

Abstract
Background/Aim: Tumor progression is one of the most serious issues to overcome cancer disease. As a model of inflammation-induced tumor progression, we used the regressive murine fibrosarcoma cell clone QR-32 and the progressive malignant clone QRsP-11, that was derived from QR-32. Heat shock protein beta-1 (Hspb1) is a molecular chaperone. Hspb1 plays roles in not only cell protection but also chemo-resistance, tumorigenicity and protection from apoptosis. In a recent study, we showed that Hspb1 was up-regulated in QRsP-11 compared to QR-32. Materials and Methods: We compared the expression levels of Hspb1, Hsf1 and Sox2 in QR-32 and QRsP-11 cells by means of western blotting. Results: Hsf1, a transcription factor for Hspb1 was not increased in QRsP-11. Sex determining region Y-box 2 (Sox2) is a transcription factor, reported to interact with Hspb1. Sox2 was up-regulated in QRsP-11 compared to QR-32. Conclusion: These results suggest that Sox2-Hspb1 signaling is a possible pathway responsible to tumor progression of QRsP-11.
- SOX2
- HSPB1
- HSF1
- tumor progression
Tumor progression is a very serious aspect of tumor disease. Progressive tumor cells acquire an invasive and metastatic phenotype, and this phenotype leads cancer patients to death. Progressive tumor cells exhibit high proliferation, growth, invasive activities compared to regressive benign tumor cells. Progressive and regressive murine fibrosarcoma tumor models (QR-32 clone and QRsP-11 clone) have been established by the Kobayashi group in Hokkaido University (1, 2). QR-32 is a regressive clone having weakly tumorigenic and non-metastatic activity. QRsP-11 is a progressive clone having a more tumorigenic and metastatic ability. QRsP-11 was derived from QR-32. Although 2×105 QR-32 cells result in regression after subcutaneous implantation in syngeneic mice, when they are subcutaneously co-implanted with gelatin sponge, they grow progressively. These progressively-growing cells (QRsP-11) had the ability to grow in mice without gelatin sponge.
Heat shock protein beta-1 (HSPB-1) is a chaperone of the small heat shock protein group (3). This molecule plays roles in protection from heat stress (4, 5) and apoptosis (6). Our previous studies identified HSPB1 as a key molecule playing an important role in gemcitabine (GEM) resistance by means of proteomic analysis for GEM-resistant and -sensitive human pancreatic adenocarcinoma cells (7, 8). Furthermore, many groups have reported that over-expression of HSPB1 is associated with cancer malignant phenotypes (9, 10). Our recent study reported the over-expression and nuclear translocation of lactoylglutathione lyase (Glo1) in QRsP-11 compared to QR-32. Furthermore, Hspb1, a methylglyoxal-adducted protein, was concomitantly over-expressed in QRsP-11 cells, as well.
In the present study, we reconfirmed the up-regulation of Hspb1 in QRsP-11 cells compared to QR-32 cells and investigated the expression level of Hsf1, one of transcription factors for Hspb1, in QR-32 and QRsP-11 cells. Furthermore, we investigated the expression level of Sex-determining region Y-box 2 (Sox2), a transcription factor reported to interact with Hspb1 (11), to be up-regulated in various cancer cells (12) and also to play important roles in maintaining the pluripotency of embryonic stem cell properties. SOX2 has been reported to promote migration, invasion and clonal formation of ovarian cancer cells (13) and promote oncogenic phenotypes in lung cancer cells (14). Therefore, in the present study, we investigated the expression of Sox2 in regressor and progressor cells.
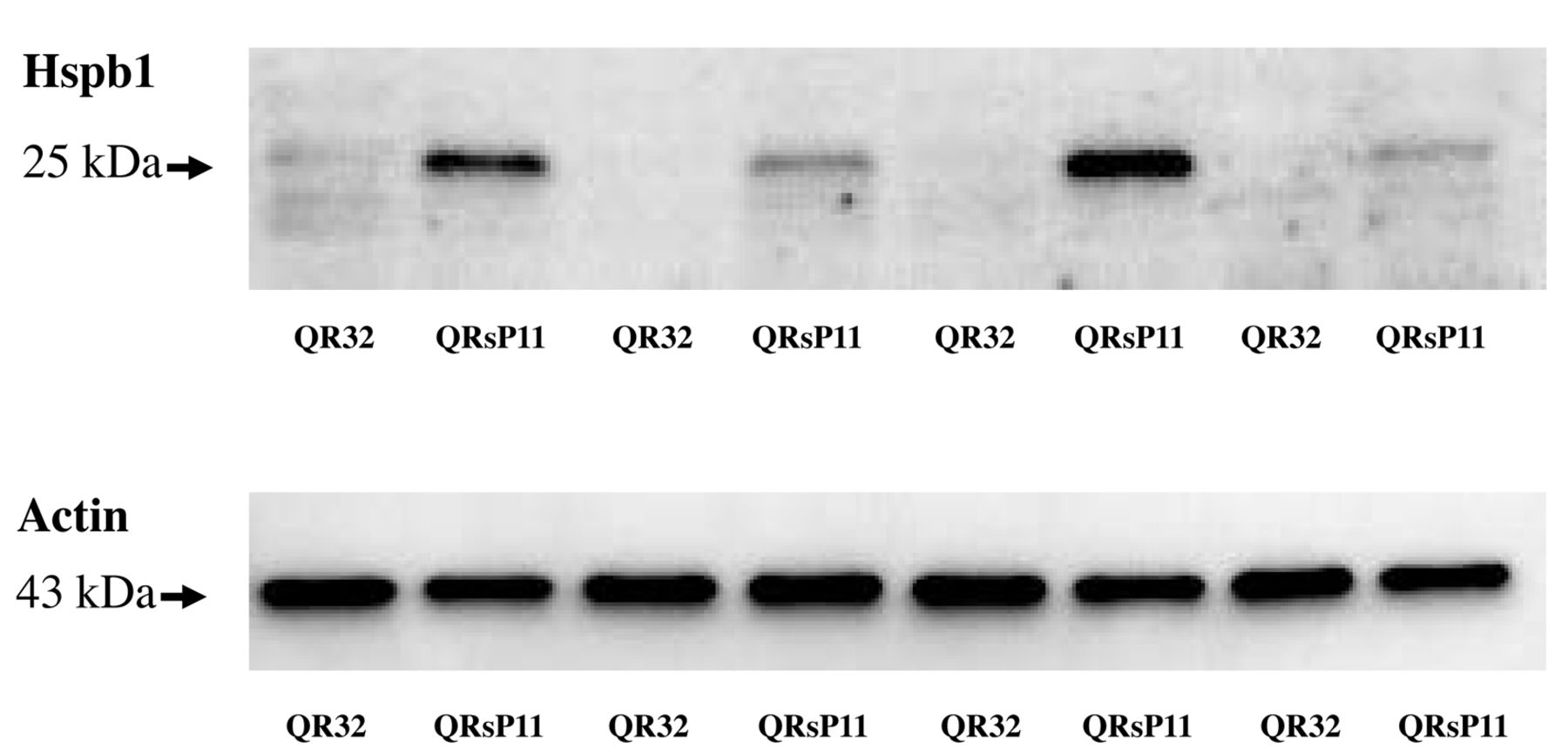
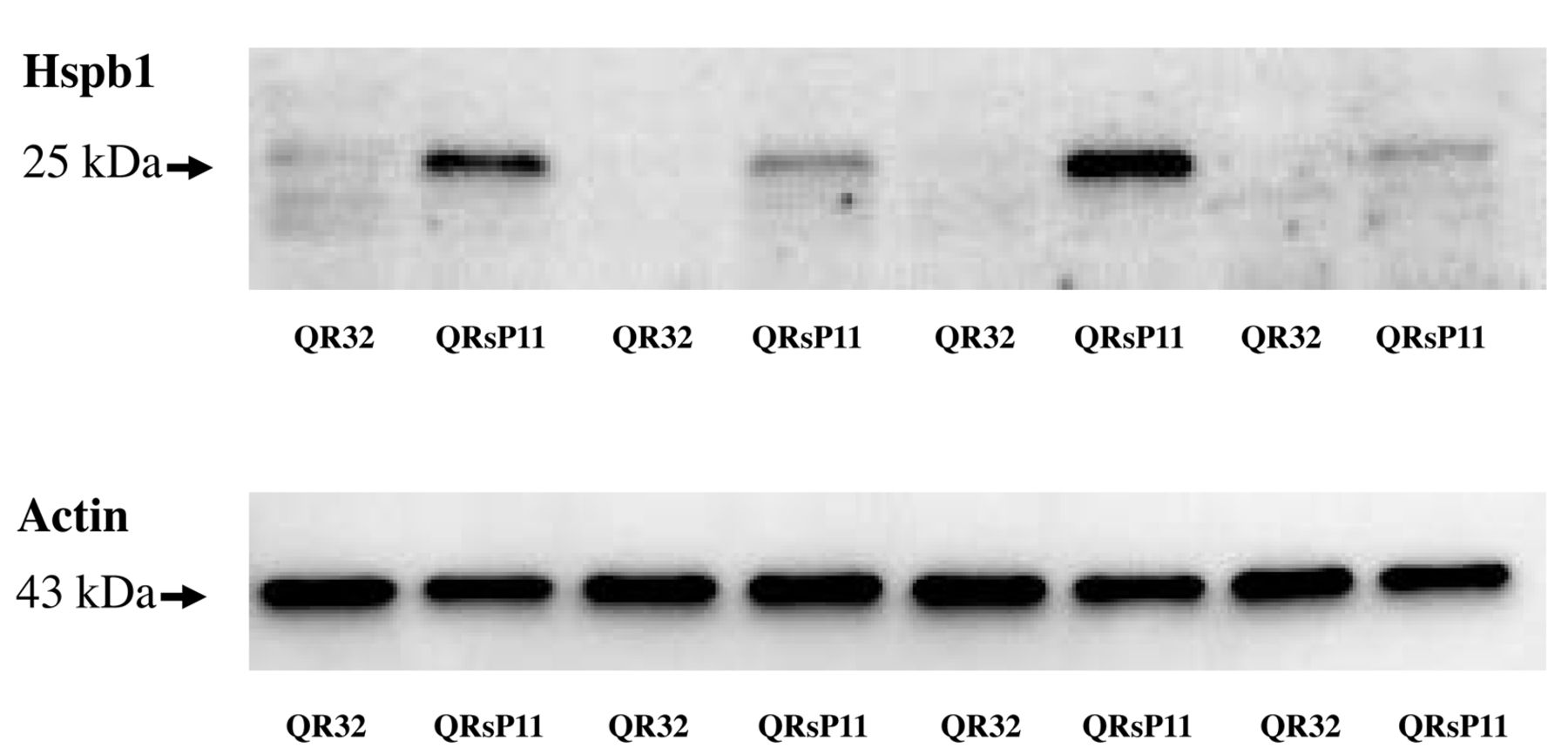
Western blot analysis of Hspb1 in murine fibrosarcoma regressor QR-32 and progressor QRsP-11 cells. Fifteen micrograms of protein were used from QR-32 and QRsP-11 cells. Bands of 27 kDa are Hspb1 and 43 kDa bands are actin as a loading control.

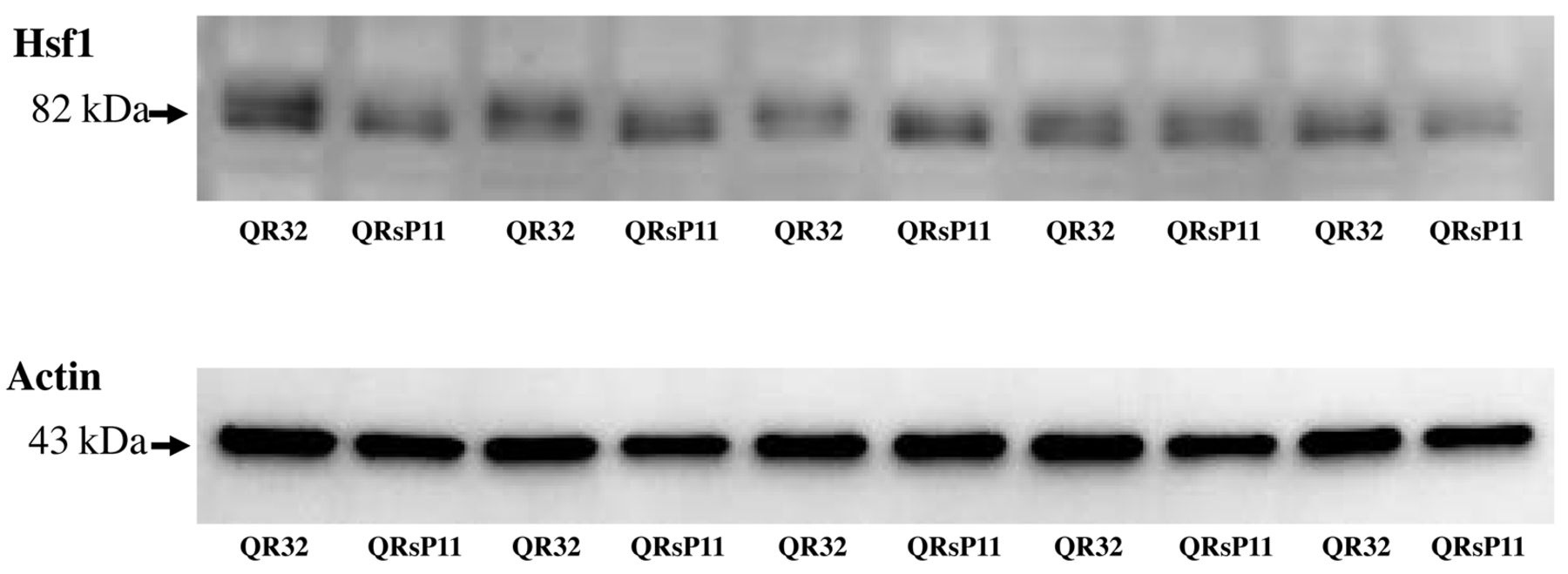
Western blot analysis of Hsf1 in murine fibrosarcoma regressor QR-32 and progressor QRsP-11 cells. Fifteen micrograms of protein were used from QR-32 and QRsP-11 cells. Bands of 82 kDa are Hsf1 and 43 kDa bands are actin as a loading control.
Materials and Methods
Tumor cell lines and culture conditions. QR-32 and QRsP-11 are murine fibrosarcoma cell lines. They were established by Kobayashi et al. (1, 2). A regressor QR-32 spontaneously regresses in syngeneic mice but, when it was co-implanted with gelatin sponge, it could grow. From this tumor, a progressor QRsP-11 was established showing strong tumorigenicity. Cells were maintained in Eagle's minimum essential medium supplemented with 10% fetal bovine serum, sodium pyruvate, non-essential amino acids and L-glutamine at 37°C.
Sample preparation. Cells were lysed in chilled lysis buffer [50 mM Tris-HCl (pH 7.5), 165 mM sodium chloride, 10 mM sodium fluoride, 1 mM sodium vanadate, 1 mM phenylmethylsulfonyl fluoride, 10 mM EDTA, 10 μg/ml aprotinin, 10 μg/ml leupeptin and 1% NP-40] on ice. After centrifugation at 21,500×g for 30 min at 4°C, the supernatants were used (15).
Western blot analysis. Fifteen micrograms of total cell lysates were subjected to sodium dodecyl sulphate-polyacrylamide gels electrophoresis (SDS-PAGE) (5-20% gradient; Bio-Rad, Hercules, CA, USA) followed by transferring onto polyvinylidene fluoride (PVDF) membranes (immobilon; Millipore, Bedford, MA, USA). The membranes were blocked with Tris-buffered saline (TBS) containing 5% skimmed milk overnight at 4°C (16). Then, the membranes were incubated with primary antibody against Hspb1 (goat polyclonal antibody to HSP27; sc-13132; dilution 1:200; Santa Cruz, Santa Cruz, CA, USA), Sox2 (goat polyclonal antibody to SOX2, sc-17320; dilution 1:1,000; Santa Cruz), Hsf1(rabbit monoclonal antibody to HSF1; dilution 1:1,000; Cell Signaling Technology, Danvers, MA, USA) and actin (goat polyclonal antibody to actin, sc-1616; dilution 1:1,000; Santa Cruz) at 4°C overnight and, then, incubated with the secondary antibody conjugated with horseradish peroxidase (dilution 1:10,000; Jackson ImmunoResearch Laboratories Inc., West Grove, PA, USA) for 1 h at room temperature after washing three times with TBS containing Tween-20 and once with TBS. Protein bands were detected by using Image Reader LAS-1000 Pro (Fujifilm Corporation, Tokyo, Japan) after treated with a chemiluminescent reagent (ImmunoStar Long Detection; Wako, Osaka, Japan) (17).

{kind=link}
{kind=link}
{kind=link}
Western blot analysis of Sox2 in murine fibrosarcoma regressor QR-32 and progressor QRsP-11 cells. Fifteen micrograms of protein were used from QR-32 and QRsP-11 cells. Bands of 35 kDa are Sox2 and 43 kDa bands are actin as a loading control.
Results
Expression of Hspb1 in QR-32 and QRsP-11. At first, we tried to reconfirm the up-regulation of HSPB1 in QRsP-11 compared to QR-32 as reported in our previous study (18). Although regressive clone QR-32 cells exhibited very faint bands of Hspb1, progressive clone QRsP-11 cells exhibited a strong band of Hspb1 (Figure 1).
The expression of Hsf1 in QR-32 and QRsP-11. HSF1 is the most famous transcription factor for HSPB1 (19). Since Hspb1 was up-regulated in QRsP-11, we speculated an up-regulation of Hsf1 in QRsP-11 cells. However, QRsP-11 did not exhibit up-regulation of Hsf1. Furthermore, the molecular size of the bands seemed to be lower in QRsP-11 cells (Figure 2). These results show that Hsf1 is not only not up-regulated, but also inactivated in QRsP-11 compared to QR-32.
The expression of Sox2 in QR-32 and QRsP-11. Since Hsf1 was not up-regulated in QRsP-11, we focused on a transcription factor, Sox2, as Hspb1 has been reported to interact with Sox2 (20). QRsP-11 exhibited high expression of Sox2 but QR-32 exhibited no expression of Sox2 (Figure 3).
Discussion
Heat shock protein beta-1 (Hspb-1) is a member of the small heat-shock protein family exhibiting chaperon-like activity (21-23). Outside the field of chaperone function, however, the roles in tumor malignant progression, increased tumorigenicity, treatment resistance and apoptosis inhibition have been a focus of attention (24, 25). In our inflammation-induced tumor progression model, it was shown that Hspb1 was up-regulated in progressor QRsP-11 cells compared to QR32 (26), a finding that was herein reconfirmed. In the present study, we further assessed the expression level of the molecules reported to associate to Hspb1.
Heat shock factor 1 (HSF1) is a transcription factor, which plays a key role in expression and activity of heat shock proteins, including HSPB1. HSF1 is the most important transcription factor to regulate heat shock response (19). Recently, some groups showed the positive relation between HSF1 and malignant transformation of tumors. Heimberger et al. reported that HSF1 plays a role in the pathogenesis of malignant myeloma (27). Toma-Jonik et al. reported that active HSF1 facilitated migration of the melanoma cells (28). Furthermore, Xia et al. reported that HSF1 supported drug-resistance of pancreatic cancer cells positively (29). Therefore, we speculated the up-regulation of Hsf1 in QRsP-11 compared to QR-32. However, Hsf1 was not found to be over-expressed in QRsP-11 compared to QR-32. Furthermore, position of bands of Hsf1 in QR32 exhibited higher than those of QRsP-11. Although we did not confirm the phosphorylation of Hsf1, the above band shift of Hsf1 may be due to phosphorylation. This means Hsf1 is not only up-regulated but also inactivated in QRsP-11 compared to QR-32 (30).
Acknowledgements
This work was supported in part by Grants-in-Aid from the Ministry of Education, Science, Sports, and Culture of Japan (no. 24501352 to Yasuhiro Kuramitsu). Immunoblot detection by LAS-1000 was performed at the Gene Research Centre of Yamaguchi University.
- Received February 20, 2015.
- Revision received February 28, 2015.
- Accepted March 3, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved