

Abstract
Background: The nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathway is critical in inflammation, proliferation and carcinogenesis. There exist three main players in this pathway. The inhibitor of NF-κB (IκB), IκB kinase (IκK)– NF-κB essential modulator (NEMO) complex and NF-κB. The IkK-NEMO complex activates NF-κB via phosphorylation of Iκβ and, eventually, leads to its proteasomal degradation. This leads to nuclear translocation of NF-κB and activation of target genes, such as cyclooxygenases and interleukins. The identification of anti-inflammatory compounds might be an effective strategy to target inflammatory disorders and cancer. Materials and Methods: In the present investigation, kaempferol was investigated in terms of its effect on NF-κB activity with a SEAP-driven reporter cell line, NF-κB DNA binding with electromobility shift assay (EMSA) and translocation of NF-κB-p65 from cytosol to the nucleus with western blot in Jurkat cells. Results: Kaempferol revealed anti-inflammatory activity, as shown in vitro and in silico. Molecular docking studies of kaempferol revealed comparable binding energies and similar docking poses on target proteins such as MG-132, a known NF-κB inhibitor. Conclusion: We conclude that kaempferol possesses anti-inflammatory activity.
- inflammation
- leukemia
- molecular docking
- NF-κB pathway
Inflammation is referred to as a protective response for eliminating injurious stimuli and initiating healing. Acute inflammation involves vasodilation, vascular leakage and leukocyte migration to injured sites, whereas chronic inflammation involves slow onset and prolonged activation, e.g. in hay fever, periodontitis, atherosclerosis and rheumatoid arthritis (1). Moreover, a strong association exists between chronic inflammation and cancer (2). Prolonged inflammation may lead to excessive production of reactive nitrogen species (RNS) and reactive oxygen species (ROS) contributing to the imbalance of pro- and antioxidants causing oxidative stress. This leads to damage of DNA, RNA, lipids and proteins. Consequent alterations, such as mutations, genomic instability, epigenetic changes and protein dysfunction, play roles in multi-stage carcinogenesis (3). Chronic inflammation causes tumorigenesis via DNA damage through RNS and ROS enhancing cell proliferation and stimulating angiogenesis (4).
Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) is a key player in inflammation, which drives the expression of pro-inflammatory and proliferative interleukins (ILs), tumor necrosis factor (TNF), interferons and cyclo-oxygenases upon response to carcinogens, growth factors and inflammatory stimuli. The NF-κB complex consists of the p50 and p65 subunits. Inactive NF-κB binds the inhibitory cytoplasmic protein IκBα. Binding of ligands to their receptors triggers the activation of IκB kinase (IκK) complex (IκK-α and IκK-β associated with NF-κB essential modulator (NEMO)). Subsequent phosphorylation and ubiquitinylation leads to proteasomal degradation of IκBα. NF-κB represents an important factor for cell proliferation and cancer progression (5, 6). Thus, targeting NF-κB is attractive to identify novel anti-inflammatory and anticancer compounds.
A target gene of NF-κB is COX-2 (cyclooxygenase 2), also known as prostaglandin-endoperoxide synthase (PTGS). It represents the rate-limiting enzyme during inflammatory prostaglandin formation. Arachidonic acids are metabolized by COX-2 to biologically-active eicosanoids, including prostaglandins. COX-2 contributes to carcinogenesis by inhibiting apoptosis (7), inducing angiogenesis (8) and by activation of chronic immune responses (9). NF-κB also promotes tumor growth in inflammation-associated cancers (10, 11).
NF-κB and IκB protein structures for molecular docking studies.
Herein, we tested the anti-inflammatory activity of kaempferol in Jurkat leukemia cells. Secreted embryonic alkaline phosphatase (SEAP)-driven NF-κB reporter cells and western blot experiments yielded supportive results for the anti-inflammatory effects. Using electromobility shift assay (EMSA), we corroborated that kaempferol indeed affects the DNA-binding activity of NF-κB. Molecular docking studies revealed that kaempferol shares comparable docking poses and binding energies with MG-132, a known NF-κB inhibitor (12, 13).
Materials and Methods
Chemicals and extracts. Kaempferol (purity ≥90%) was purchased from Enzo Life Sciences GmbH (Lörrach, Germany). MG-132 was obtained from Invitrogen (San Diego, CA, USA). Stock solutions (10μM) were prepared in dimethyl sulfoxide (DMSO) and stored at −20°C. Tumor necrosis factor-α (TNFα) was purchased from Sino Biological Inc (Beijing, China). Stock solutions (100 μg/ml) were prepared in sterile double distilled water and stored at −20°C. Quanti Blue was purchased from InvitroGen and prepared according to the manufacturer's recommendations.
Cell culture. Human Jurkat T leukemia cells were obtained from the Institute of Pharmaceutical Sciences (Albert-Ludwigs-University, Freiburg, Germany). Cells were maintained under standard conditions (37°C, 5% CO2) in RPMI medium (Gibco BRL, Eggenstein, Germany) supplemented with 10% fetal calf serum (FCS) and 1% penicillin/streptomycin (100 U/ml penicillin, 100 μg/ml streptomycin). Cells were passaged twice weekly. All experiments were performed with logarithmically growing cells.
NF-κB reporter assay. HEK293 cells stably expressing HEK-Blue-Null1 vector and SEAP on a NF-κB promoter were obtained from Invitrogen. The cells were cultured according to manufacturer's recommendations and treated with varying concentrations of kaempferol (5 μM, 20 μM and 40 μM) for 1 h, then induced with different concentrations of TNF-α (15, 30 and 100 ng/ml) for 24 h. DMSO served as a mock control. NF-κB activation was detected by measuring SEAP spectrophotometrically at 630 nm upon Quanti Blue addition (InvitroGen). MG-132 was used as a positive control compound to inhibit NF-κB (12, 13).
EMSA. Jurkat T cells (100,000 cells) were plated in 5 ml wells. After 24 h, cells were treated with 4 ng/ml TNF-α alone for 1 h and, then, treated for 72 h with 10 μM kaempferol. For control, cells were left untreated or were only treated for 73 h with 4 ng/ml TNF-α alone. Subsequently, cells were harvested by centrifugation and total NF-κB protein extracts were prepared. Extracts were incubated with labeled oligonucleotide (33P-labeled ATP) containing the NF-κB binding sequence and separated by electrophoresis. After gel drying, Fluorescent Image Analyzer (FLA-3000 Series, Fujifilm, Japan) was used to detect the labeled NF-KB oligonucleotide complex. Only the active NF-κB binds labeled oligonucleotides, not the inactive complex. Detailed conditions have already been described (14).
Western blotting. Jurkat T cells were treated by following the same approach; 1 h TNF-α induction (4 ng/ml) followed by 72 h treatment with 10 μM kaempferol. Control cells were left untreated or were only treated for 73 h with 4 ng/ml TNF-α alone. Cytoplasmic and nuclear protein extracts were prepared with NE-PER nuclear and cytoplasmic extraction reagent (Thermo Scientific, Rockford, IL, USA) supplemented with EDTA-Free Halt Protease Inhibitor Cocktail (Thermo Scientific) according to the manufacturer's protocol. Protein concentrations were determined by triplicate Nanodrop measurements with Nanodrop 1000 spectrophotomer (Thermo Scientific). Nuclear p65 levels were determined using rabbit anti-NF-κB p65 polyclonal antibody (1:1000; Thermo Scientific). Histone H3 protein levels, determined with a rabbit anti-Histone H3 polyclonal antibody (1:2000; Cell Signalling, Danvers, MA, USA), served as loading control. MG-132 (0.1 μM) was used as the positive control.
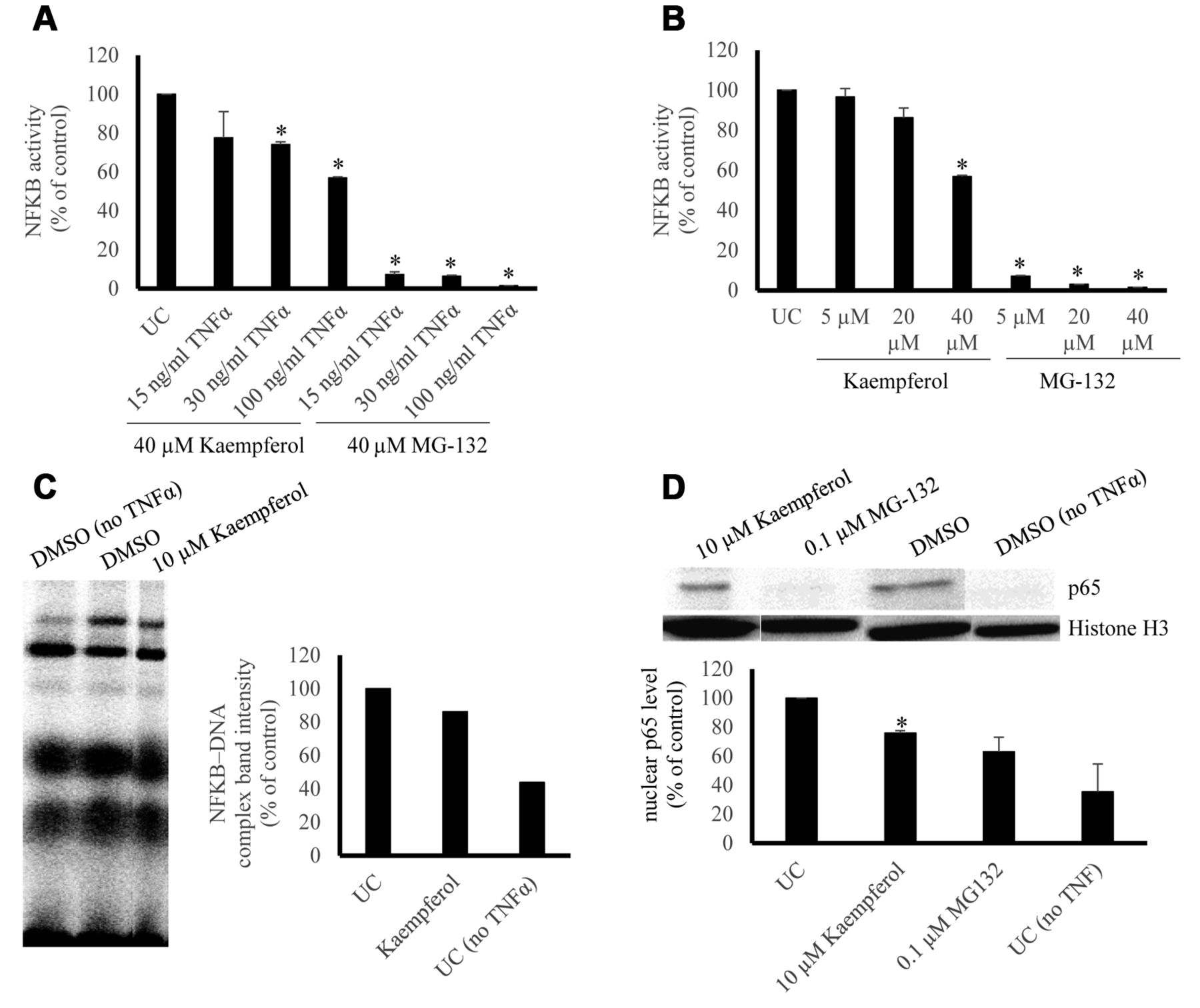
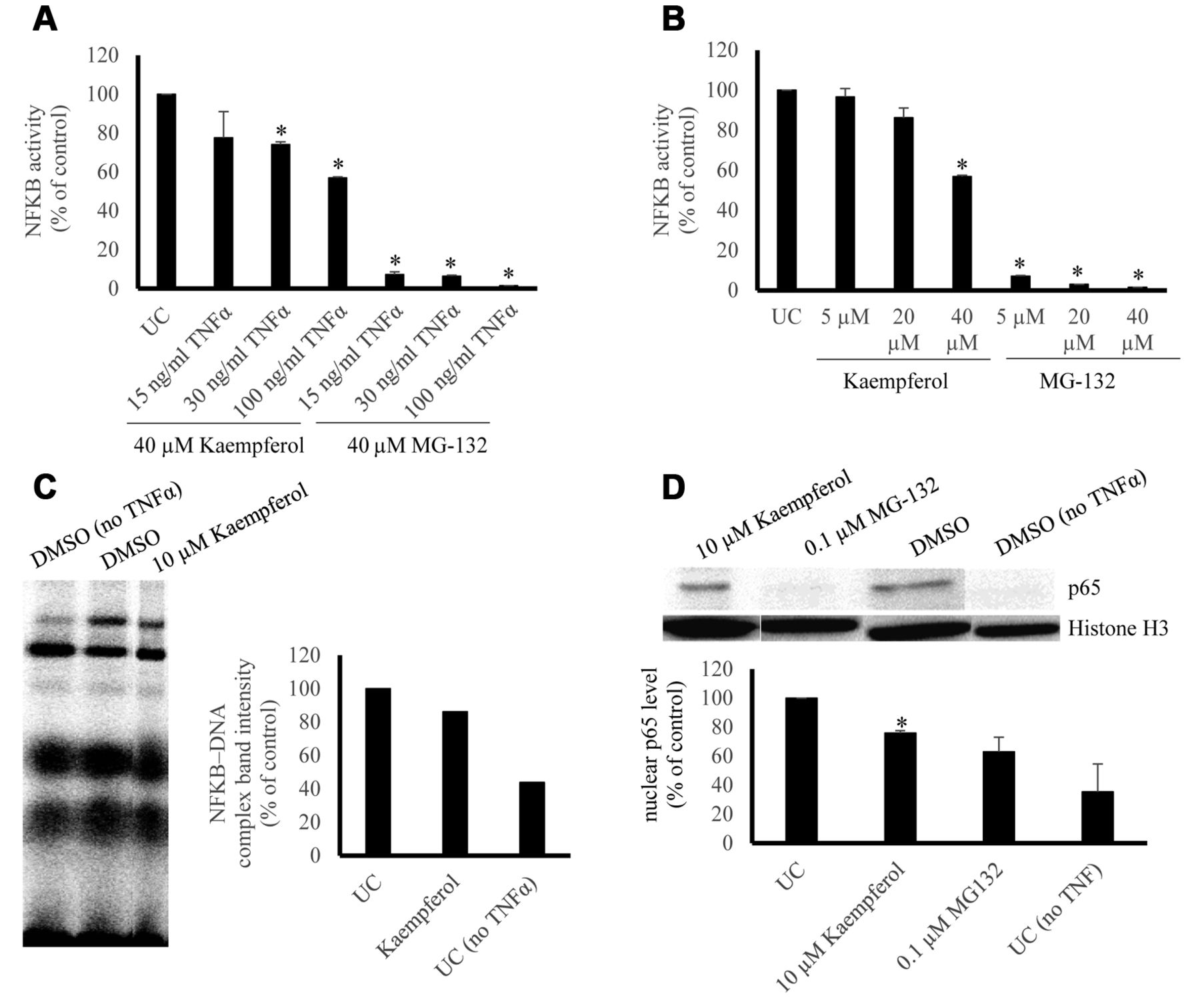
Effect of kaempferol on NF-κB activity, NF-κB–DNA binding and NF-κB p65 nuclear translocation. (A) Dose kinetics of TNFα and 40 μM kaempferol in comparison to 40 μM MG-132. Quantification was performed according to the untreated control and three repetitions were performed. (*p-value <0.05). (B) Dose kinetics of kaempferol and 100 ng/ml TNFα in comparison to MG-132. Quantification was performed according to the untreated control and three repetitions were performed. (*p-value < 0.05). (C) Effect of kaempferol on DNA binding activity of NF-κB. Kaempferol (10 μM) was tested in terms of its activity against NF-κB-DNA binding by EMSA. Quantification was performed by normalizing the NF-κB-DNA band intensities according to the untreated control. (D) Effect of kaempferol on nuclear NF-κB-p65 protein level. Normalized nuclear p65 protein levels were acquired according to the untreated control and three repetitions were performed. (*p-value < 0.05).
Molecular docking. Molecular docking was performed with the AutoDock 4 software (The Scripps Research Institute, CA, USA) on target NF-κB pathway proteins to evaluate the docking poses of kaempferol and MG-132. Visual molecular dynamics (VMD) and AutoDock Tools 1.5.7rc1 visualized docking poses. The selected proteins, their Protein Data Bank (PDB) IDs, target regions on the proteins and their relevant docking residues are represented in Table I. Grid parameters for the dockings are depicted in Table II. Defined molecular docking with 2,500,000 energy evaluations and 250 runs covering the regions of interest, as shown in Table I, were performed three times and the average of the lowest binding energies, mean binding energies and predicted inhibition constants were determined.
Statistics. The Student's t-test was performed with two tails and unequal variance. Experiments yielding p-values lower than 0.05 were considered statistically significant.
Results
NF-κB reporter assay. Kaempferol (40 μM) induced a dose-dependent inhibition of NF-κB activity in SEAP-driven NF-κB reporter cells with varying TNFα concentrations with weaker inhibition compared to MG-132 (Figure 1A). Comparable results were obtained by kaempferol with 100 ng/ml TNFα (Figure 1B).
EMSA. Kaempferol (10 μM) inhibited NF-κB–DNA interaction in the range of 15-20 % as shown by EMSA (Figure 1C).
Grid parameters for molecular docking studies.
Western blotting. TNF-α induction for 1 h followed by 72-h treatment with 10 μM kaempferol yielded inhibition of nuclear translocation of NF-κB p65 (24.2%±1.8), whereas 0.1 μM MG-132 yielded inhibition of (36.9%±10.1) (Figure 1D).
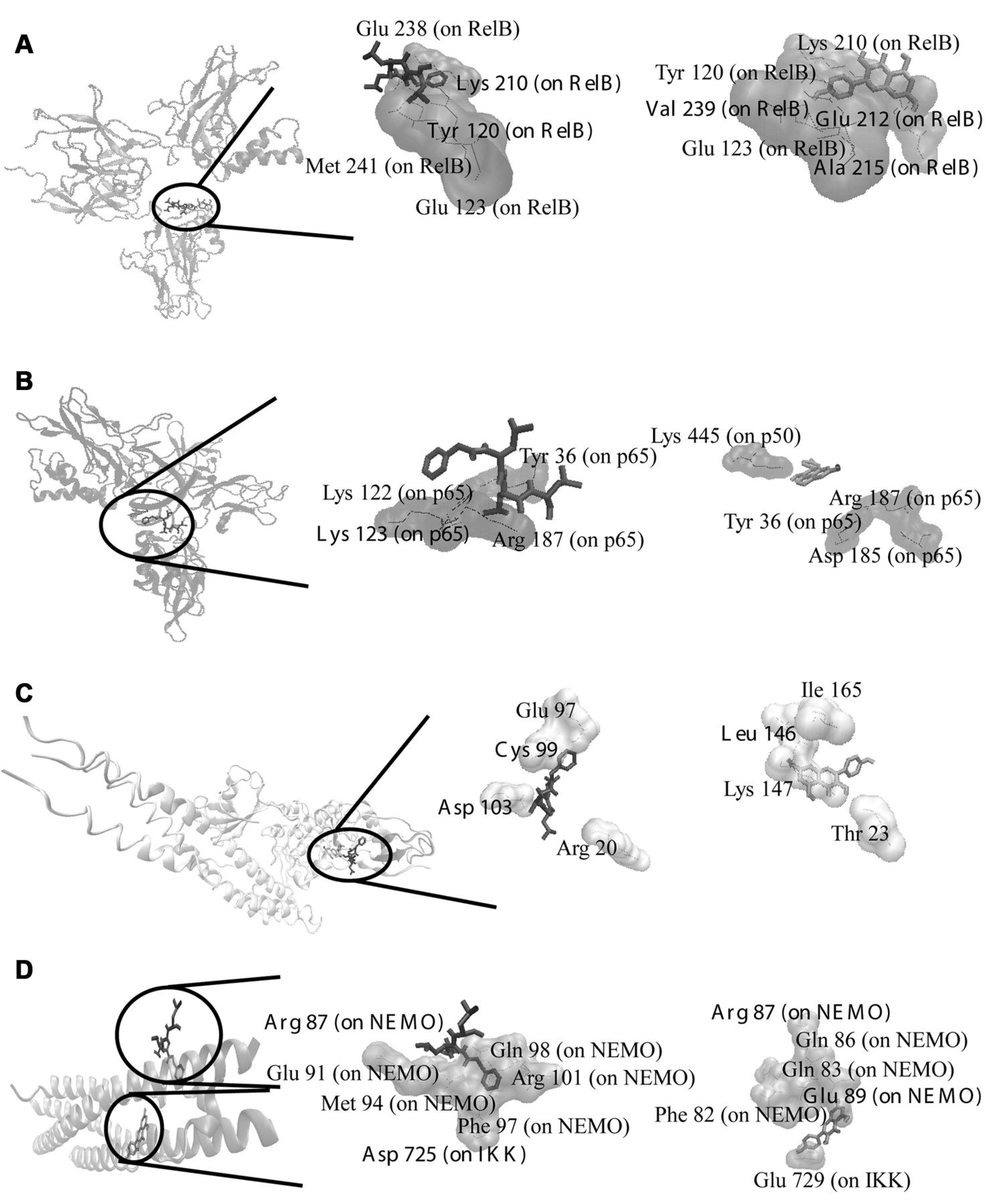
Molecular docking. In silico calculations were conducted on NF-κB pathway proteins. The molecular docking results are shown in Table III and Figure 2. The residues forming hydrogen bonds with kaempferol residing at the pharmacophore regions mentioned in Table I are labeled bold in Table III. Kaempferol showed similar docking poses and comparable binding energies as MG-132. The highest affinity was observed with a binding energy of −9.00±0.02 kcal/mol on NF-κB–DNA implying that kaempferol may intercalate into DNA and inhibit DNA binding of NF-κB.
Discussion
NF-κB is a transcription factor with a key role in inflammation since it is responsible for the expression of various pro-inflammatory and proliferative genes, such as interleukins, tumor necrosis factors, interferons and cyclooxygenases. Its over-expression contributes to proliferation, carcinogenesis and cancer progression (6, 15). Therefore, NF-κB may be a suitable target for drug development. For this purpose, we evaluated the anti-inflammatory effects of kaempferol by EMSA, SEAP-driven NF-κB reporter assay and Western blotting regarding NF-κB-DNA binding, NF-κB activity and NF-κB-p65 nuclear translocation. Kaempferol inhibited NF-κB-DNA binding and activity and decreased nuclear NF-κB p65 expression.
Molecular docking of kaempferol to proteins of the NF-κB pathway.
{kind=link}
{kind=link}
Molecular docking studies of kaempferol on NF-κB pathway proteins. Known inhibitor MG-132 (black bond representation), kaempferol (light gray bond representation). (A) Docking poses into the DNA binding site of NF-κB (PDB code: 3DO7 in cartoon representation). (B) Docking poses into the DNA binding site of NF-κB-DNA complex (PDB code: 1VKX in cartoon representation). (C) Docking poses into the ATP binding site of IκK (PDB code: 3RZF in cartoon representation). (D) Docking poses into the interaction site of IκK-NEMO complex (PDB code: 3BRT in cartoon representation). The residues involved in hydrogen bond interaction are mentioned in Table III.
Kaempferol may act in an anti-inflammatory manner by binding to NF-κB, IκK-NEMO association domain and IκK. The fact that it bound with even higher affinities to DNA-bound NF-κB than free NF-κB potentiates the DNA binding inhibition of kaempferol. Indeed, this was supported by EMSA assays. Thus, different sites of the NF-κB pathway may be targeted by kaempferol. Previous studies have shown that kaempferol is an anti-inflammatory compound (16) with activity against cancer in vivo (17). Flavonoids possess anti-inflammatory activity (18) and anticancer activity as shown by an in vivo study (19).
Taken together our results show that kaempferol may act as anticancer compound by affecting the NF-κB pathway. Further analyses in pre-clinical and clinical studies are warranted to clarify the therapeutic potential of kaempferol for clinical use.
- Received January 29, 2015.
- Revision received February 10, 2015.
- Accepted February 12, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved