

Abstract
Background/Aim: Melanoma cells express high levels of HLA class II, cell surface antigen-presenting proteins, which is an anomalous phenotype among solid tumors. There has never been a satisfying explanation for how this HLA class II-positive phenotype is related to tumor development. Lugini and colleagues demonstrated that melanoma cells have the capacity to engulf T-cells. We considered the possibility that this capacity could be dependent on HLA class II expression. Materials and Methods: We co-cultured melanoma and CD4-positive, labeled, Jurkat-C T-cells. The melanoma cells were transformed with an expression vector for CIITA, the obligate HLA class II gene transactivator. We then assayed for the transfer of label to the melanoma cells. Results: CIITA expression facilitated engulfment of the T-cell material but not material from B-cells. Conclusion: The results suggest a possible mechanism for HLA class II-positive melanoma cells in blunting an anti-tumor response and suggest a possible target for melanoma therapy.
- HLA class II
- T-cell
- CIITA
- melanoma
- immune response
- Jurkat-C
Melanoma cells exhibit a curious and significant phenotype that is almost unique among solid tumor cells: A large percentage of melanoma cells, represented by either isolated tumor cell lines or different biopsy specimens, are constitutively positive for the HLA class II antigen-presenting molecules (1-3). Tumor cells representing most, if not all, other solid tumors do not constitutively express HLA class II. Such cells are limited to HLA class II induction by interferon-gamma (IFN-γ) unless the cells have IFN-γ signaling pathway defects, which are indeed common among solid tumor lines (4-7).
Constitutive expression of HLA class II in melanoma cell lines requires activation of one of the promoters for class II transactivator (CIITA) by constitutive activation of the mitogen activated protein kinase (MAPK) pathway (8,9). MAPK is considered an important driver of tumorigenesis in melanoma. CIITA expression may occur due to the direct effects of malignant transformation. However, it has also been proposed that constitutive CIITA expression in melanoma cells is due to fusion of tumor cells with CIITA-constitutive macrophage (10-12). CIITA expression has been linked to a reduction in apoptosis (13). In fact, other solid tumors, besides melanoma, express CIITA but without the corresponding high level of HLA class II seen in melanoma cell lines and in many patients' biopsies (14). CIITA is involved in B-lymphoma translocations but with the apparent effect of HLA class II down-regulation, proposed as a possible mechanism of blunting the anti-tumor response (15).
Materials and Methods
Cells and HLA-DR staining. Cell culture and antibody staining for flow cytometry for detection of HLA-DR were as described (16). Briefly, melanoma and lung cells were maintained in RPMI, 10% fetal calf serum, penicillin, streptomycin and glutamine. To assay for surface HLA-DR, cells were scraped, recovered by centrifugation and re-suspended in 1% human serum in PBS to block Fc-receptors. Commercially available PE-labeled, anti-HLA-DR (Pharmagen) or isotype control was added according to vendor's instructions for one hour at 4°C. Cells were recovered by centrifugation and re-suspended in 1% human serum in PBS for flow cytometry.
Jurkat-C eFluor-670 labeling and cell co-culturing for fusion assays. Jurkat-C were labeled according to vendor's (eBioscience) instructions and then immediately added to a 50% confluent plate of adherent cells. Approximately 1 million labeled Jurkat-C cells were added to 10 ml of media in a 100 mm plate of adherent cells. Cells were co-cultured for 48 hours before the assay for adherent cell uptake of eFluor-670, either by microscopy or flow cytometry.
Microscopy. Photomicrographs were acquired with a Zeiss (Zeiss Microsystems, Manheim, Germany) AxioObserver inverted fluorescent microscope; enclosed in a full incubation environment controlled at 37°C and 5% CO2; with a Zeiss MrM monochrome CCD detector; through a 20X/0.5NA Plan NeoFluor dry objective; and viewed, prepared and exported with the Zeiss Axiovert v4.8.3 software suite (Zeiss) (www.zeiss.com). Phase contrast and Cy5 (620nm/60 excitation; 700nm/75 emission) images were acquired sequentially and the fluorescent images were pseudo-colored red. Exposure time (1,500 ms), gain, offset and similar acquisition settings were identical for all samples. Each image set was repeated five times.
Image analysis. Raw Zeiss .zvi files were exported as merged and individual channel uncompressed .tif files. The merged phase contrast and red pseudo-colored images were imported into Definiens Developer v2.0 (Definiens, Munich, Germany). A morphological filter rule set was designed to isolate and classify the small, round, suspended T-cells and the larger, elongated, flat cancer cells. The cancer cells were further segmented into individual cells using the phase contrast membrane definition as a border to a shrink and grow process. The number of adherent cancer cells was quantified for each sample for all five repetitive image sets. The amount of red positivity for each cell was subsequently quantified continuously and all cells with a region larger than 10 pixels and also presented with expression of red fluorescence greater than a threshold of 150 (dynamic range for the 8 bit image) were considered red positive. The percentage of red positive adherent cells to all adherent cells was calculated.
Flow cytometry. Samples were assayed with a BD LSR II flow cytometer (Becton Dickinson). Tumor cells and lymphocytes were distinguished on forward and side scatter plots. eFluor-670 labeled cells were excited with a 633nm laser and the fluorescence emission was detected in the 660/20 nm range. Results were analyzed with the BD Facsdiva Software V6.1.3 and mean, as well as median fluorescent values associated with cell populations were exported to Microsoft Excel files.
Results
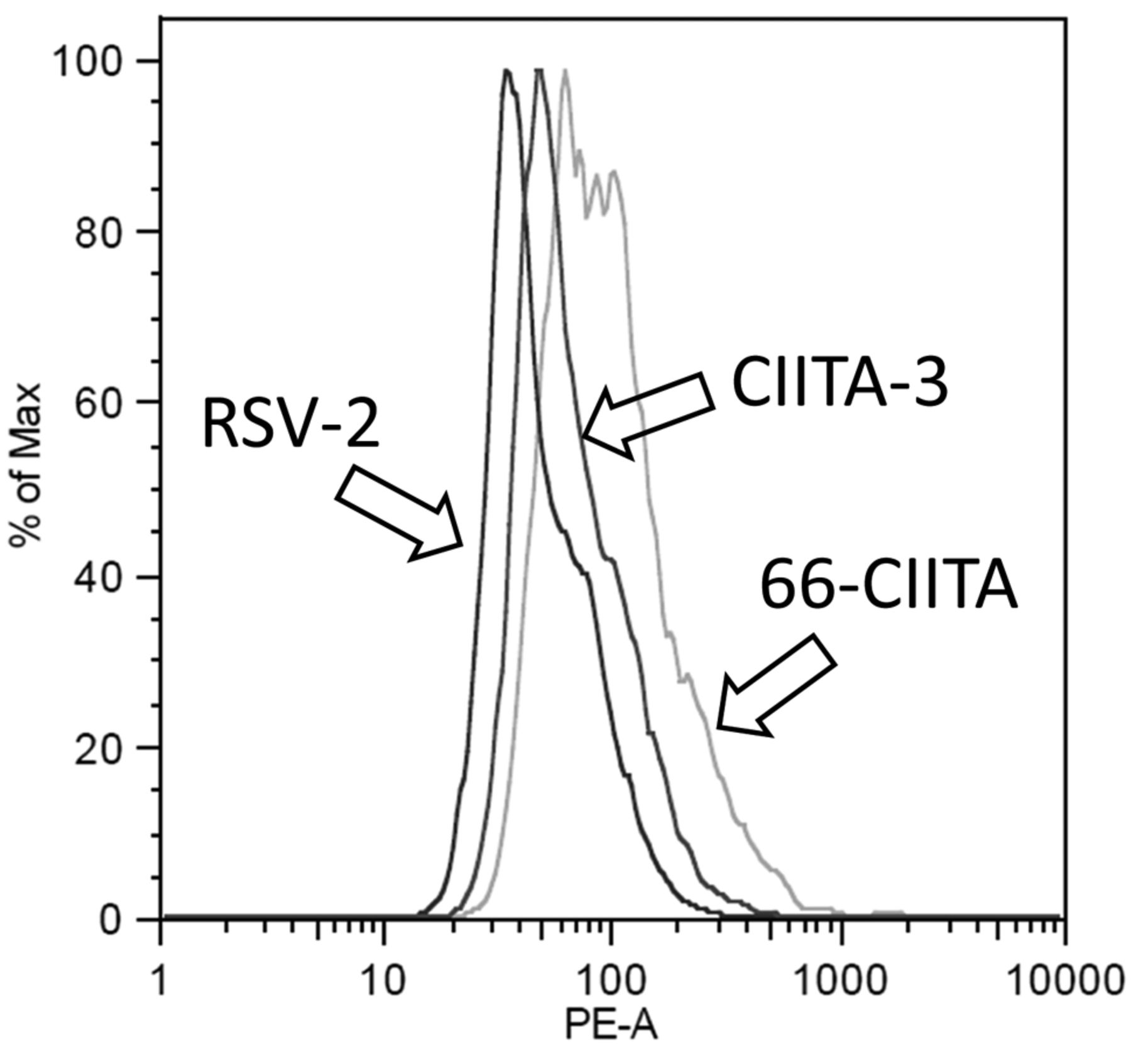
Lugini et al. (17) observed that melanoma cells have the capacity to consume T-cells; however, their work did not suggest any relevant mechanism. This function adds a new dimension to the ways tumor cells interact with the immune system, in addition to evasion or co-option (18), and provides a role for HLA class II that could be opposite to HLA class II's presumed role of improving the anti-tumor response in other settings (15, 19). Herein, we investigated the possibility that the common expression of HLA class II on melanoma cells provides the trait permitting T-cell engulfment by the melanoma cells. This hypothesis is difficult, if not impossible, to test using HLA class II blocking antibodies because of the capacity of such antibodies to facilitate HLA-DR-mediated apoptosis (16, 20). Thus, we transformed (originally HLA class II-negative) melanoma cells using an expression vector for CIITA and verified expression of HLA-DR; the canonical HLA class II protein (Figure 1) (16).

Detection of HLA-DR (Phycoerithryn-labeled) in CIITA transformants by flow cytometry. Cells were stained with PE-labeled, commercially anti-HLA-DR exactly as described (23,24). 66-CIITA (CIITA transformant of H2009 NSCLC; right histogram), CIITA-3 (middle histogram) and RSV-2 (left histogram).
We labeled CD4-positive, Jurkat-C cells with eFluor-670 and co-cultured these cells with CIITA-3, transformant of 1286 melanoma cells, isolated as described elsewhere (16). Previous work has indicated that CIITA-3 not only expresses HLA-DR (Figure 1) but this expression is functional in an HLA-DR-dependent apoptosis assay (16). We also mixed the labeled Jurkat-C cells with H2009 non-small cell lung cells (NSCLC) that had been transformed with the CIITA expression vector (66-CIITA) and, thereby, expressed HLA-DR (Figure 1), as well as with the control, 1286 melanoma cells, transformed with an empty expression vector, termed RSV-2. We observed uptake of the eFluor dye by the adherent cancer cells, as indicated in Figure 2 and we quantified the microscope images, which indicated that both CIITA transformants had more eFluor dye than the RSV-2 cells (Table I). The CIITA-3 melanoma cells displayed fluorescence in 9.1% of the cells compared to 4.2% of the RSV-2 HLA class II-negative cells (p<0.01). Remarkably, 45.8% of H2009 CIITA-transformed cells (66-CIITA) expressed eFLuor dye, consistent with the higher level of HLA-DR expression in these cells (Figure 2) (p<0.0005 compared to 4.2% RSV-2). However, the H2009 cells are not melanoma cells and, despite the increased level of HLA-DR on these cells, it is possible that other factors play a role in the dramatically high level of eFluor uptate, such as a higher level of H2009 phagocytosis.
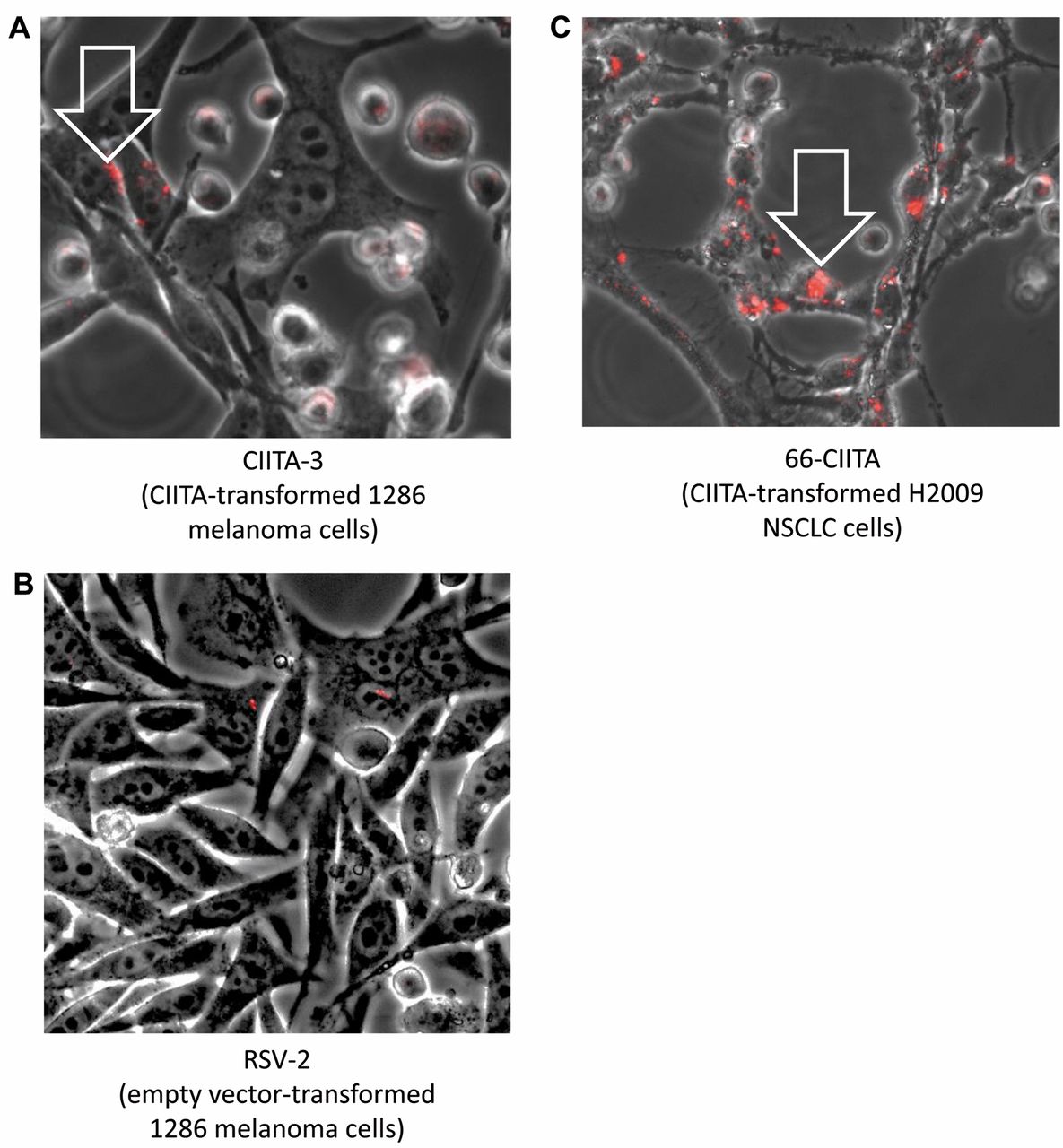
Microscopic detection of eFluor-670 in adherent tumor cells at ×200 total magnification in live cells. Adherent CTIIA-3 transformed cells display significant internalized punctate red fluorophore (white arrow) (A); whereas RSV-2 (empty vector, control transformed) cells confirmed a lack of fluorophore detection (B). 66-CIITA similarly presented with internalized red punctate staining (C).
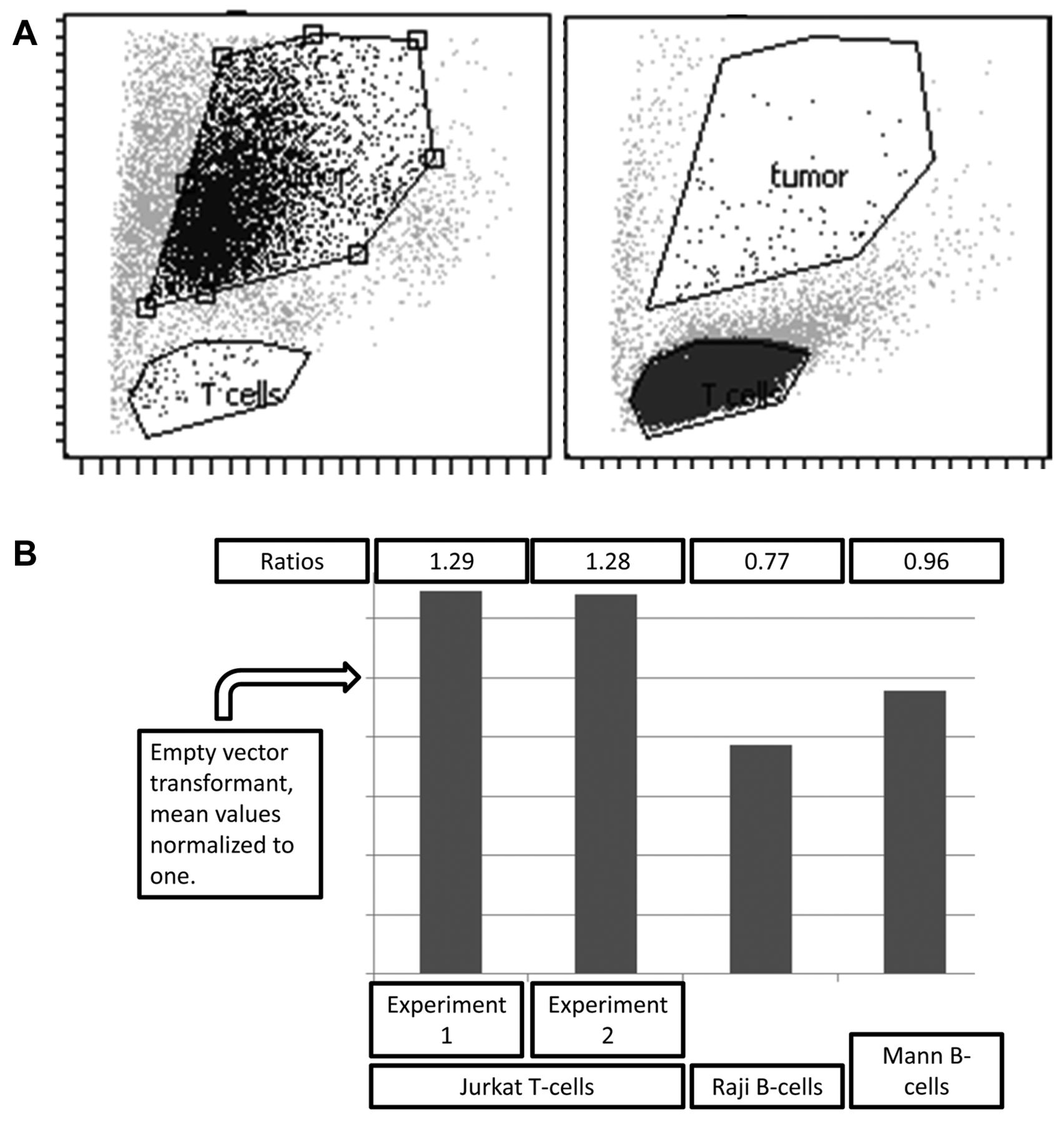
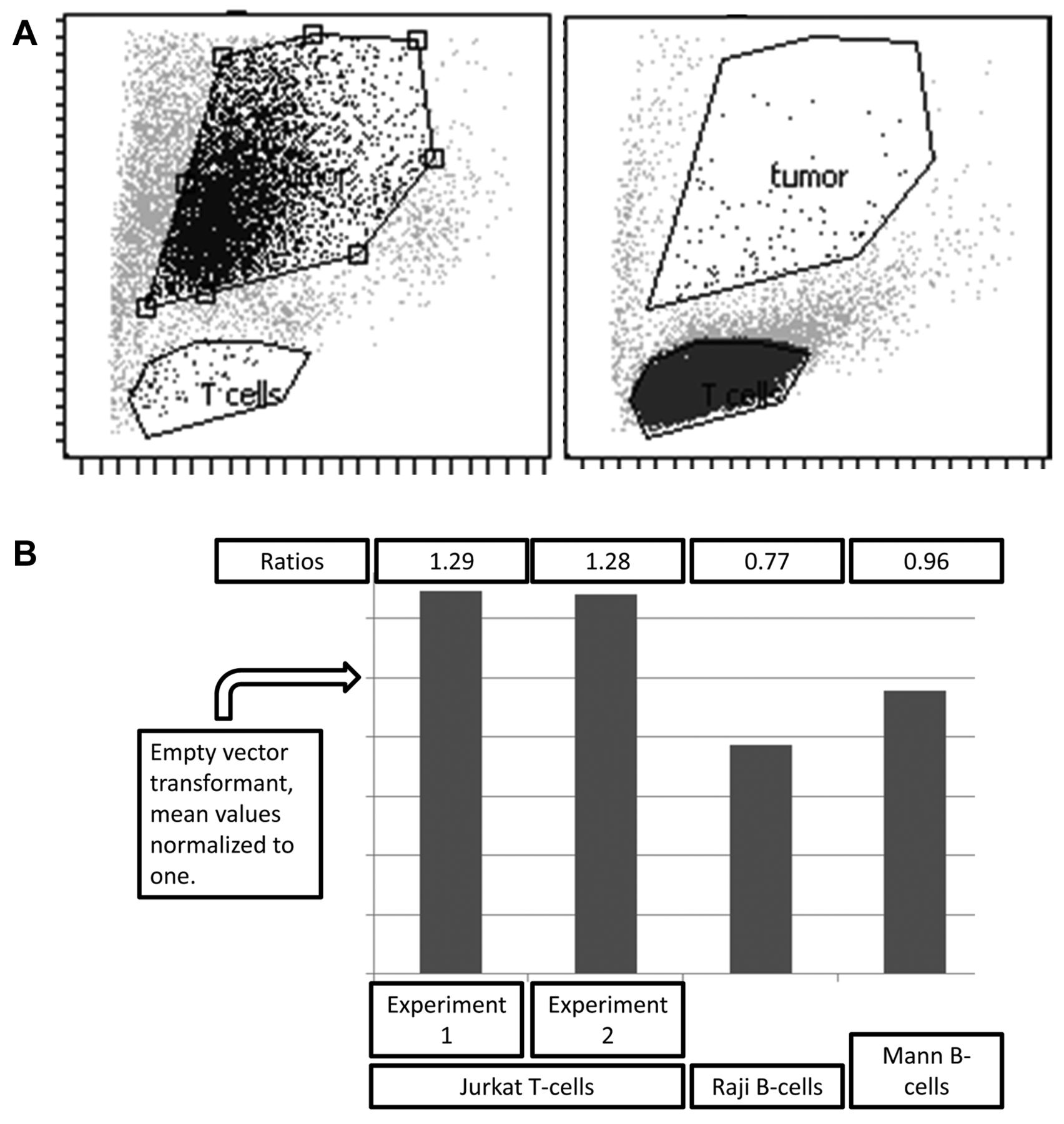
We next assayed for eFluor uptake by flow cytometry (Figure 3), which indicated that the presence of HLA class II led to a 30% increase in uptake of Jurkat-C eFluor by the CIITA-3 melanoma cells but had no effect on uptake of material from two eFluor labeled B-cell lines, Raji and Mann, which lack CD4 or other HLA-class II binding; T-cell receptor components.
Discussion
The above data are consistent with the conclusion that HLA class II expression facilitates uptake of Jurkat-C T-cell material, raising the question of whether HLA class II expression on melanoma cells in patients would affect the T-cell response against the melanoma cells. Also, these data provide a potential mechanism for theories of metastasis, whereby lymphocyte-tumor cell fusions are at the heart of tumor cell migration (10, 21, 22). However, it should be noted that CIITA expression can lead to other melanoma cell alterations and, despite the work by Lugini and colleagues (17), there are potential, alternative processes that could lead to the transfer of eFluor dye to the melanoma cells, such as exosome-mediated transfer or other processes of debris engulfment.
{kind=link}
{kind=link}
{kind=link}
Detection of eFluor-670 uptake by tumors using flow cytometry. Flow cytometry gates indicating detection of eFluor-670 in the adherent tumor cells versus Jurkat-C T-cells. The two plots (3A) represent isolated cells indicating the function of the gates for detection of each cell type, respectively, and, in particular, indicating lack of gate overlap. Bar graphs (3B) indicating increased level of eFluor-670 uptake by the CIITA-3 cells, compared with the RSV-2 cells, in two separate experiments (where the value for the RSV-2 cells is normalized to one, indicated by arrow head, y-axis).
Percent of eFluor-670 labeled tumor cells following incubation with eFluor-670 labeled Jurkat-C T-cells.
- Received September 3, 2014.
- Revision received September 25, 2014.
- Accepted September 30, 2014.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved