditioned media (CM) from cATMSCs and from IFN-β-cATMSCs were 4.82%, 6.96%, and 12.41%, respectively. Data are expressed as the mean+SD. *p<0.05 and **p<0.01.
RT-PCR. Total RNA was extracted using an easy-BLUE™ Total RNA Extraction kit (iNtRON Biotechnology, Seoul, Korea). Complementary DNA templates from each sample were prepared from 3 μg of total RNA primed with oligodT primers using 400 U of Moloney murine leukemia virus reverse transcriptase (M-MLV RT) (Invitrogen), followed by 30 PCR amplification cycles (94°C for 30 s, annealing at 57°C for 30 s, and extension at 72°C for 90 s). The sense and antisense primers of each primer pair were designed to bind different exons to exclude DNA contamination: canine IFN-β (Ifnb1) (sense: 5 - GAGAGGATCCAATGACCAGTAGATGCATCCT-3, antisense: 5 - ATTTGATGTTGGCGGGAT-3, 561 bp amplicon). Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) was used as the reaction standard: sense: 5 -GGTCACCAGGGCTGCTTT-3, antisense: 5 - ATTTGATGTTGGCGGGAT-3, 209 bp amplicon). Each PCR product was analyzed using 1.5% agarose gel electrophoresis.
ELISA. The amount of IFN-β secreted by IFN-β-cATMSCs into the medium was quantified using a canine IFN-β ELISA kit (BlueGene Biotech, Shanghai, China) according to the manufacturer's protocol. IFN-β-cATMSCs were plated at 1×106 cells per well in a 10 cm plate. After 48 and 72 h, the medium was collected and centrifuged for 5 min at ×1000 g and filtered using Millex-HV 0.45 μm filters. The concentration of IFN-β in the medium was determined according to the manufacturer's protocol using recombinant IFN-β as a standard, with assays performed in triplicate. Briefly, 100 μl of medium was placed into each well; after incubation for 2 h at 37°C, wells were washed three times with wash solution. Each well was treated with 100 μl of diluted reagent B and incubated for 1 h at 37°C. After washing five times, 90 μl of substrate solution was added to each well and incubated for 30 min in the dark in a closed chamber at 37°C. After adding 50 μl of stop solution, absorbance at 450 nm was determined using a microplate reader (Bio-Rad, Hercules, CA, USA). Cytokine production was expressed as pg/ml.
Preparation of conditioned media. Conditioned media (CM) from cATMSCs and cIFN-β-cATMSCs were collected as follows: 1×106 cells of each type of cell were cultured on 10 cm plates with 10 ml of DMEM/low-glucose supplemented with 10% FBS, 50 U/ml penicillin, and 50 μg/ml streptomycin. The media was harvested following 48 h and 72 h incubations and filtered through 0.45 um filters and stored at −80°C prior to use. DMEM supplemented with 10% FBS, 50 U/ml penicillin, and 50 μg/ml streptomycin was used as defined medium (DM).
Cell viability assay. To determine the inhibitory effect of CM from IFN-β-cATMSCs on the growth of LMeC cells, a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT) assay was performed. One thousand LMeC cells were seeded in triplicate in 96-well plates and incubated for 24 h at 37°C and 5% CO2. After incubation, 100 μl of either DM, CM from cATMSCs, or CM from cIFN-β-cATMSCs were added to each well. After 24, 48, and 72 h, 20 μl of MTT (5 mg/ml) diluted with PBS was added to each well and plates were incubated for 2 h at 37°C. The supernatants were removed and the remaining formazan crystals at the bottom of each well were dissolved in 100 μl of dimethyl sulfoxide and shaken on the shaker. Absorbance was determined at 450 nm using a microplate reader (Bio-Rad).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
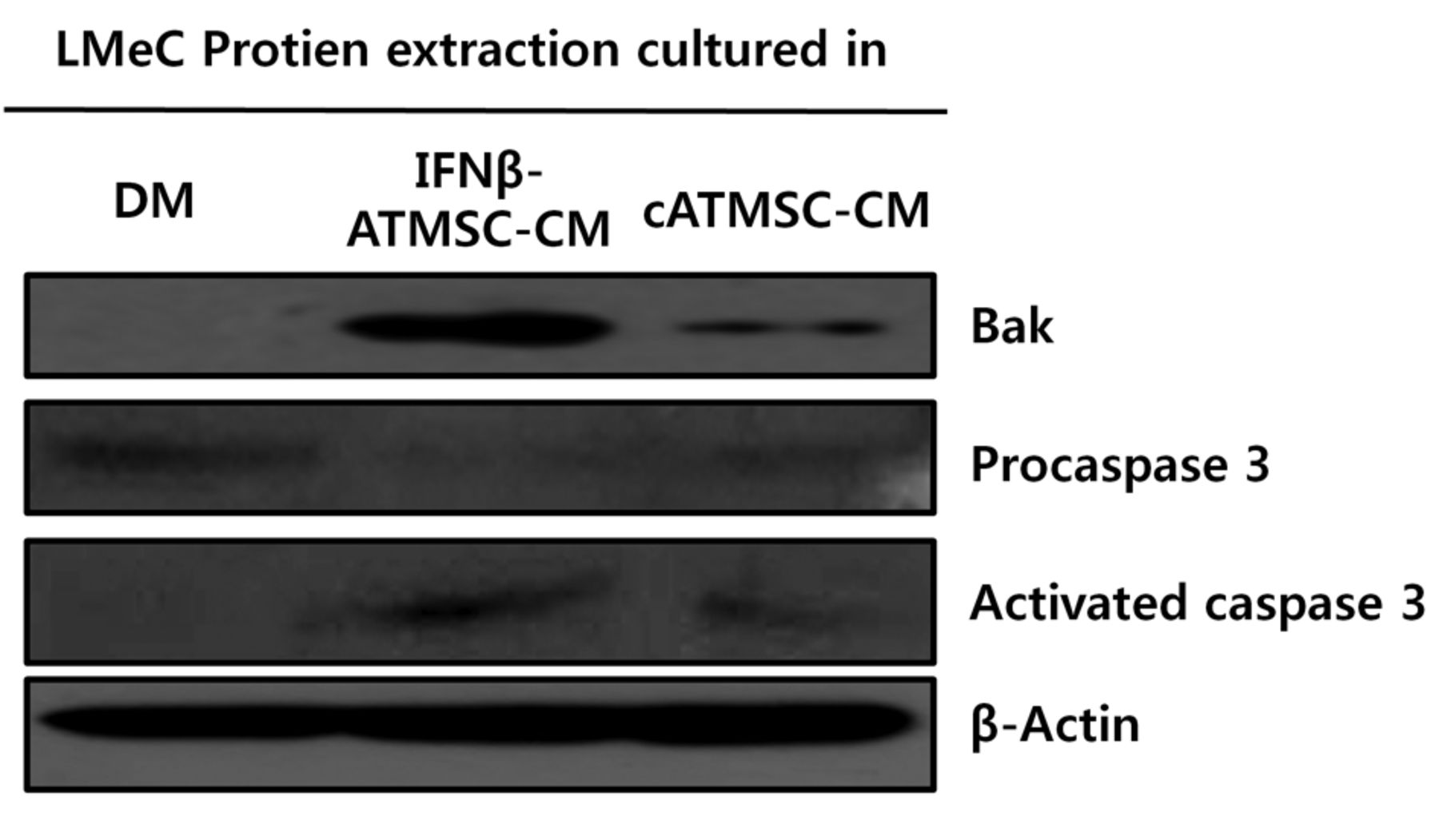
Activation of caspase 3 and Bcl-2 homologous antagonist killer (Bak) in LMeC cells incubated with conditioned media (CM) from interferon-β-overexpressing canine adipose tissue mesenchymal stem cells (IFN-β-cATMSCs). LMeC cells were incubated with defined media (DM), cATMSC-CM, and cIFN-β-cATMSC-CM for 24 h; equal amounts of extracted protein were separated by western blot. Expression of activated caspase 3 and Bak was increased in both LMeC cells on incubation with both CM sources; enhanced expression of these markers was observed in LMeC cells incubated with IFN-β-cATMSC-CM.
Cell-cycle analysis. LMeC melanoma cells (1×105 cells/well) were cultured in a 60 mm plate with DM, CM from cATMSCs, or CM from cIFN-β-cATMSCs for 24 h. Cells were then trypsinized, fixed with 70% ethanol and labeled with propidium iodide (PI) (Sigma). For cell-cycle analysis, cells were labeled with PI in the presence of RNase A (50 g/ml, 30 min, 37°C in the dark), and resuspended in PBS. Samples were run on a FACScan flow cytometer and the data were analyzed using FACScan flow cytometer software (Becton-Dickinson, FL, NJ, USA).
Apoptotic cell analysis. LMeC melanoma cells (1×105 cells/well) were cultured in a 60 mm plate with DM, CM from cATMSCs, or CM from cIFN-β-cATMSCs for 24 h. Cells were then trypsinized, fixed with 70% ethanol. Ten microliters of an annexin V stock solution was added to the cells and these were incubated for 30 min at 4°C. Next, cells were incubated with 5 μl of PI(Sigma). Cells were immediately analyzed using a FACScan flow cytometer (Becton-Dickinson).
Western blot. LMeC cells were seeded at 1×106 in a 10 cm culture plate in DMEM/high glucose supplemented with 10% FBS, 50 U/ml penicillin, and 50 μg/ml streptomycin. After 24 h, DM, CM from cATMSCs, or CM from cIFN-β-cATMSCs were added to each plate. Following an additional incubation of 24 h, all cells were harvested. Whole-cell lysates were prepared using a Proprep™ protein extraction solution (iNtRON Biotechnology). Equal amounts of protein were separated by SDS polyacrylamide gel electrophoresis, transferred to a Hybond-C nitrocellulose membrane (Amersham Pharmacia Biotech Ltd., Little Chalfont, Buckinghamshire, UK), and hybridized to an appropriate primary antibodies for cyclin D1, Bak, Procaspase3, and Activated caspase3 and horseradish peroxidase conjugated secondary antibody followed by subsequent detection by ECL. All primary and secondary antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Statistical analysis. Statistical analyses were performed using GraphPad Prism version 5 software (GraphPad Software Inc., San Diego, CA, USA). Differences were considered significant when p<0.05 compared with the corresponding control value, as determined by a one-way ANOVA with post-hoc Scheffé or Student's t-test.
Results
IFN-β expression in canine IFN-β-cATMSCs. Expression of canine Ifnb1 in cATMSCs was confirmed by RT-PCR and the protein expression was determined by an ELISA assay (Figure 1). Expression of Ifnb1 was up-regulated in IFN-β-cATMSCs. Expression of canine IFN-β protein after 48 h incubation was 67.7 pg/ml, which further increased following 72 h incubation (166 pg/ml). While the cATMSCs rarely expressed canine IFN-β, cIFN-β expression in cIFN-β-cATMSCs was abundant in the supernatant of the transduced group. These results indicate that canine Ifnb1 was successfully transduced by the lentiviral vector system.
Effect of CM from cATMSCs or IFN-β-cATMSCs on canine melanoma cells. To investigate the effect of CM from IFN-β-cATMSCs on melanoma cells, the growth of LMeC cells cultured in DM, cATMSC-CM, or cIFN-β-cATMSC-CM was assessed by the MTT assay (Figure 2). At each time point, proliferation of LMeC cells cultured in cATMSC-CM or IFN-β-cATMSC-CM was inhibited compared to cells cultured in DM. Thus, regardless of cIFN-β expression, incubation with CM resulted in growth inhibition of canine melanoma cells. In addition, CM from IFN-β-cATMSCs had an enhanced growth-inhibitory effect compared to that from cATMSCs.
Cell-cycle arrest of canine melanoma cells treated with CM from IFN-β-cATMSCs compared to CM from cATMSCs. Cell-cycle analysis was performed to investigate the effects of CM from cATMSCs and IFN-β-cATMSCs on the proliferation of canine melanoma cells (Figure 3A). Flow cytometric cell-cycle analysis showed that the proportion G0/G1 phase LMeC cells cultured in CM from cATMSCs or IFN-β-cATMSCs was higher (p<0.01) compared to that of controls. S-Phase cell-cycle arrest occurred concurrently with a decrease in the percentage of G0/G1 phase cells cultured in CM from IFN-β-cATMSCs. LMeC cells cultured in CM from IFN-β-cATMSCs exhibited a decrease in S-phase cells (p<0.01) compared to controls. A decrease in the G2/M phase was also observed in CM-treated LMeC cells (p<0.01) compared to controls. These results show that cIFN-β-cATMSC inhibits progression of canine melanoma cell cycle by preventing mitosis.
We also evaluated protein expression in cells treated with CM from IFN-β-cATMSCs after 24 h. This treatment down-regulated expression of cyclin D1, while expression in LMeC cells incubated with CM from cATMSCs was not changed (Figure 3B). This result indicates cell-cycle arrest of LMeC cells associated with IFN-β-cATMSC treatment.
Apoptosis of canine melanoma cells treated with CM of cATMSCs or INF-β-cATMSCs. Annexin V− Fluorescein isothiocyanate binds phosphatidylserine residues that are re-distributed from the inner to the outer leaflet of the cell membrane during apoptosis. After loss of membrane integrity, PI can enter the cell and intercalate into DNA (25). Figure 4 shows the percentages of annexin V− and PI− stained cells in response to incubation with CM from cATMSCs and IFN-β-cATMSCs (24 h). Viable cells are annexin V−/PI−, annexin V+/PI− cells are in the early apoptotic process, whereas annexin V+/PI+ cells have lost cell membrane integrity and have taken up PI; necrotic cells are annexin V−/PI+. The percentage of annexin V+/PI+ and annexin V+/PI− cells cultured in DM, CM from cATMSCs and CM from IFN-β-cATMSCs were 4.82%, 6.96%, and 12.41%, respectively. These data suggest that CM from cATMSCs and IFN-β-cATMSCs induced apoptosis of canine melanoma cells, with the latter treatment having a more potent effect significantly.
Activated caspase-3 and Bak in canine melanoma cells incubated with CM from cATMSCs or IFN-β-cATMSCs. To evaluate the apoptosis-inducing effects of IFN-β-cATMSCs on melanoma cells, protein expression of procaspase-3, activated caspase-3, and Bak were evaluated by western blot. As shown Figure 5, levels of procaspase-3 decreased in response to incubation with CM from either cATMSCs or IFN-β-cATMSCs compared with DM. The levels of active forms of caspase-3 increased in response to CM from IFN-β-cATMSCs. Expression of Bak was significantly up-regulated in both groups incubated with CM, with an enhanced effect observed with CM from IFN-β-cATMSCs. These results suggest that CM from IFN-β-cATMSCs promotes apoptosis compared to CM from cATMSCs of canine melanoma cells via activated caspase 3.
Discussion
The focus of this study was on induction of apoptosis and growth inhibition of canine melanoma cell lines by treatment with CM from IFN-β-cATMSCs. Canine IFN-β was overexpressed by a lentiviral system and was highly secreted from canine ATMSCs. CM from cATMSCs and IFN-β-cATMSCs had a growth-inhibitory effect on canine melanoma cells by cell-cycle arrest and induction of apoptosis. The cIFN-β-cATMSCs had a significantly growth-inhibitory effect compared with control cATMSCs.
Canine melanoma tumors originate from melanocytes but the cause is unclear. This form of tumor mainly occurs in the skin (>90%), with secondary tumors being found on the digits (5%). Melanomas are locally invasive tumors that often infiltrate deep into the bone and have a high rate of metastasis. Although local control and aggressive surgical removal are very important in melanoma treatment, it can sometimes be difficult to remove the entire tumor without removing bone. In dogs with incompletely excised or metastasized tumors, chemotherapy consisting of carboplatin or cisplatin has been used. Although novel therapies have been explored, such as small molecules that block tyrosine kinases (26, 27), as well as immunotherapies and gene therapies (5-9), other effective therapies are needed.
IFN-α and IFN-β belong to a family of cytokines that are known for anti-viral, immunomodulatory, and anti-proliferative effects (11, 12). IFN-β can affect all phases of the cell cycle, most commonly by promoting arrest at the G1 phase or by lengthening all phases (G1, G2 and S) (28). Cyclin D1 is a key regulator that influences the transition from G0/G1 to S phase. Activation of cyclin D1 decreased in response to the addition of CM from cIFN-β-cATMSCs compared to addition of DM. Thus, a compound in the CM from cIFN-β-cATMSCs potentially interfered with the cell cycle of LMeC cells by delaying the transition from G1 to S phase. Finocchiaro et al. showed that combined treatment with surgery and a cytokine-enhanced vaccine with IFN-β significantly delayed or prevented post-surgical recurrence and distant metastasis, thereby increasing disease-free and overall survival with maintenance of quality of life (3). In other studies, local expression of IFN-β induced direct tumor cell cytotoxicity both in vitro (29, 30) and in vivo (16, 31). Delivering IFNB1 by gene therapy to the tumor site also led to potent antitumor effects (24). These results are consistent with our study using IFN-β delivered by ATMSCs. Melanoma cell growth was inhibited by CM from cATMSCs or IFN-β-cATMSCs, with more potent inhibition observed using CM from IFN-β-overexpressing cATMSCs.
Inducing apoptosis is an important anti-tumor mechanism. Malignant cells commonly have defects in apoptosis and inhibition of this process can lead to tumorigenesis and resistance to therapy (32, 33). Apoptosis is a tightly regulated and highly efficient cell death system that requires interaction between various factors. Induction of apoptosis is critical for an effective response to chemotherapy and most current anticancer strategies (34).
There are intrinsic and extrinsic pathways leading to apoptosis. The extrinsic pathway is triggered following the activation of cell surface-associated death receptors such as the Fas receptor and tumor necrosis factor receptor (TNFR). The intrinsic pathway is activated by cellular stress and is regulated primarily at the mitochondrial level by the BCL2 family of proteins (35). The Fas receptors on TNFRs are ligated and an adaptor protein, Fas-associated death domain, is activated and recruited, with subsequent activation of caspase-8. Activated caspase-8 cleaves additional downstream caspases, including caspase-3. Consequently, caspases elicit the morphological hallmarks of apoptosis, including cell rounding, cytoskeletal collapse, cytoplasmic condensation, DNA fragmentation and formation of apoptotic bodies that are rapidly phagocytosed and digested by macrophages or neighboring cells (36, 37). Bak, a Bcl2 homologous antagonist/killer protein, is a pro-apoptotic member of the Bcl2 gene family that contributes to the pathogenesis of some types of cancer (19). In our study, activation of caspase-3 and Bak was associated with the apoptotic effects of CM containing canine IFN-β on canine melanoma cells. An increase in the active forms of caspase-3 in response to CM from IFN-β-cATMSCs correlated with changes in procaspase-3 levels in LMeC cells cultured CM from IFN-β-cATMSCs. These in vitro data indicate that canine IFN-β appears to promote apoptosis of these cancer cells via a caspase-dependent pathway.
Our data demonstrated that melanoma cell growth was inhibited by CM from both cATMSCs and IFN-β-overexpressing cATMSCs, with the effect being more potent in the latter. In addition, the cell cycle of melanoma cells treated with CM from IFN-β-cATMSCs was arrested, as demonstrated by a decreased proportion of S-phase cells. Active forms of caspase-3 increased in response to culture with CM from IFN-β-cATMSCs and were closely correlated with changes in procaspase-3 expression. Our findings indicate that CM from IFN-β-cATMSCs has anti-proliferative effects on canine melanoma cells. These cATMSCs secrete sufficient IFN-β to inhibit canine melanoma tumors by inducing apoptosis. The antitumor effects of cATMSC and cIFN-β-cATMSC could be therapeutically beneficial in the treatment of melanoma.
Acknowledgements
We acknowledge funding support from the Korean government through a National Research Foundation of Korea grant (number 2009-0080803). This study was partially supported by the Research Institute for Veterinary Science and BK21 Plus Program for Creative Veterinary Science Research, Seoul National University.
Footnotes
* These Authors contributed equally to this study.
Conflicts of Interest
The Authors declare that they have no competing interests with regard to this study.
- Received April 20, 2015.
- Revision received May 22, 2015.
- Accepted May 26, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved