

Abstract
Budding or the presence poorly differentiated clusters at the boundary of cancer tissue is a pathologically important finding and serves as a prognostic factor in colorectal cancer. However, few studies have examined the cancer tissue boundary in clinical samples. The purpose of the present study was to examine gene expression at the tumor front of colon cancer in surgically resected samples. Cancer tissues were obtained by laser microdissection of 20 surgically resected specimens. Genes with significantly different microarray signals between the tumor front and the tumor center were identified. Among genes showing significant up-regulation at the tumor front were six chemokines [chemokine c-c motif ligand (CCL)2 and -18, chemokine (C-X-C motif) ligand (CXCL)9-11, and interleukin 8 (IL8)], and two apoptosis-related molecules [ubiquitin D (UBD) and baculoviral iap repeat-containing 3 (BIRC3)]. Expression of laminin gamma 2 (LAMC2), matrix metallopeptidase 7 (MMP7) and epithelial–mesenchymal transition (EMT)-related molecules were elevated in the tumor front, but their fold changes were smaller than those of the aforementioned genes. These results suggest that chemokines, in addition to EMT-related molecules, may play important roles in invasion of colon cancer.
- Gene-expression profiling
- colon cancer
- tumor front
- chemokines
- epithelial–mesenchymal transition
- microarray
Budding and the presence of poorly differentiated clusters scattered in the stroma at the invasive tumor margin are pathologically important findings as predictors of lymph node metastasis and poor prognostic factors (1). Few basic studies have examined the site of cancer invasion, including budding and poorly differentiated clusters, and screening of molecular features related to cancer invasion has rarely been performed in clinical samples (2).
During invasion, cancer cells often change from an epithelial to a mesenchymal type through a pathway involving the so-called epithelial–mesenchymal transition (EMT). Recent studies have shown that this pathway involves transforming growth factor-beta, epidermal growth factor and fibroblast growth factor (3). Several studies have examined the expression status of these molecules at the tumor front (TF) and have shown elevated expression of EMT-related molecules (3-5), suggesting that EMT plays a crucial role in cancer invasion. However, other molecules whose expression differs at the TF are mostly unknown. Thus, the goal of the present study was to identify molecules with differential expression at the TF.
Materials and Methods
Patients. Patients referred to the Department of Surgery, Kyorin University Hospital between April 2010 and November 2011, aged 50-80 years, had well- or moderately-differentiated adenocarcinoma, and were scheduled for open laparotomy for colorectal cancer, were considered eligible for the study. Exclusion criteria included previous chemotherapy or radiotherapy for any carcinoma and treatment or history of familial adenomatous polyposis, hereditary non-polyposis colorectal cancer and inflammatory bowel disease.
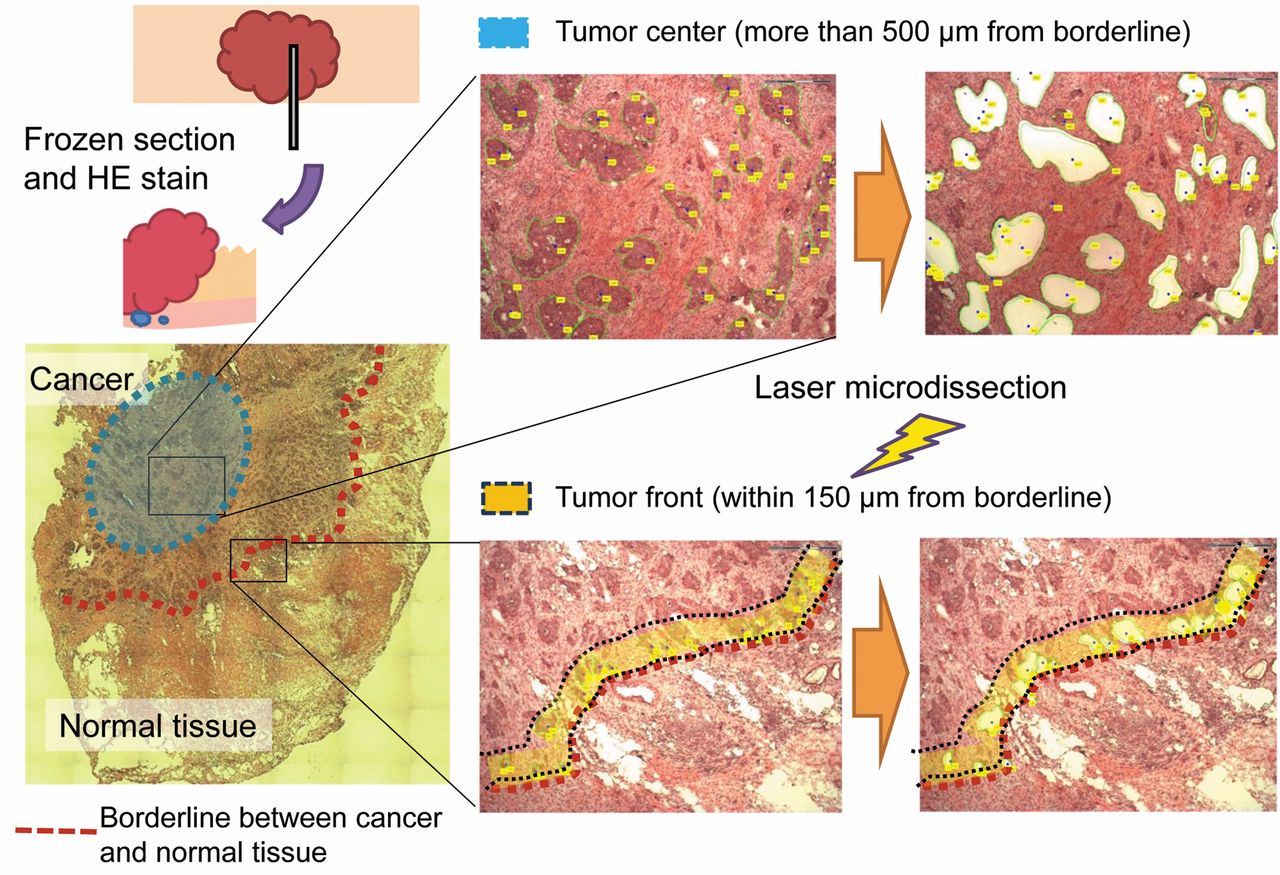
Samples and microdissection. Tissues samples were frozen in OCT (Sakura Finetek, Tokyo, Japan). The TF was defined as the region <150 μm from the border of the cancer and the tumor center (TC) was defined as >500 μm from the border. Cells at the TF and the TC in cancer tissues and epithelial cells from normal mucosa were obtained using laser microdissection (Carl Zeiss, Jena, Germany) (Figure 1).
RNA extraction, reverse transcription and amplification. Total RNA was isolated using an RNeasy Mini kit (Qiagen, Chatsworth, GA, USA) and the quality was checked using a Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). Only samples with RNA integrity number >7.0 were used in analysis.
Quantities of RNA were very small; an Ovation Pico WTA containing oligo dT primer and random primer (Nugen, San Carlos, CA, USA) and a TaqMan® PreAmp Master Mix (Life Technologies, Carlsbad, CA, USA) were used for pre-amplification with an Mx3000P (Agilent Technologies).

Process for obtaining cancer cells from each location by laser microdissection.
Microarray analysis. Gene-expression profiles at TF and TC of each patients were determined using Human Gene ST 1.0 (Affimetrix, Santa Clara, CA, USA). Background subtrac¬tion and normalization using the robust multi-array average algorithm method was performed using GeneSpring GX 12 (Agilent Technologies). Fold-change values for genes were calculated as the ratio of the signal values of the target site compared with the control site. The Benjamini-Hochberg false-discovery rate was used for multiple testing corrections. Microarray data were submitted to the Gene Expression Omnibus with accession number GSE65480.
Real-time RT-PCR. mRNA expression levels of selected and EMT-related genes were analyzed with pre-amplified cDNA, Brilliant III® Fast Master Mix (Agilent Technologies), and Taqman probes (Applied Biosystems), following the manufacturer's instructions. β-Actin (ACTB) expression was used as an internal control and the fold change of target genes was expressed relative to this control.
Results
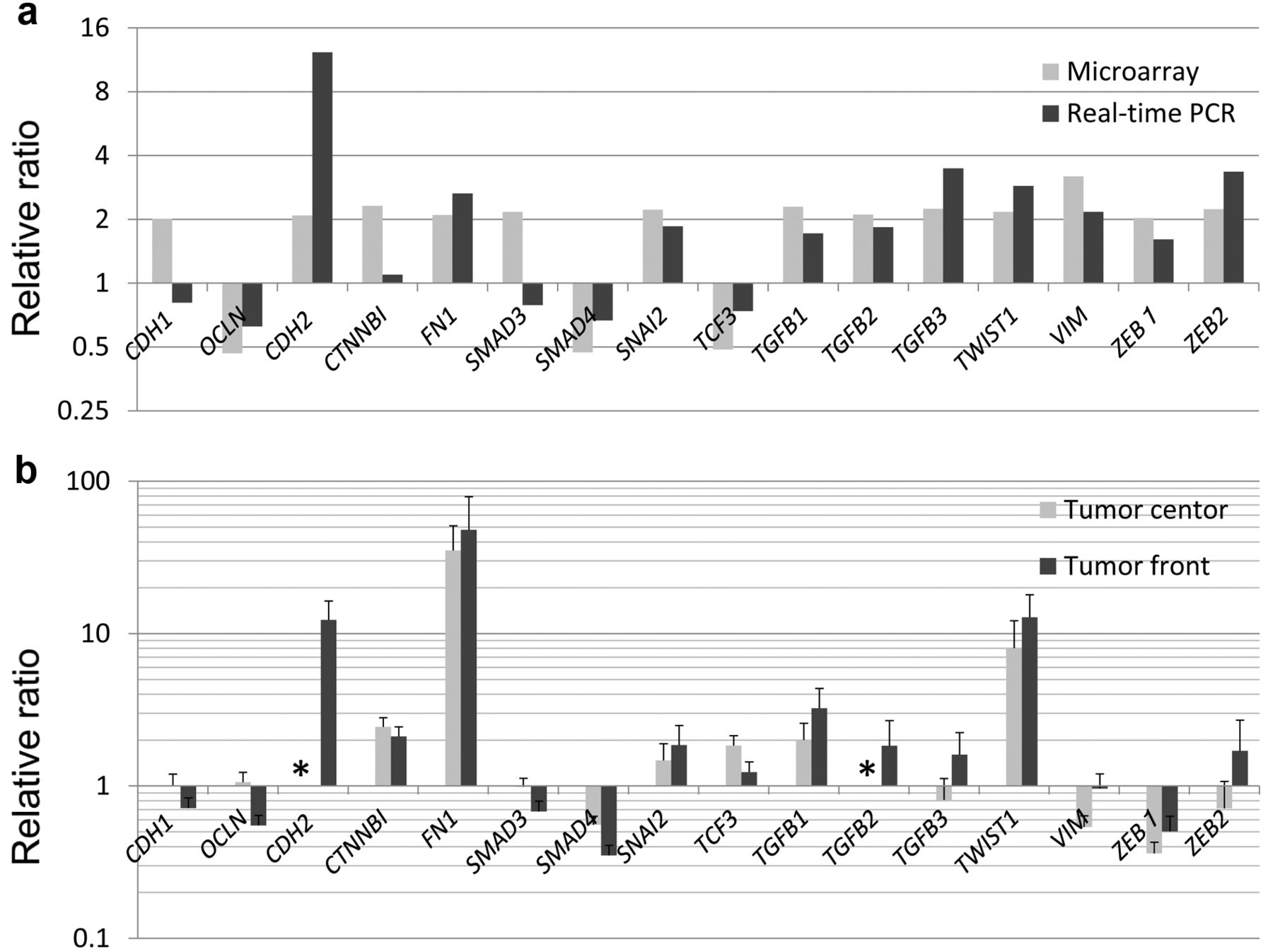
Patients and pathological findings. From April 2009 to December 2011, 20 patients were recruited into the study (Table I). Out of these patients, 11 had a poor differentiation at the TF. There was no significant difference in clinicopathological findings among cases based on pathological type, stage, or level of differentiation at the TF. Site-specific expression at the TF. There were 17 genes with a fold change of ≥4 in the expression level at the TF with p<0.001 in microarray analysis. Among these genes, we focused on those with known functions, including six chemokines, two apoptosis-related molecules, and two extracellular matrix and degrading enzymes (Table II). Elevated expression at the TF in microarray analysis was confirmed using qPCR for all genes examined (Figure 2a). Genes with a higher expression at the TF compared to those in the TC also had higher expression in the TF than in normal mucosa (Figure 2b).
Patients' characteristics.

Gene expression at the invasive front determined by microarray analysis and real-time polymerase chain reaction. CCL18: Chemokine c-c motif ligand 18; CXCL8-11: c-x-c motif chemokine 8-11; MMP7: matrix metallopeptidase 7; UBD: ubiquitin d; LAMC2: laminin gamma 2; BIRC3: baculoviral iap repeat containing 3; CCL2: chemokine c-c motif ligand.
Microarray data.

{kind=link}
{kind=link}
{kind=link}
Comparison of gene expression in normal mucosa and cancer tissue by real-time polymerase chain reaction. Ratios are shown for expression in cancer tissue to that in normal mucosa. The bars show relative ratios of expression of each gene with the mean standard error. Expression levels of C-X-C motif chemokine 10 (CXCL10) and matrix metalloproteinase 7 (MMP7) are shown as ratios relative to expression at the tumor center, since there was almost no expression of these genes in normal mucosa. CDH1: E-cadherin; CDH2: N-cadherin; OCLN: occluding; CTNNB1: catenin (cadherin-associated protein), beta 1; FN1: fibronectin 1; SMAD3/4: smad family member 3/4; SNAI2: snail family zinc finger 2; TCF3: transcription factor 3: TGFB1-3: transforming growth factor, beta 1; TWIST1: twist family bhlh transcription factor 1; VIM: vimentin; ZEB1/2: zinc finger e-box binding homeobox 1/2.
Gene expression of EMT-related molecules at the TF. In microarray data, expression of markers of increased EMT (6, 7) was higher at the TF than in the TC. In contrast, a marker for decreased EMT, occludin (OCLN), had a lower expression at the TF than in the TC, although that for E-cadherin (CDH1) was not lower at the TF. Of note, none of these genes fulfilled the criteria of fold change ≥4.0 used for selection of genes with a significant change of expression level in microarray analysis (data not shown). These changes in expression levels were confirmed by qPCR for all genes except CDH1 and SMAD family member 3 (SMAD3) (Figure 3a). Expression levels of fibronectin 1 (FN1) and TWIST family BHLH transcription factor 1 (TWIST1) were increased in cancer tissues compared to normal mucosa (Figure 3b).
Discussion
The importance of this study lies in the identification of genes showing TF-specific changes through screening of clinical samples. Expression of six chemokines (CCL18, CXCL9-11, IL8 and CCL2), two apoptosis-related molecules (UBD and BIRC3), and LAMC2 and MMP7 increased at the TF. To the best of our knowledge, this study is the first to identify these molecules as showing changes in expression at the TF in colon cancer. It is noteworthy that expression of many chemokines was significantly higher at the TF than in the TC.
Tissue expression of CCL18 is correlated with prognosis in colorectal cancer (8), and it has also recently been suggested to be an inducer of EMT in lung cancer (9). Chemokines CXCL9-11 act on a common receptor, CXCR3, and increase the migratory ability of cells (10). Another chemokine, IL2, also increases the mobility of cancer cells (11).
In addition, expression of EMT marker molecules also changed slightly at the TF. Previous studies have shown high expression levels of β-catenin (12) and laminin-5γ2 (13) and a low expression level of E-cadherin (14) at the TF. Our results are mostly consistent with these studies. While expression of several EMT-related molecules changed in microarray analysis, they did not fulfill selection criteria because of the small fold change. Since many molecules exhibit crosstalk with EMT-related molecules in the EMT pathway (15), it will be necessary to analyze the association between chemokines and EMT-related molecules.
The present study has several limitations. Firstly, contamination with some other cells by laser microdissection was unavoidable and we could not study interactive effects with surrounding cancer cells. Secondly, a clear definition of TF and TC has not been established. We assumed that a distance of 150 μm was required to ensure that only cancer cells at the TF were collected because the smallest cancer ducts were about 50 to 100 μm. We also provisionally defined the TC as >500 μm from the border because a region of about three times the area of the TF does not contain TF.
Within these limitations, our results revealed that the expression of chemokines and EMT-related molecules changes at the TF, and that these changes are greater for chemokines. These results suggest that chemokines and EMT-related molecules play crucial roles in cancer invasion. Further study is required to clarify the precise mechanism by which these molecules promote cancer cell invasion.
Acknowledgements
This work was supported by the Japan Society for the Promotion of Science (Kakenhi grant 21791305 to T. K.).
The Authors thank Professor Sakurai and Dr. Otsuka.
Footnotes
Conflicts of Interest
The Authors have no conflict of interest.
- Received September 20, 2015.
- Revision received October 16, 2015.
- Accepted October 26, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved