

Abstract
N-glycosylation of integrins plays an important role in cancer progression. Increased αvβ3 integrin expression during melanoma progression is well-documented but the role of its glycans in tumorigenesis is still poorly understood. In the present study we used the WM793 primary melanoma cell line and its highly metastatic variant, WM1205Lu, to examine αvβ3 glycosylation. Lectin precipitation, enzyme digestion and the use of swainsonine (SW) showed that αvβ3 integrin glycosylation differs significantly between primary and metastatic melanoma cells. High-mannose structures and complex glycans with bisecting N-acetylglucosamine (GlcNAc) were more abundant in both subunits of primary cells. We also observed a shift in the sialylation of αvβ3 integrin related to reduction of α2-6-linked sialic acid expression and an increase of α2-3 sialylation of both subunits in melanoma progression. Metastatic melanoma migration on vitronectin (VN) was reduced in the presence of antibody against αvβ3 and the lectins phytohemagglutinin-L (PHA-L), Sambucus nigra agglutinin (SNA) and Maackia amurensis (MAA) in woundhealing assays. Our results show that the acquisition of metastatic competence by melanoma cells is accompanied by alteration of αvβ3 integrin glycosylation and that both αvβ3 and β1-6-branched sialylated complex-type N-glycans promote metastatic melanoma migration on VN.
- αvβ3 integrin
- glycosylation
- melanoma
- migration
- vitronectin
Aberrant glycosylation of integrins is closely associated with the metastatic competence of different cancers (1, 2), including melanoma (3, 4). Integrin αvβ3 is highly N-glycosylated during post-translational processing; the αv subunit possesses 13 potential sites of asparagine-linked (N-linked) glycosylation and the β3 subunit possesses 6 of them (5). Integrin αvβ3 is an adhesion protein important in tumor angiogenesis, metastasis and inflammation. αvβ3 is the main vitronectin (VN) receptor but it also binds other extracellular proteins, fibronectin, fibrinogen, laminin, von Willebrand factor and collagens (3, 6). It probably is the most prominent surface receptor up-regulated in different tumor types and it is, thus, considered to be one of the most important prognostic biomarkers of cancer (7). Humanized monoclonal antibodies against αvβ3 have been tested on mice and also in clinical trials as anticancer drugs to inhibit tumor growth, angiogenesis and metastasis (8).
Melanoma is one of the most common and most aggressive human cancers. Acquisition of metastatic competence by melanoma cells growing in dermis as the vertical growth phase (VGP) is generally an irreversible step of melanoma progression. Most therapies are ineffective when melanoma is diagnosed at that stage. The formation of metastatic lesions in distinct organs is the leading cause of death among melanoma patients. The search for effective ways to inhibit propagation of melanoma demands a comprehensive examination of the changes occurring during the transition from VGP to metastatic lesions. Glycosylation is an essential process in the contact of cells with extracellular matrix proteins (ECM). Changes in the N-oligosaccharide composition of integrins can modify the adhesive interactions of tumor cells with the surrounding environment, thus facilitating their spread. Increased αvβ3 integrin expression during melanoma progression is well documented (9-11) but the role of its glycans in tumorigenesis is still poorly understood. In the present study we analyzed αvβ3 integrin glycosylation in the last two stages of melanoma progression. For this, we chose two genetically-related human melanoma cell lines: WM793 primary cells derived from early VGP and WM1205Lu cells from lung metastasis. Our results show that the acquisition of metastatic competence by melanoma cells is accompanied by alteration of αvβ3 integrin N-glycosylation. High-mannose oligosaccharides and sialylated complex-type N-glycans with bisected N-acetylglucosamine (GlcNAc) undergo change in WM1205Lu metastatic cells. We also report that both αvβ3 integrin and sialylated complex-type oligosaccharides promote only metastatic and not primary melanoma migration on VN.
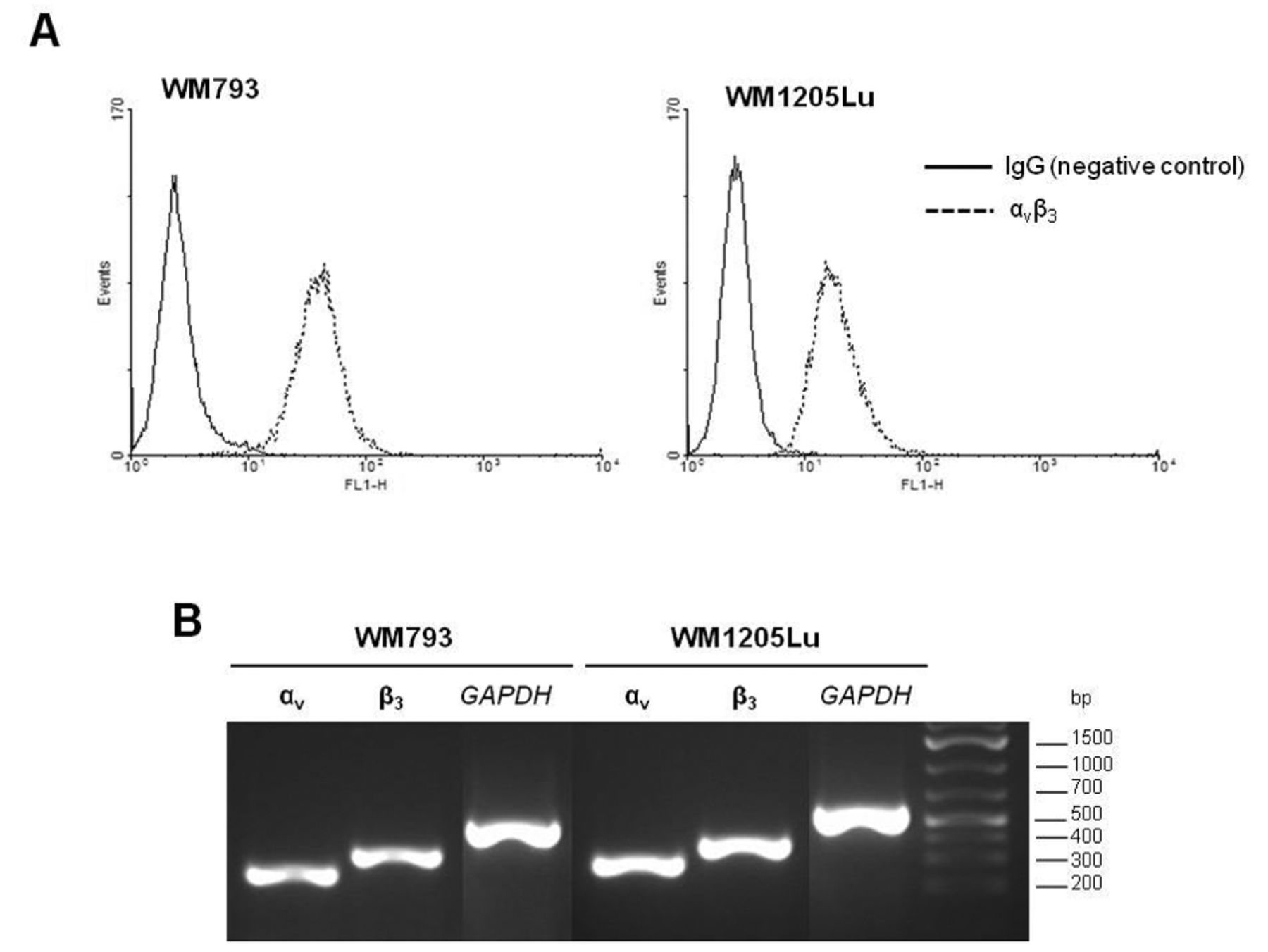
Primary WM793 and metastatic WM1205Lu melanoma cells show similarly high expression of αvβ3 integrin on protein (A) and gene (B) levels. (A) Cells were trypsinized and incubated with anti-αvβ3 primary monoclonal antibody (MAB1976), followed by incubation with the secondary antibody conjugated with FITC. Integrin αvβ3 expression was determined by flow cytometry, removing nonspecific fluorescence relative to the IgG control. (B) Total RNA (1 μg) was reverse-transcribed and cDNA was amplified by RT-PCR. The PCR products or αv, β3 and GAPDH (endogenous control) were resolved on 2% agarose gel containing ethidium bromide.
Materials and Methods
Cell lines and reagents. VGP WM793 and highly metastatic WM1205Lu human melanoma cell lines were obtained through participation in the ESTDAB project (the European Searchable Tumour Cell-Line Database; IPD-ESTDAB: http://www.ebi.ac.uk/ipd/estdab/).
Polyclonal primary antibodies used in western blotting against αv (AB1930) and β3 (AB1932) integrin subunits were obtained from Millipore (Chemicon, Temecula, CA, USA). Goat anti-rabbit horse-radish peroxidase (HRP)-conjugated IgG (7074S) was purchased from Cell Signaling Technology (Danvers, MA, USA). Alkaline phosphatase-conjugated sheep anti-rabbit IgG (AP322A) for Western blotting and mouse anti-αvβ3 (MAB1976, clone LM609) used in flow cytometry, wound-healing assays and confocal microscopy were obtained from Millipore (Chemicon). FITC-conjugated rabbit anti-mouse IgG, F(ab’)2 (F0313) and mouse IgG1 (X0931), used as a negative control in flow cytometry, were obtained from Dako (Glostrup, Denmark) and Cy3-conjugated goat anti-mouse (115-166-003) from Jackson ImmunoResearch Laboratories (West Grove, PA, USA). Biotinylated lectins (GNA, B-1245; PHA-L, B-1115; PHA-E, B-1125; DSA, B-1185; LEL, B-1175; SNA, B-1305; MAA, B-1265; AAA, B-1395) and agarose-bound streptavidin (SA-5010) were obtained from Vector Labs (Burlingame, CA, USA). Vitronectin (CC080) was purchased from Millipore (Chemicon). Swainsonine (SW) isolated from Metarrhizium anisopliae (S8195) was purchased from Sigma-Aldrich (St. Louis, MO, USA). N-glycosidase F (PNGase F) isolated from Flavobacterium meningosepticum (11 365 181 001) and endo-β-N-acetylgluco saminidase H (Endo H) from Streptomyces plicatus (11 088 726 001) were obtained from Roche (Mannheim, Germany), Streptococcus pneumoniae sialidase (N7271) from Sigma-Aldrich and Arthrobacter ureafaciens sialidase (GK80040) from Glyko (Prozyme, Hayward, CA, USA).

Glycosylation of αvβ3 integrin differs between primary and metastatic melanoma cells in the amount of (A) total N-glycans, (B) high-mannose and hybrid structures and (C) sialic acids but not (D) β1-6-branched structures. (A) N-glycans were released from glycoproteins using PNGase F from Flavobacterium meningosepticum, (B) high-mannose and hybrid type glycans using Endo H from Streptomyces plicatus, (C) α2-3-linked sialic acids using Streptococcus pneumoniae sialidase and α2-3,6,8-linked sialic acids using Arthrobacter ureafaciens sialidase; (D) synthesis of complex-type glycans, including β1-6-branched glycans, was blocked using swainsonine (SW) from Metarrhizium anisopliae. The amount of glycans in αv and β3 integrin subunits was calculated based on loss of molecular mass in SDS-PAGE. Values are means±SD of three independent experiments, asterisked values differ significantly between primary and metastatic cells at p<0.05 by Duncan's test.
Cell culture conditions and cell lysate preparation. The cells were grown in RPMI1640 medium (Invitrogen, Paisley, UK) supplemented with 10% FBS (Gibco, Paisley, UK), 100 U/ml penicillin and 100 μg/ml streptomycin (Sigma-Aldrich) at 37°C in a CO2 incubator (Lab Line Instruments, Melrose Park, IL, USA). Cells were harvested upon 70-80% confluency using 0.25% trypsin solution containing 0.02% EDTA (T3924; Sigma-Aldrich). For some experiments the cells were cultured in the presence of swainsonine (10 μg per 1 ml medium) for 24 h.

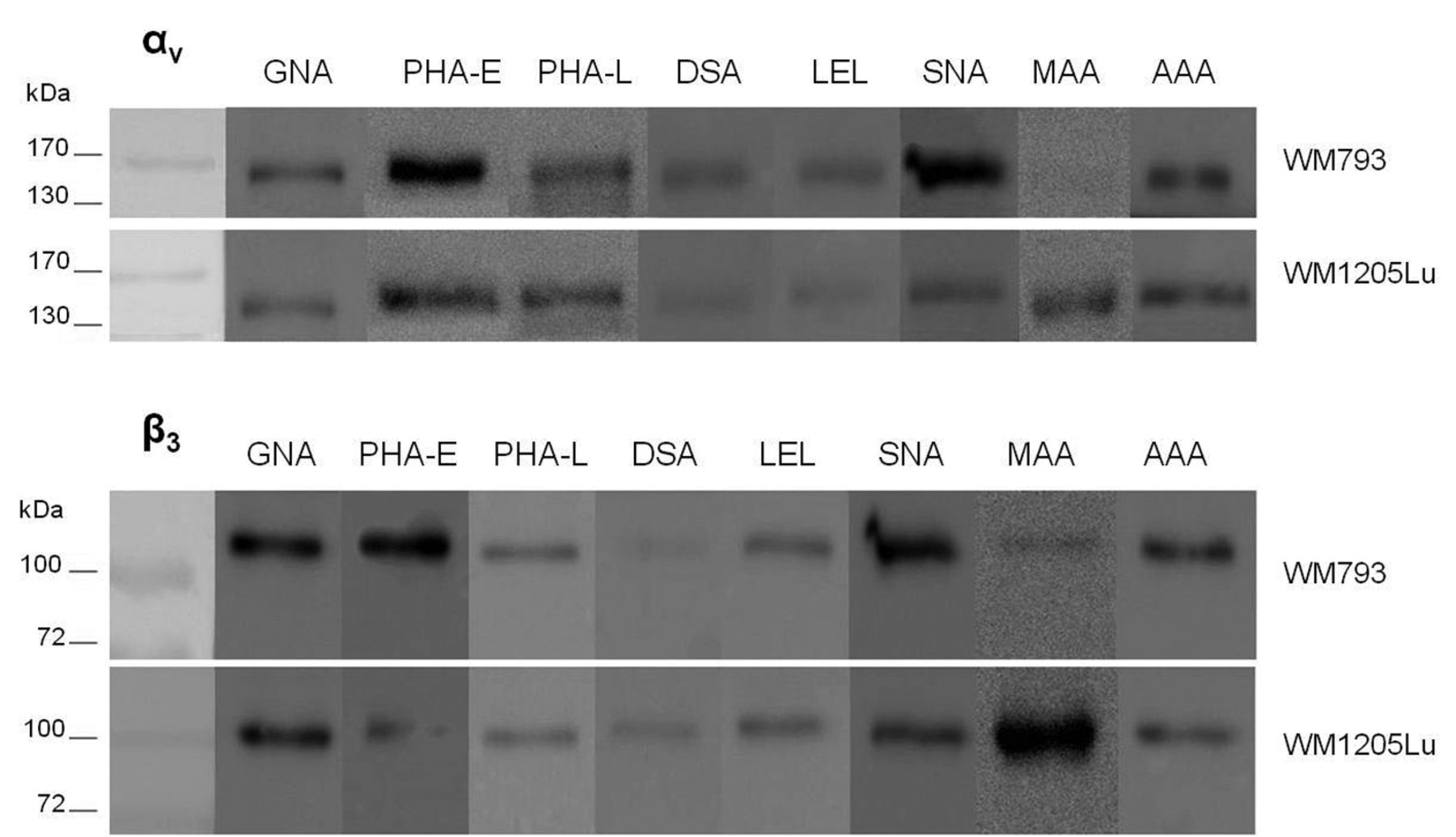
The content of high-mannose glycans and sialylated complex-type structures with bisecting GlcNAc in αvβ3 integrin changes during acquisition of the metastatic phenotype. Cell lysate glycoproteins (100 μg) were incubated with biotinylated lectins GNA, PHA-L, PHA-E, DSA, LEL, SNA, MAA and AAA, follow by precipitation using agarose-bound streptavidin. Glycoproteins released by boiling in Laemmli sample buffer were destined for αv and β3 integrin subunit immunodetection.
The cells were lysed in RIPA buffer (89900; Thermo Scientific, Rockford, IL, USA) containing protease inhibitor cocktail (P2714; Sigma-Aldrich). The mixture was centrifuged (15,000 rpm) for 30 min at 4°C. The protein concentration was quantified in the collected supernatant using the Total Protein Kit, Micro Lowry, Peterson's Modification (TP0300; Sigma-Aldrich) to enable comparison of equal total protein amounts between samples of the two cell lines.
Reverse transcription-polymerase chain reaction (RT-PCR) for αvβ3 integrin expression. Semiquantitative RT-PCR was used to assess αv (ITGAV) and β3 (ITGB3) gene expression. RNA was extracted from WM793 and WM1205Lu cells using the RNeasy Protect Mini Kit (Qiagen, Hilden, Germany) and reverse transcription was carried out using the Omniscript RT kit (Qiagen) according to the manufacturer's instructions. PCR was performed using Taq PCR Master Mix (Qiagen) in a DNA Engine PTC-200 thermocycler (MJ Research, Watertown, MA, USA). Each reaction consisted of 35 cycles of denaturation (1 min at 94°C), primer hybridization (1 min at 59°C for both αv and β3 and 59.6°C for GAPDH) and elongation (2 min at 72°C). The reaction began from an initial denaturation (3 min at 94°C) and the final step was elongation (10 min at 72°C). The expression of αv and β3 integrin subunits was normalized to the expression level of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) used as housekeeping gene. The primer sequences were as follows: GACTGTGTGGAAGACAATGTCTGTAAACCC (αv forward), CCAGCTAAGAGTTGAGTTCCAGCC (αv reverse), CTGGTGTTTACCACTGATGCCAAG (β3 forward), TGTTGAGGCAGGTGGCATTGAAGG (β3 reverse), 5’-CCACCCATGGCAAATTCCATGGCA-3’ (GAPDH forward), and 5’-TCTAGACGGCAGGTCAGGTCCACC-3’ (GAPDH reverse).
The PCR amplification products were resolved on 2% agarose gel containing ethidium bromide and were visualized in a transilluminator (UVItec, Cambridge, UK). The amplification product lengths were 305 bp for αv, 393 bp for β3 and 599 bp for GAPDH.
Flow cytometry. The cells were harvested by trypsinization and washed in cold PBS. Then the cells were incubated 45 min with mouse antibody against the αvβ3 complex (1:100), followed by incubation with polyclonal rabbit anti-mouse antibody conjugated with FITC (1:10) for 45 min. After both incubations, the cells were centrifuged (1,100 rpm, 5 min, 4°C) and washed with cold PBS. Stained cells were analyzed with a FACSCalibur flow cytometer (BD Biosciences, San Diego, CA, USA) using the CellQuest software (BD Biosciences, San Diego, CA, USA). The expression level of αvβ3 integrin was determined relative to nonspecific binding of mouse IgG1 (negative control).
Enzymatic digestion of glycans. The N-oligosaccharides were released from the glycoproteins with peptide-N-glycosidase F from Flavobacterium meningosepticum (11 365 181 001; Roche). The cell extract proteins were denatured 3 min in 50 mM sodium phosphate buffer, pH 7.5, containing 5% SDS. After cooling to RT, SDS was neutralized with 10% nonidet P-40 (NP-40). Glycans were digested overnight at 37°C using 0.5 mU PNGase F per 15 μg glycoproteins.
The content of high-mannose and hybrid glycans in integrin subunits was determined using Endo H from Streptomyces plicatus. The cell glycoproteins were suspended in sodium phosphate buffer, pH 5.5, to obtain the 50 mM final buffer concentration. Protein denaturation was carried out in the presence of 2% SDS for 3 min. After cooling to RT, 25 mU Endo H per 15 μg glycoproteins was added and the samples were incubated overnight at 37°C.
The cell extract proteins (15 μg) were treated with Streptococcus pneumoniae α2-3 sialidase and Arthrobacter ureafaciens sialidase, which specifically removes α2-3,6,8-linked sialic acids. Glycoproteins were digested in the incubation buffers (50 mM sodium phosphate buffer, pH 6.0, for α2-3 sialidase; 50 mM sodium acetate, pH 5.0, for α2-3,6,8 sialidase) at 37°C overnight.
In each of the above cases, the reaction was stopped by heating at 100°C for 10 min after overnight incubation at 37°C. Both digested and nondigested samples were boiled in Laemmli sample buffer (LSB, 161-0737; Bio-Rad, Hercules, CA, USA) prior to SDS-PAGE on 10% polyacrylamide gels. Glycan amounts were calculated based on loss of molecular mass in UVImap Image Quantification software (UVItec).
Lectin precipitation. Cell lysate proteins (100 μg) were incubated with 4 μl biotinylated lectins (GNA, PHA-L, PHA-E, DSA, LEL, SNA, MAA, AAA) in HEPES buffer (10 mM HEPES, pH 7.5, containing 150 mM NaCl, 0.1 mM CaCl2 and 0.01 mM MnCl2). After overnight incubation at 4°C, the glycoprotein-lectin complexes were precipitated using 20 μl agarose-bound streptavidin for 3 h at 4°C. After centrifugation (14,500 rpm, 5 min, RT), the supernatants from lectin-treated lysates were removed. The precipitates were washed three times in HEPES buffer and once in PBS and glycoproteins were released from the glycoprotein-lectin-agarose complexes by boiling in LSB at 100°C for 10 min. After final centrifugation, the supernatants were collected and destined for integrin subunit immunodetection.
Western blotting. Glycosylation of integrin subunits was analyzed by Western blotting. Equal amounts of proteins (15 μg) in LSB (for the β3 subunit, 5% β-mercaptoethanol was added) were separated on 10% SDS-PAGE gel, transferred to a PVDF membrane and detected using the appropriate antibodies. After blocking in 1% BSA in TBST, the membranes were incubated for 1 h at RT with polyclonal rabbit anti-αv and anti-β3 antibodies diluted 1:1000 in 1% BSA in TBST. Then, goat anti-rabbit IgG was applied in a 1:4000 dilution, followed by incubation at RT for 1 h. The secondary anti-rabbit antibodies were conjugated with alkaline phosphatase (AP) or horseradish peroxidase (HRP) and the integrin subunits were visualized by colorimetric reaction and chemilumi nescence, respectively. Colorimetric visualization was done by incubating the membranes with BCIP and NBT substrates for AP (Roche) and chemiluminescence using Western blotting luminol chemiluminescence reagent (Santa Cruz Biotech.) in the Gnome System (Syngen, Cambridge, UK).
Cell adhesion assay. Cell adhesion abilities were tested as described previously by Janik et al. (12) with minor modifications. A 96-well plate was coated with vitronectin (VN, 0.1μg/ml PBS) and left overnight at 4°C. Control cells and cells treated with swainsonine (10 μg/ml) 24 h before the assay were starved in serum-free RPMI 30 min before collecting by trypsinization. The cells (5×104) were seeded into wells coated with VN and allowed to adhere for 1 h at 37°C. After removal of nonadhering cells, adhering cells were stained with calcein for 20 min. Fluorescence was measured at 495/520 nm with a microtiter plate reader (Infinite F200 Pro; Tecan, Grödig, Austria). Nonspecific binding of cells to BSA-coated wells was subtracted.
Confocal microscopy. Cells were seeded on VN-coated (0.1 μg/ml PBS) or uncoated coverslips overnight at 4°C. Before staining, the cells were washed three times with PBS, fixed in 2% PFA for 20 min at RT and incubated with monoclonal antibodies against αvβ3 integrin (1:10 in 2% BSA in PBS) for 45 min, followed by Cy3-conjugated goat anti-mouse antibody (1:100 in 2% BSA in PBS) for 45 min at RT. The slides were mounted with Vectashield media with DAPI, the coverslips were sealed and were kept at 4°C. Cell fluorescence was visualized using a Zeiss confocal microscope (Jena, Germany) and analyzed in an LSM Image Browser (Zeiss, Jena, Germany).
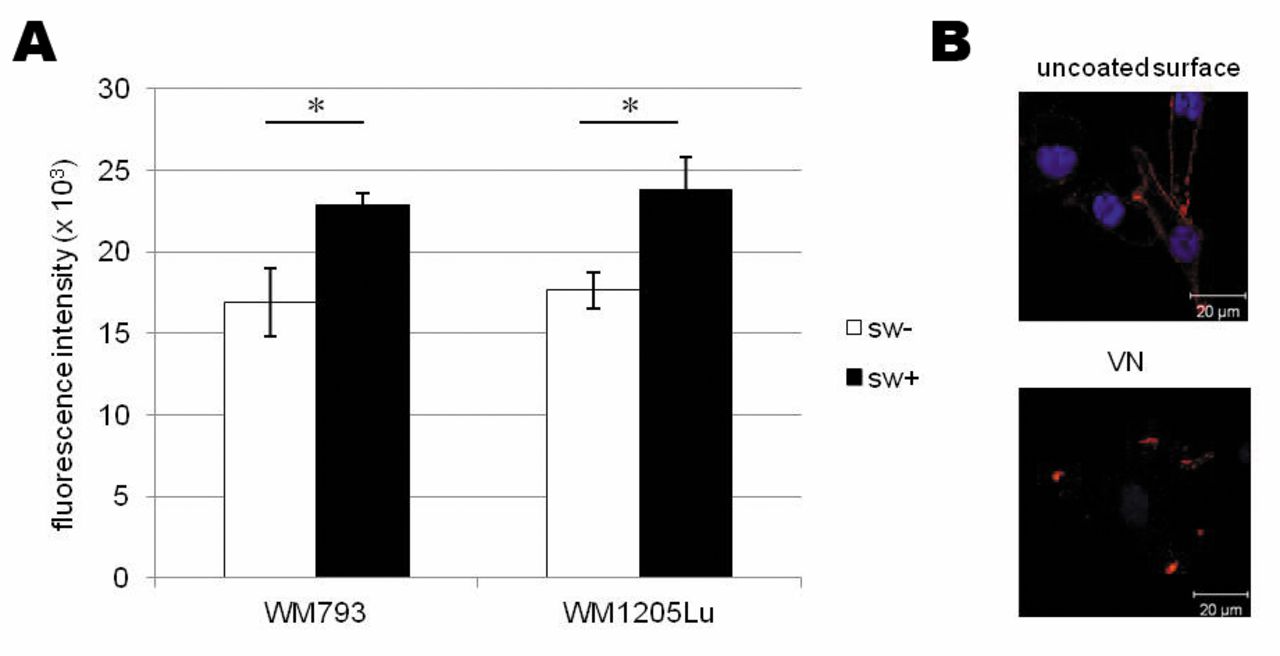
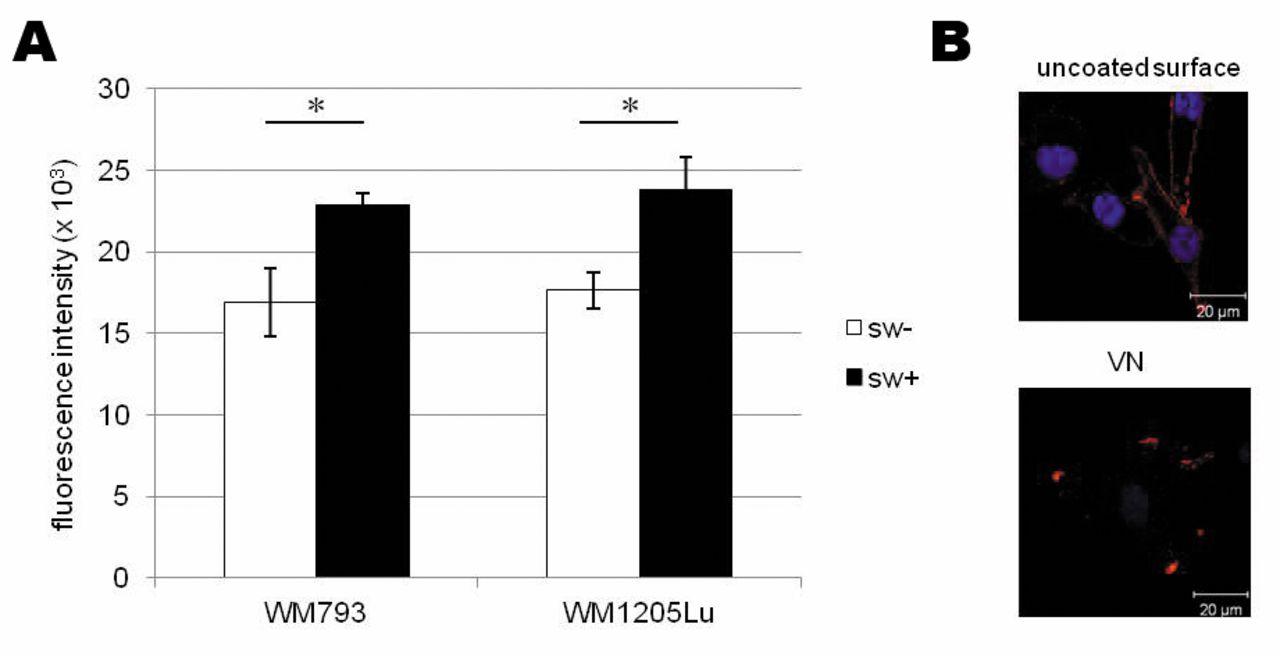
Melanoma cells bind strongly to vitronectin (VN). (A) Both WM793 primary and WM1205Lu metastatic cell adhesion to VN is affected by β1-6-branched N-glycans as determined with the use of swainsonine (SW). Values are means±SD of three repetitions, and differ significantly between treated and untreated cells at p<0.05 by Duncan's test. (B) Integrin αvβ3 clusters in focal adhesions of melanoma cells adhering to VN, unlike dispersed expression on the whole membrane of cells growing on an uncoated surface.
Wound-healing assay. Cells were seeded onto a 6-well plate precoated overnight at 4°C with VN. Three scratches were made in the cell monolayer of each well with a yellow pipette tip. Migrating cells were incubated 24 h in the presence of antibodies against αvβ3, lectins PHA-L, PHA-E, SNA and MAA, as well as 1-deoxymannojirimycin (dMM), an inhibitor of N-glycan synthesis that induces accumulation of oligomannose-type chains. The wound sites were photographed in 10 random fields using the Zeiss AxioVision Rel.4.8 image analysis software. The average extent of wound closure was determined by measuring the width of the wound at the starting point and after 24 h.
Statistical analyses. The results are given as means of three measurements±standard deviation (SD). The Duncan's test was used to compare mean values. p-Values<0.05 were considered significant.
Results
Expression of αvβ3 integrin on gene and protein levels. In this study we evaluated αvβ3 integrin expression on gene and protein levels. The expression of αv (ITGAV) and β3 (ITGB3) genes was normalized to GAPDH housekeeping gene expression in semiquantitative RT-PCR. We did not observe differences in αv and β3 subunit expression between primary WM793 and metastatic WM1205Lu cells (Figure 1B). Monoclonal antibody against the αvβ3 complex was used to detect its cell surface expression. Flow cytometry analysis showed comparatively high expression of integrin in both melanoma cell lines (Figure 1A). Nearly all WM793 and WM1205Lu cells expressed αvβ3 (98.74% and 99.35% of cells, respectively).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
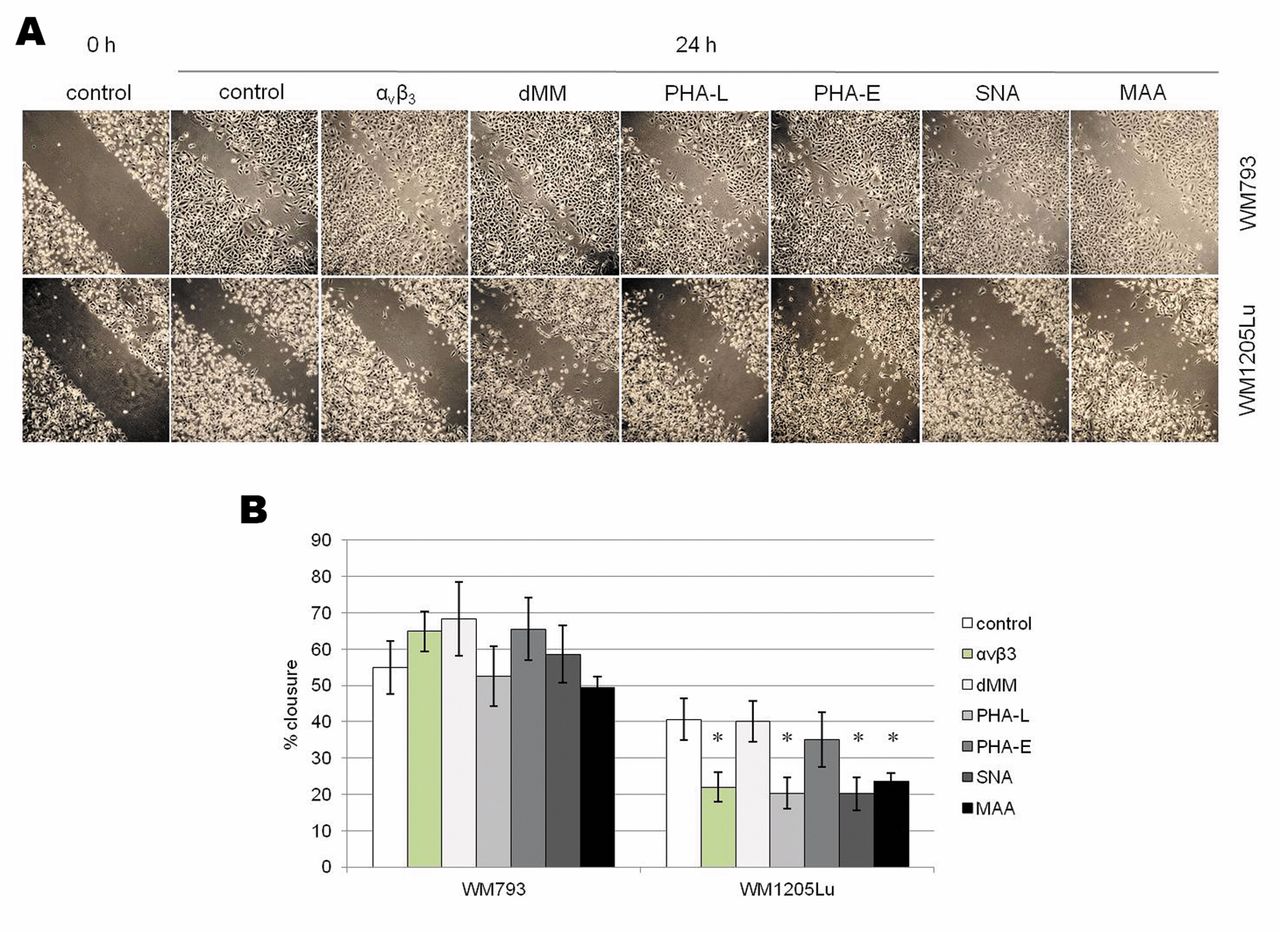
Integrin αvβ3 and sialylated β1-6-branched complex-type N-glycans are involved in metastatic melanoma migration on VN. Wound-healing migration assays were performed with melanoma cells growing on VN-coated plates and treated with anti-αvβ3 monoclonal antibody, lectins PHA-E, PHA-L, SNA and MAA, as well as dMM inhibitor. Cells were scratched with pipette tips. Each wound was photographed (A) and the rate of migration into the wound area was assessed at the starting point and after 24 h (B). Values are means±SD of three wounds, asterisked values differ significantly between treated and untreated cells at p<0.05 by Duncan's test.
Glycosylation of αvβ3 integrin. Content of N-glycans. N-linked oligosaccharides in each subunit were released by PNGase F cleaving and their content was quantified by calculation of loss of molecular weight after enzyme treatment versus that of undigested glycoproteins. We determined that both integrin subunits are heavily N-glycosylated. N-glycan amounts were higher in both αvβ3 subunits from WM793 primary melanoma cells (Figure 2A). In primary cells approximately 30% of αv molecular weight was from N-glycans; the corresponding figure for metastatic cells was 28%. The pool of N-glycans in β3 subunits from primary cells accounted for 16% of all glycoprotein molecules; for metastatic cells the figure was 12%.
High-mannose and hybrid-type N-glycans. We applied two different approaches to estimate oligomannose and hybrid-type N-glycans. Galanthus nivalis lectin (GNA) was used for precipitation of these N-glycan types and endo-β-N-actylglucosaminidase H (Endo H) enzyme for cleaving them. Endo H digestion showed that the αv and β3 subunits from WM793 primary cells are richer in these structures than those from metastatic cells (Figure 2B). GNA precipitation confirmed the higher content of high-mannose and hybrid glycans in both subunits from WM793 primary cells (Figure 3).
Sialylation of αvβ3 integrin. In most sialoglycoconjugates, sialic acid residues are attached terminally to complex-type N-linked oligosaccharides by α2-6 or α2-3 linkages. To study the sialylation of melanoma cells we used linkage-specific lectins from Sambucus nigra (SNA) and Maakia amurensis (MAA), which bind sialic acid linked by α2-6 and by α2-3, respectively. We also estimated melanoma sialylation using bacterial sialidases that catalyze the hydrolysis of terminal sialic acids linked by α2-3 and α2-3,6,8. Lectin precipitation and neuraminidase digestion showed differences in the expression of sialic acids between primary and metastatic melanoma cells. Treatment of cell lysate proteins with α2-3 sialidase prior to SDS-PAGE caused different mobility shifts of the αv subunit in the melanoma cell lines due to differences in their amounts of sialic acids (Figure 2C). Subunit αv expressed in WM1205Lu metastatic cells possessed more α2-3-linked sialic acids than in WM793 primary melanoma cells but total sialylation was signifi cantly higher in the αv integrin subunit in WM793 primary cells than in the WM1205Lu metastatic cell line. It was not possible to determine α2-3-linked sialic acid in the β3 subunit by this method because the shift in this subunit's mobility was too small.
Immunodetection of both subunits in MAA precipitates showed that α2-3-linked sialic acids recognized by MAA were much more abundant in αvβ3 integrin from metastatic than from primary cells. Sialic acids occupying terminal α2-6 linkages detected by SNA lectin were observed mainly in αvβ3 integrin from WM793 primary cells (Figure 3). The results of MAA lectin precipitation correspond to S. pneumoniae sialidase-induced α2-3 desialylation of the αv subunit. Both methods showed higher content of α2-3-bound sialic acids in WM1205Lu metastatic cells than in WM793 primary cells. We could not determinate sialic acid content in the β3 subunit by neuraminidase digestion because the differences in electrophoretic mobility between digested and nondigested proteins were too small.
Bisecting GlcNAc and β1-6-branched N-glycans. We analyzed the expression of the bisecting GlcNAc (N-acetylglucosamine) residue β1-4-linked to β Man of the N-glycan core in αvβ3 integrin by precipitation with Phaseolus vulgaris erythroagglutinin (PHA-E). Both subunits of primary melanoma were precipitated in higher amounts than in the metastatic line (Figure 3).
The Golgi α-mannosidase II inhibitor swainsonine (SW) was used to assess the amount of complex-type glycans, among them β1-6-branched GlcNAc in αvβ3 integrin subunits. Immunodetection of both subunits did not show differences in motility shifts as calculated by subtracting the molecular mass of SW-treated and untreated proteins (Figure 2D). This result was verified by precipitation with phytohemagglutinin-L (PHA-L) lectin, which specifically recognizes β1-6-branched complex-type oligosaccharides. The values for PHA-L precipitates in both subunits confirm the results obtained with the use of SW (Figure 3). The intensity of the bands for the αv and β3 subunits did not differ in intensity between primary and metastatic cells.
Fucosylation and polylactosamine structures. Precipitation with Aleuria aurantia lectin (AAA), specific for α1-6 Fuc, did not show differences in the content of this sugar residue in αvβ3 integrin between the two melanoma cell lines. Binding of integrin subunits by AAA was similar in WM793 and WM1205Lu cells (Figure 3).
To detect glycans with lactosamine repeats GlcNAcβ(1-4)Gal in integrin subunits, for precipitation we used two lectins with different specificity, Datura stramonium lectin (DSA) and Lycopersicon esculentum lectin (LEL), followed by immunodetection to estimate the amount of 2-3 and ≥3 lactosamine units, respectively. The two cell lines showed similar intensity of the bands corresponding to precipitated αv and β3 subunits (Figure 3).
Melanoma cell adhesion to VN. In functional assays we used VN, the main ECM ligand for the αvβ3 receptor (13); their interaction is implicated in melanoma migration (12). To determine whether β1-6-branched complex-type N-glycans participate in melanoma cell adhesion to VN, we performed adhesion assays for cells treated with SW. Swainsonine's ability to block synthesis of complex-type N-glycans, among them β1-6-linked structures, facilitates the study of their function. In the two melanoma cell lines, we observed similar increases of adhesion to VN after SW treatment (Figure 4A). We conclude that β1-6-linked complex-type glycans affect melanoma cell adhesion to VN.
Confocal microscopy of immunostaining showed that αvβ3 is distributed over the whole surface of melanoma cells cultured on an uncoated surface; in the presence of VN, αvβ3 was clustered in focal adhesion structures (Figure 4B). Figure 4B shows only the results for WM1205Lu cells; the same effect was observed in both melanoma cell lines.
Role of αvβ3 integrin and glycosylation in melanoma migration on VN. We used monoclonal antibodies against αvβ3 integrin, unlabeled lectins and glycosylation inhibitor in assays of melanoma wound healing on VN to examine the effects of integrins and the effects of glycosylation on melanoma migration. Antibodies against αvβ3 significantly reduced metastatic cell migration on VN (ca. 50% of control) but did not influence primary cell motility (Figure 5). Lectin PHA-L and both lectins with sialic acid specificity (SNA, MAA) blocked WM1205Lu cell motility but not WM793 cell motility (Figure 5) indicating that sialylated complex-type glycans with β1-6 branches are involved in metastatic melanoma migration. Lectin PHA-E and dMM inhibitor had no effect on the migration of melanoma cells on VN.
The faster wound closing by WM793 primary cells than by WM1205Lu metastatic cells (Figure 5) was not the effect of higher migration ability but rather the effect of the much larger area occupied by individual primary cells after scratching. We have also observed this effect in studies of WM793 migration on an uncoated surface and on fibronectin; an explanation for it has been given in previous publications (14, 15).
Discussion
While it is clear that the progression of many tumors is accompanied by increased αvβ3 integrin expression (10, 16), no fixed rules have been established on how αvβ3 glycosylation is altered in tumorigenesis (3). In the transition from the radial growth phase (RGP) to the VGP, rapid up-regulation of αvβ3 integrin expression has been shown to be required for melanoma cells to cross the basement membrane (BM) separating the epidermis from the dermis (11, 17-19). In our study, flow cytometry revealed high expression of the αvβ3 complex in both melanoma cell lines (ca. 99% of cells) (Figure 1). This is not surprising, considering that the WM793 and WM1205Lu lines represent the last two stages of melanoma progression; however, the transition from the primary to the metastatic stage requires changes allowing cells to cross into blood vessels, remain in circulation and form new lesions in distinct organs after leaving the circulation. It is known that the glycosylation of cell surface proteins must undergo alteration(s) in order for cancer cells to interact with new cell partners and adapt to new environments (20, 21). Rearrangement of glycans accompanies all steps of carcinogenesis, from transformation of normal cells (22, 23) to metastasis (15). To determine the glycosylation profile in the final stages of melanoma progression, we chose αvβ3 integrin, considered a biomarker of melanoma progression. PNGase digestion showed intensive N-glycosylation of αvβ3 integrin, especially the αv subunit in which nearly a third of the molecule is built of N-glycans (Figure 2A). We investigated the glycosylation pattern of αvβ3 by immunoblotting the αv and β3 subunits in lectin precipitates and by determining molecular mass loss after specific enzyme cleaving and in the presence of SW inhibitor. In both integrin subunits, we found significant differences between primary and metastatic melanoma cells in the content of high-mannose structures and sialylated complex-type glycans with bisected GlcNAc.
Oligomannose and hybrid-type N-glycans are immature structures; they are an intermediate step on the N-glycan synthesis pathway leading to the formation of complex-type structures. The content of high-mannose and hybrid type N-glycans in both αvβ3 integrin subunits was higher for WM793 primary cells than for WM1205Lu metastatic cells (Figures 2B, 3). These incompletely processed glycans are not frequently associated with cancer (24) but changes in their level during tumor progression have been shown in other studies (25-27). Up to now, however, there is no consensus as to the direction of these changes. De Leoz et al. (25) observed elevation of high-mannose-type glycans in mouse and human sera proteins during breast cancer progression. Among other alterations found by Ozcan et al. (26) in analyses of structural changes in serum glycoproteins, levels of oligomannose glycans in patients with gastric cancers were lower than in the control group. Kinoshita et al. (27) showed reduction of high-mannose glycans in melanoma cell lines during tumor progression; they found slightly lower expression of these structures in the WM266-4 metastatic cell line than in WM115 primary cells from the same patient. Glycosylation is a very sensitive process that depends on many factors, mainly the tissue-specific expression and activity of glycosyltransferases and glycosidases (28); different cancer-dependent effects on the glyco sylation profile, including high-mannose expression, are possible. So far, it is not clear whether these structures play a role in cell interaction (29). We did not observe differences in melanoma motility on VN after applying dMM, an inhibitor of N-glycan processing that blocks mannosidase I and leads to accumulation of high-mannose glycans (Figure 5). Zhang et al.'s work (30) did not disclose the functional significance of oligomannose structures but, based on comparable binding of dMM-treated H7721 human hepato carcinoma cells and control cells to human umbilical vein epithelial cells (HUVEC), concluded that high-mannose N-glycans do not mediate cell-cell interaction. Schraen-Maschke and Zanetta (31) determined that oligomannose and hybrid-type N-glycans are involved in homotypic aggregation and proliferation of C6 rat glioblastoma. Liwosz et al. (32) showed that the high-mannose/hybrid type N-glycans present on E-cadherin destabilize epithelial cell-cell interactions. A more detailed study is needed to find out whether the decrease of oligomannose glycans in αvβ3 integrin we observed during melanoma progression has a functional significance in melanoma behavior besides the behavior of migration on VN.
It is interesting that high-mannose and hybrid-type structures dominate in the β3 subunits (Figures 2B, 3) representing 73% and 83% of the total N-glycan pool in β3 from primary and metastatic cells, respectively, while, in the αv subunit, they account for only 36% in primary and 32% in metastatic cells. Other work also indicates β subunits as the main carriers of oligomannose structures, especially β1 glycoprotein (33). Kawano et al. (34) showed the presence of high-mannose structures on β1 subunit in mouse metastatic melanoma B16F1. Veiga et al. (35) found that oligomannose glycans are expressed only in the premature isoform of β1, of lower molecular weight, in B16F10 cells. The authors of the study showed that such glycosylated β1 precursors do not occur on the cell surface; however, other reports demonstrated the presence of such glycosylated β1 forms also in the cell membrane (36).
A number of studies have made clear the importance of sialylation of adhesion proteins in intercellular and cell-ECM contact and shown that any alteration of the sialic acid composition on the cell surface will change the cell's interactions (37, 38). We analyzed terminal sialic acids linked by α2-6 or α2-3 to complex-type N-oligosaccharides using the lectins SNA and MAA, respectively. Sialylation of α2-6 was more abundant in both subunits of primary cells and sialylation of α2-3 in metastatic cells (Figure 3). Neuraminidase digestion confirmed the higher content of α2-3-linked sialic acids in αvβ3 integrin from metastatic cells (Figure 2C). The role of α2-6 and a 2-3 sialylation in tumor progression has been examined previously (38). Cui et al. (39) found that high expression of α2-3-linked sialic acid was associated with the metastatic competence of breast cancer. Ranjan and Kalraiya (40) demonstrated that sialic acids linked by α2-6 to β1-6-branched complex-type glycans contribute to the invasiveness of melanoma cells by modulating B16F10 cell adhesion to ECM and BM. Some studies have assessed sialylation of integrin and the functional role of sialic acid alteration. Seals et al. (33) found that expression of the ras oncogene in HD3 colon epithelial cells raised α2-6 sialylation in β1 integrins but not in β3 and β5; it resulted in altered adhesion to collagen I. Previously, we demonstrated both types of sialic acids in αvβ3 integrin from metastatic melanoma cells (41). Citing Voura et al.'s (42) finding that the L1 molecule interacts with αvβ3 integrin on WM239 cells during their transendothelial migration, we suggested that the α2-3 sialic acids present on αvβ3 integrin in WM239 cells act as a ligand for the L1 molecule and that these interactions are crucial in transendothelial migration of melanoma cells. In our present model, WM1205Lu metastatic cells able to pass through the vascular endothelium showed elevated α2-3-linked sialic acids in αvβ3. We speculate that the role of these integrin structures in cancer cell extravasation, which we proposed in Kremser et al. (41), may also apply to WM1205Lu cells. Numerous studies suggest that sialic acid alterations are carcinoma-specific. Here we showed that α2-3- and α2-6-linked sialic acid expression on αvβ3 integrin changes during melanoma development.
Wound-healing assays showed the effect of αvβ3 integrin and α2-3- and α2-6-linked sialic acids only in WM1205Lu metastatic cell migration on VN. Inhibition of the migration of metastatic cells in the presence of SNA and MAA lectins was comparable, despite differences in α2-6 and α2-3 sialylation of αvβ3. This may mean that other α2-6-sialylated receptors are involved in the contact of metastatic melanoma cells with VN. Janik et al. (12) observed reduction of cutaneous melanoma migration on VN in the presence of both anti-αvβ3 and anti-αvβ5. Subunit β5 is also an N-glycosylated protein but its glycosylation is poorly known.
The β1-6 branching of both the αv and the β3 subunits, unlike sialylation, did not differ between primary and melanoma cell lines (Figures 2D, 3). The β1-6 branching of αvβ3 integrin remained unchanged despite an increase of β N-acetylglucosaminyltransferase V (GnT-V) enzyme expression during the transition from VGP to the metastatic phenotype observed in the same cell line pair in our previous study (15). This may mean that αvβ3 integrin is a substrate for GnT-V in an early stage of melanoma progression and that further change in the enzyme level during acquisition of the metastatic phenotype does not influence the β1-6 branching of this protein. Some studies strongly suggest that synthesis of GlcNAc β1-6-branched glycans begins in an early stage of cell transformation. In nude mice, Demetriou et al. (43) showed that normal lung epithelial cells transfected with GnT-V cDNA demonstrate a cancer-like phenotype due to decreased adhesion to fibronectin (FN) and collagen IV, loss of contact inhibition and increased tumorigenicity. Ito et al. (44) compared GnT-V expression between human hepatocellular carcinoma tissues and normal liver cells and also concluded that up-regulation of this enzyme appears in an early phase of carcinogenesis.
Interestingly, lectin precipitation and SW treatment revealed that αv chains possess much higher content of β1-6-branched structures than β3 glycoproteins in both analyzed cell lines (Figures 2D, 3). Demetriou et al. (43) showed that the αv subunit, not β3, was the substrate for GnT-V in Mv1Lu lung epithelial cells transfected with this enzyme. We found that β3 subunits are more abundant in high-mannose N-glycans than αv in both melanoma lines (Figures 2B, 3).
In cancer development, an increase of the number of antennae in complex-type N-glycans was observed resulting in synthesis of N-glycans of higher molecular weight (45). The data from our work do not confirm this observation. PNGase digestion showed a lower amount of N-glycans in both αvβ3 subunits from metastatic cells than from primary cells (Figure 2A). In both melanoma cell lines, the higher pool of N-glycans in αvβ3 from primary cells was the result of higher amounts of oligomannose structures (Figures 2B, 3) and, generally, more abundant sialylation (Figure 2C) but not the result of differences in branching of complex-type glycans (Figures 2D, 3).
Elevated β1-6 N-glycosylation is closely related to tumorigenesis and metastasis (46-48). In our study, PHA-L reduced WM1205Lu motility but not WM793 motility on VN indicating that β1-6 glycans acted only in metastatic melanoma migration on VN (Figure 5). Binding of both cell lines to VN was similarly stronger after culture in the presence of SW (Figure 4A), likely the result of having similar amounts of β1-6 glycans in αvβ3 integrin. It is well documented that malignant cells have altered adhesion, which facilitates cell migration to other sites in the body where they establish new tumors. Increased β1-6 branching is one of the best-characterized N-glycan alterations associated with malignant transformation and progression (49, 50). Our previous work showed that inhibition of β1-6 branching synthesis in the Golgi apparatus by SW (51, 52) or blocking of these structures on the cell surface by PHA-L lectin (41, 53) boosted cell adhesion to extracellular matrix proteins and impaired migration on ECM components.
In view of the similarity of β1-6 glycosylation of αvβ3 in primary and metastatic melanoma, why did we observe the involvement of β1-6 glycans only in metastatic cells migrating on VN? It might be due to the different glycosylation profile of αvβ3 integrin, which is involved in migration of metastatic but not of primary cells on VN (Figure 5). PHA-E precipitation clearly showed that complex-type N-glycans in both subunits of αvβ3 integrin from WM793 primary cells are modified by bisecting GlcNAc more strongly than in the WM1205Lu metastatic line (Figure 3). The significantly higher content of structures with bisecting GlcNAc in αvβ3 integrin from primary cells may be the reason why WM793 migration was suppressed, despite the high amount of β1-6 glycans in this integrin. N-oligosaccharides with bisecting GlcNAc have been shown to suppress cancer development (48, 54). An increase of bisecting GlcNAc structures has been found to be associated with loss of metastatic potential (21, 55). In a demonstration of antagonism between bisecting GlcNAc and β1-6-branched glycans, Zhao et al. (56) showed that GnT-V and GnT-III enzymes competitively modified α3β1 integrin, thus influencing α3β1-mediated cell migration. A recent work by Bubka et al. (57) did not exclude the presence of GnT-III and GnT-V products on the same glycoprotein in melanoma cells: over-expression of GnT-III leading to higher synthesis of bisecting GlcNAc was accompanied by higher β1-6 branching in cell glycoproteins, among them the melanoma cell adhesion molecule (MCAM). Taken together, these results suggest that the absence of WM793 primary melanoma migration on VN can be explained by the specifics of αvβ3 integrin glycosylation.
Previously, we established that inhibition of β1-6 branching synthesis by SW impaired both WM793 primary and WM1205Lu metastatic cell migration on FN (15). In the present experiments, blocking of β1-6-branched glycans by PHA-L lectin only reduced the migration of metastatic cells on VN. Apparently, the effect of β1-6-branched glycans on melanoma motility depends on ECM components that require interactions with other integrin receptors. Moreover, it is clear that the interactions of cancer cells with ECM proteins change during cancer progression and cell motility on VN via αvβ3 integrin seems to be most important during metastasis (6). Adhesive contact of cells with ECM proteins is needed to trigger the cell signaling pathways that control the metastatic phenotype. We observed clustering of αvβ3 integrin in focal adhesions of melanoma cells adhering to VN, differing from the dispersed expression seen on the whole membrane of cells growing on uncoated surface (Figure 4B). The ability of αvβ3 to cluster in focal adhesions on VN, unlike αvβ5 integrin, was described more than a decade ago (58). Clustering of αvβ3 in focal adhesion structures in the presence of VN points to the important role of α3β1-VN interactions in cancer cells.
Movement of cancer cells through the ECM requires the activity of matrix metalloproteinases (MMPs). Gil et al. (59) observed differences in MMP profiles between WM793 primary and WM1205Lu metastatic cells. In zymography tests, they found MMP-2 and MMP-9 activity in metastatic melanoma and only MMP-2 activity in primary cells. We showed high expression of αvβ3 in both of these cell lines but only the metastatic cells responded to αvβ3 monoclonal IgG, which blocked their motility on VN. It is well documented that αvβ3 integrin can regulate the expression and activity of MMPs in cancer cells (60-62) and that acquisition of the melanoma metastatic phenotype requires both elevated αvβ3 and the activity of MMPs (3). Here we also showed that, in melanoma, the malignancy process is accompanied by alteration of αvβ3 glycosylation, which also contributes to metastatic behavior.
It is worth noting that we observed reduction of melanoma cell migration on VN in the presence of the same clone LM609 of mouse antibody against αvβ3 that has been used in preclinical tests of the therapeutic drug Avastin. Several humanized, improved versions of this antibody have been tested in clinical trials as agents to block tumor angiogenesis and metastasis. It is thought that applying a dual therapy directed against αvβ3 and αvβ5, two integrins involved in tumor angiogenesis, will give better therapeutic results (8). The finding that αvβ3 glycosylation is significantly altered during melanoma progression may prove useful in the effort to develop more effective anti-cancer therapies.
Many studies on altered glycosylation in various cancers have examined the total pool of sera or cell glycoproteins, thus yielding information about general tendencies in tumorigenesis. Herein, we focused on one specific cell surface protein. We demonstrated, for the first time, that the pool of high-mannose and complex-type glycans with bisecting GlcNAc on αvβ3 integrin decreases during acquisition of the metastatic phenotype. We also observed a shift of αvβ3 sialylation from α2-6 in the primary line to α2-3 in metastatic melanoma cells. Future work has to focus on findings that may help in developing targeted therapies aimed at stopping the progression of melanoma, such as increased levels of αvβ3 integrin and alteration of αvβ3 N-glycosylation during acquisition of metastatic competence.
Acknowledgements
The Authors would like to thank Michael Jacobs for line-editing the manuscript for submission. This work was supported by a grant from the Polish Ministry of Science and Higher Education (grant no. N N301 0322 34). The Confocal Microscopy Laboratory (Institute of Zoology, Jagiellonian University) kindly gave us access to its LSM 510 META Axiovert 200M, ConfoCor 3 confocal microscope (Carl Zeiss MicroImaging GmbH, Jena, Germany).
- Received December 7, 2014.
- Revision received December 19, 2014.
- Accepted December 23, 2014.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved