

Abstract
Background: Cluster of differentiation 147 (CD147)/basigin on the malignant tumor cell surface is critical for tumor proliferation, invasiveness, metastasis, and angiogenesis. CD147 expressed on malignant melanoma cells can induce tumor cell invasion by stimulating the production of matrix metalloproteinases (MMPs) by surrounding fibroblasts. Membrane vesicles, microvesicles and exosomes have attracted attention, as vehicles of functional molecules and their association with CD147 has been reported. Cleaved CD147 fragments released from tumor cells were reported to interact with fibroblasts. We investigated the intercellular mechanisms by which CD147 stimulates fibroblasts to induce MMP2 activity. Materials and Methods: CD147 was knocked-down using short hairpin RNA (shRNA). The stimulatory effect of CD147 in cell culture supernatants, microvesicles, and exosomes on the enzymatic activity of MMP2 was examined by gelatin zymography. Results: Supernatants from A375 control cells induced increased enzymatic activity of fibroblasts; such activity was significantly lower in CD147 knock-down cells. Conclusion: Cleaved CD147 plays a pivotal role in stimulating fibroblasts to induce MMP2 activity.
- CD147
- malignant melanoma
- matrix metalloproteinases
- microvesicles
- exosomes
Cluster of differentiation 147 (CD147)/basigin, an integral plasma membrane protein belonging to the immunoglobulin (Ig) superfamily, is widely expressed in various tissues. It is particularly enriched on the surface of malignant tumor cells, including malignant melanoma (MM) cells (1-6). MM is characterized by its highly aggressive nature and high potential for invasion and metastasis. In previous studies we documented that CD147 expressed on MM cells plays an important role in tumor proliferation, invasiveness, and metastasis, and in vitro and in vivo angiogenesis (3, 7, 8).
Matrix metalloproteinases (MMPs) are a family of enzymes that degrade different components of the extracellular matrix, and a number of studies indicate that they play an important role in tissue remodeling, tumor invasion, and metastasis (9, 10). Earlier studies have reported the involvement of MMPs in the migration, invasion, metastasis, and progression of MM. The elevated expression of MMP1, MMP2, and MMP9 was correlated with the migration and invasion of cultured human MM cells (10, 11). Our previous immunohistochemiical studies and invasion assays using transwell Boyden chambers showed that CD147 expressed on MM cells induced tumor cell invasion by stimulating the production of MMPs by fibroblasts (2) that were not in direct contact with MM cells, indicating the presence of intercellular communication between MM cells and fibroblasts. These communications primarily involve soluble factors such as cytokines, chemokines and growth factors, and their specific cell-surface receptors. Another mode of intercellular communication via membrane vesicles is of increasing interest (12). There are two types of membrane vesicles, microvesicles and exosomes; they are vehicles of functional molecules and may be involved in several physiological processes. Microvesicles are formed from the plasma membrane and released by surface shedding. Exosomes derive from exocytosis of cytoplasmic multivesicular bodies; an association between microvesicles and CD147-mediated tumor-stromal interactions have been documented (13-15). On the other hand, Egawa et al. demonstrated that cleaved fragments of CD147 are released from tumor cells independently of membrane vesicles (16).
In this study, we investigated the intercellular mechanisms that play a role in the stimulation by CD147 of fibroblasts to induce MMP2 activity. We examined whether the cleaved form of CD147 or membrane vesicles are involved.
Materials and Methods
Cell culture. We purchased the human MM cell line A375 from American Type Culture Collection ATCC (Manassas, VA, USA) and normal human dermal fibroblasts (NHFBs) from Kurabo (Osaka, Japan). All cells were grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin-amphotericin B solution (Invitrogen, Carlsbad, CA, USA) and incubated at 37°C in a humidified atmosphere containing 5% CO2.
Lentivirus infection. U6-vshRNA-CMV-PUR lentivirus encoding CD147 short hairpin RNA (shRNA) (CD147 shRNA lentivirus) was constructed and produced by Shanghai GeneChem Co. Ltd. (Shanghai, China) (17). The sequences were sequence 1-1, '-TGTCGTCAGAACACATCAACTTCAAGAGAGTTGATGTGTTC TGACGACTTTTTTC-3’; sequence 1-2, 5’-TCGAGAAAAAA GTCGTCAGAACACATCAACTCTCTTGAAGTTGATGTGTTCT GACGACA-3’. Infection was according to the manufacturer's instructions. Briefly, A375 cells were seeded in 12-well plates in 1000 μl of growth medium without antibiotics. When the cells reached 70% confluence in about 24 h, they were infected with either empty lentivirus (control; CD147 control lentivirus) or CD147 shRNA lentivirus. The cells were incubated with virus for 4 h at 37°C and the medium was then replaced with 2 ml of fresh medium. The infection of cells with CD147 control or CD147 shRNA lentivirus was confirmed by their resistance to puromycin (Sigma-Aldrich Corp, St. Louis, MO, USA). Cells were exposed for 7-10 days to medium containing 400 ng/ml puromycin, maintained for at least two weeks in medium containing 200 ng/ml puromycin, and then stable cell clones were harvested.
Preparation of microvesicles and exosomes. Microvesicles and exosomes were prepared as described elsewhere (18). Briefly, cells were cultured with serum-free DMEM for 48 h and then 400 ml of the culture medium were harvested and centrifuged (500 × g, 10 min) to remove cells and large debris. The supernatant was centrifuged at 20,000 × g for 20 min to obtain pellets containing microvesicles. Exosomes were harvested by centrifuging the supernatant at 100,000 × g for 70 min (18). The medium after removing cells, cell debris, microvesicles, and exosomes was used as supernatant (Figure 1).
Electron microscopy. Microvesicles and exosomes were fixed for 30 min at 4°C in 2% osmium tetroxide solution (TAAB, Berkshire, UK) and then post-fixed in 2.5% glutaraldehyde and 2% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for 30 min at room temperature. Samples were dehydrated by passing through a graded ethanol series and embedded in Epon 812 (TAAB). Ultrathin sections were mounted on copper grids, contrasted with uranyl acetate and lead citrate, and observed under an electron microscope (Hitachi H-7650; Hitachi High-Technologies Co., Tokyo, Japan).
Western blotting. Total protein was isolated from the cultured cells. Briefly, after washing three times with ice-cold phosphate-buffered saline, the cells were suspended in lysis buffer (150 mmol/l NaCl, 10 mmol/l Tris, 0.1% sodium dodecylsulfate, 1.0% Triton X-100, 1% deoxycholates, 5 mmol/l ethylene diaminetetra acetate) on ice for 30 min and then subjected to centrifugation. The supernatant was concentrated using an Amicon Ultra-15 device with a 10-k molecular weight (MW) cut-off (Millipore, MA, USA). The protein concentration was measured with bicinchoninic acid protein assay (Pierce, Rockford, IL, USA) according to the manufacturer's instructions. Equal amounts of proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and electrotransferred to polyvinylidene fluoride membranes. Non-specific binding sites were blocked overnight at 4°C in Tris-buffered saline containing 5% non-fat dry milk and 0.1% Tween-20. The blots were incubated for 1 h at room temperature with primary antibodies against CD81 (dilution 1:1,000; System Biosciences, Mountainview, CA, USA) or human CD147 (dilution 1:2,000; Invitrogen) and then for 1 h at room temperature with the secondary antibodies. The signals were visualized using a SuperSignal West Pico chemiluminescent substrate (Thermo; Shibuya, Tokyo, Japan).
Zymography. Gelatinolytic activities in conditioned media were demonstrated by zymography using gelatin as a substrate. Microvesicles and exosomes obtained from 400 ml of culture medium were resuspended in 400 ml of fresh medium each. NHFBs were seeded at a density of 2.5×104 cells/well in 24-well flat bottom plates and incubated with supernatants, microvesicles, and exosomes for 72 h. Samples (10 μl) were electrophoresed by 9.5% SDS-PAGE on gels containing 1 mg/ml gelatin. After electrophoresis, the gels were incubated at room temperature for 1 h in a solution containing 2.5% Triton X-100 and 10 mM Tris-HCl (pH 8.0) to remove SDS, incubated at 37°C for 20 h in reaction buffer [50 mM CaCl2, 10 μM ZnCl2, 50 mM Tris-HCl (pH 8.0)], and stained with 1% Coomassie blue R250 in 10% ethanol and 5% acetic acid. Enzyme activity was visualized as a colorless band on a blue background.
Statistical analysis. Differences between two groups were evaluated by the t-test. Dunnett's procedure was used for multiple comparisons against the control. All reported p-values are two-tailed. Differences of p<0.05 were considered significant.
Results
shRNA-mediated silencing of CD147 expression in A375 cells. Specific shRNA targeting CD147 was used for knock-down of CD147 expression. Western blots showed that CD147 expression was significantly lower in A375 cells transfected with CD147 shRNA lentivirus than CD147 control lentivirus (Figure 2).
Identification of microvesicles and exosomes in conditioned medium. Electron microscopic examination revealed that pellets ultracentrifuged at 20,000 ×g contained round structures measuring 100-800 nm in diameter that were surrounded by a limiting membrane (Figure 3A). Pellets obtained after centrifugation at 100,000 ×g contained a heterogeneous population of round vesicles measuring approximately 100 nm in diameter with a phospholipid bilayer (Figure 3B). These findings are consistent with the characteristics of microvesicles and exosomes, respectively. Vesicles were confirmed to be exosomes by western blotting using antibodies against CD81, a recognized exosomal marker (Figure 3C).

Preparation of microvesicles (MVs), exosomes (EXs), and supernatant (SP). The culture medium was harvested and centrifuged (500 × g, 10 min) to remove cells and large debris. The supernatant was then centrifuged at 20,000 × g for 20 min to obtain pellets containing microvesicles. Exosomes were harvested by centrifuging the supernatant at 100,000 × g for 70 min. The medium after removing cells, cell debris, microvesicles, and exosomes was used as supernatant.
Identification of CD147 in supernatant, microvesicles, and exosomes. Western blotting showed that CD147 was expressed in the supernatant and on microvesicles and exosomes (Figure. 4). The molecular weight (MW) of CD147 on microvesicles and exosomes was approximately 45 kDa (lanes 2 and 6, Figure 4), identical to the MW of CD147 in wild-type A375 cell lysates (lane 1, Figure 4). In contrast, the MW of CD147 in the supernatants was 20 kDa (lane 4, Figure 4), indicating that they contained the cleaved form CD147. The CD147 in all supernatants, microvesicles, and exosomes was effectively silenced by CD147 shRNA (lanes 3, 5 and 7, Figure 4).
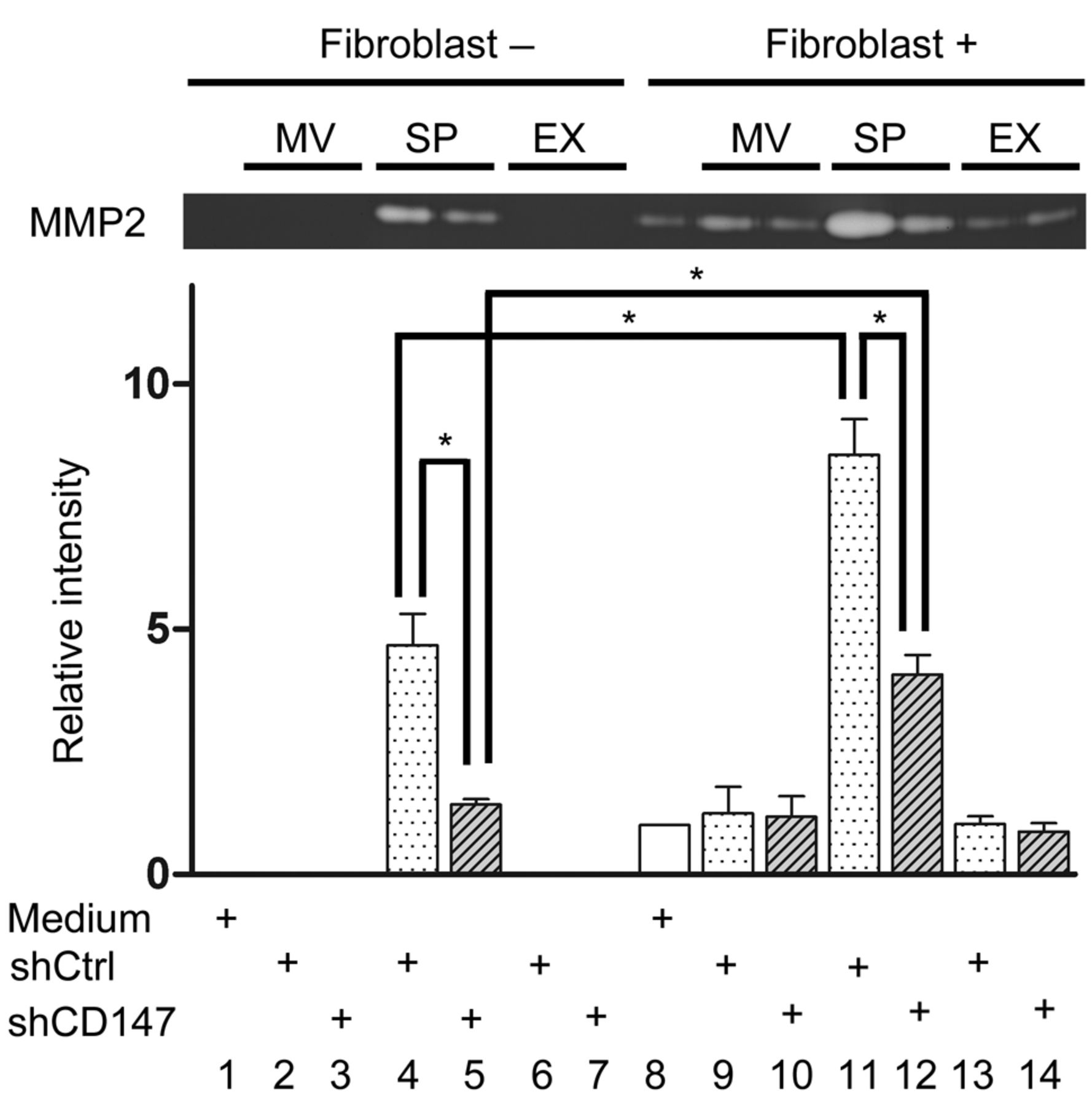
Induction of the gelatinolytic activity of MMP-2 produced by fibroblasts. The stimulatory effect of CD147 in the supernatants, microvesicles, and exosomes on the enzymatic activity of MMP2 was examined by gelatin zymography (Figure 5). The supernatant of A375 cells transfected with CD147 control lentivirus showed MMP2 activity that was inhibited by CD147 silencing (lanes 4 and 5, Figure 5). Microvesicles and exosomes manifested no gelatinolytic activity (lanes 2 and 6 Figure 5). No enzymatic activity was induced when fibroblasts were stimulated by microvesicles and exosomes (lanes 9, 10, 13 and 14, Figure 5). The supernatant of A375 cells transfected with CD147 control lentivirus induced increased enzymatic activity (lane 11, Figure 5); it was significantly lower in A375 cells transfected with CD147 shRNA lentivirus (lane 12, Figure 5). These results demonstrate that the cleaved form of CD147 in the supernatant stimulated fibroblasts to induce MMP2 activity.
Discussion
CD147, originally identified as a cell-surface protein of the immunoglobulin superfamily (1), is highly expressed on the surface of various tumor cells and is involved in tumor progression and metastasis (2, 3). We previously showed that CD147 expressed on MM cells interacts with surrounding fibroblasts to stimulate the production of MMP2, which plays an important role in tumor invasiveness and metastasis (2, 3). However, it remained unclear how CD147 was released from the cell membrane to interact with fibroblasts. Peinado et al. reported that melanoma-derived exosomes contribute to metastatic invasion by carrying messenger proteins that direct bone marrow-derived cells towards a prometastatic phenotype (18). Exosomes and microvesicles are small extracellular membrane vesicles that are thought to serve as extracellular messengers to mediate cell–cell communication. Exosomes are formed within late endocytic compartments called multivesicular bodies and are secreted upon fusion with the plasma membrane. Their diameter ranges from 30-100 nm (19). Microvesicles are derived directly from plasma membrane; they have a diameter ranging from 100-1,000 nm. Millimaggi et al. found that CD147 was expressed in microvesicles derived from epithelial ovarian cancer cells and that CD147-positive vesicles promoted the angiogenic phenotype in endothelial cells (15). Palma et al. reported that CD147 was an enriched surface marker for exosomes in breast cancer cells (20). These findings suggest that CD147 on microvesicles and exosomes plays an important role in tumor–stroma cell interactions. On the other hand, according to Egawa et al., CD147 was cleaved into 22-kDa fragments by membrane type-1 MMP and released into the culture medium of human fibrosarcoma and epidermoid carcinoma cells (16). They proposed that the released 22-kDa fragments stimulate fibroblasts to produce MMPs because they featured the N-terminal Ig-like domain of CD147 that is important for MMP induction.

A: Short hairpin RNA (shRNA)-mediated inhibition of Cluster of differentiation 147 (CD147) protein expression in A375 cells. B: Quantitative comparison of the level of CD147 expression. shCD147: A375 cells transfected with CD147 shRNA lentivirus; shCtrl: A375 cells transfected with control lentivirus. The data shown are representative of three independent experiments.

Electron microscopy reveals the structure of microvesicles and exosomes. Representative electron microscope image of the 20,000 × g pellet fraction (A) and the 100,000 × g pellet fraction (B). Western blotting showed that exosomes express Cluster of differentiation 81 (CD81), a recognized exosomal marker (C). shCD147: A375 cells transfected with CD147 shRNA lentivirus; shCtrl: A375 cells transfected with control lentivirus.
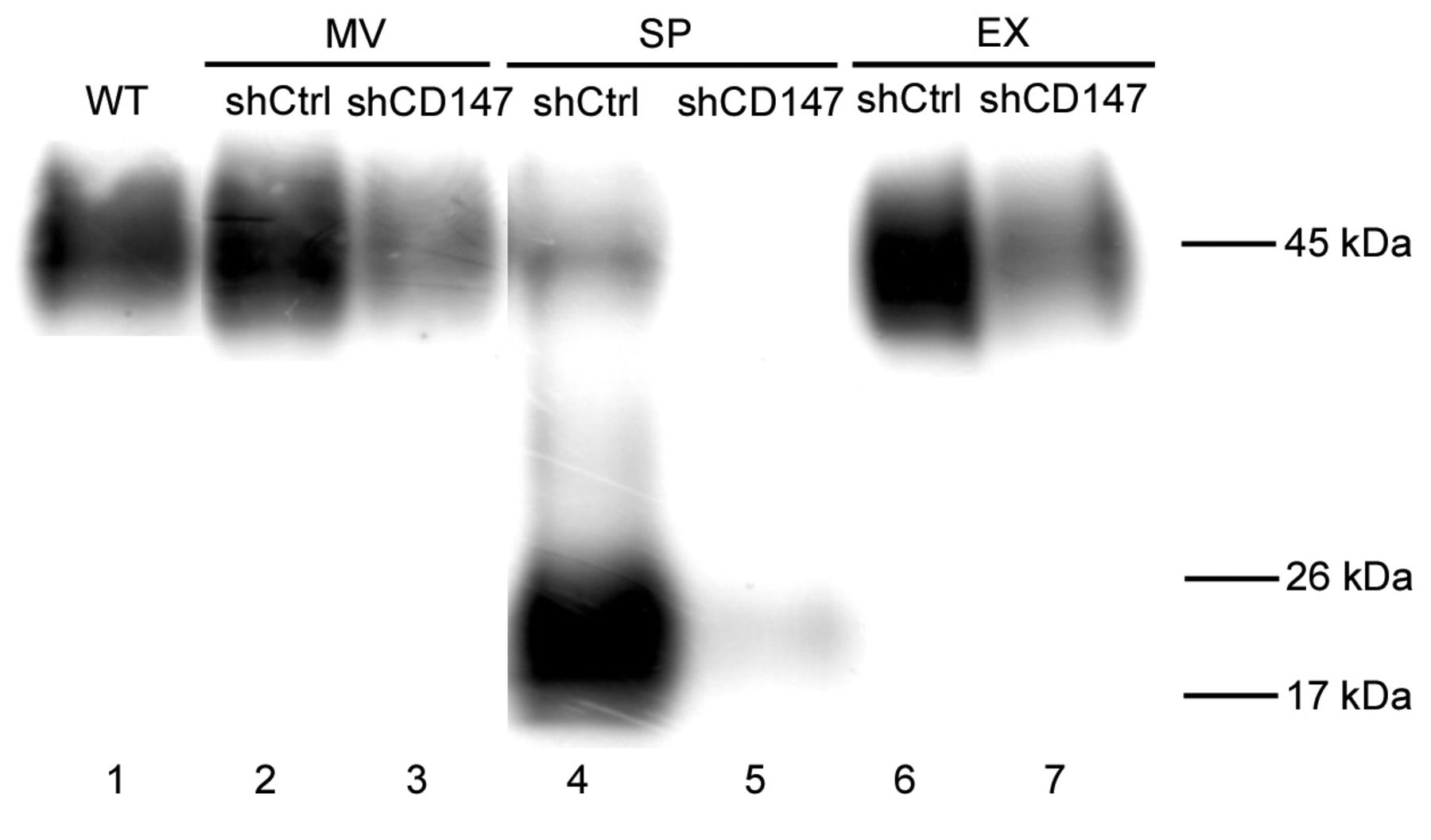
Western blots showing that Cluster of differentiation 147 (CD147) is expressed in microvesicles (MVs), exosomes (EXs), and supernatant (SP) from A375 cells transfected with CD147 shRNA lentivirus (shCD147) or with control lentivirus (shCtrl). Lane 1, Wild-type A375 cells, non-transfected.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The effects of microvesicles (MVs), exosomes (EXs), and supernatant (SP) on the activity of matrix metalloproteinase-2 (MMP2). The top panel is a representative gelatin zymography image. The graph is a quantitative comparison of the MMP2 activity. The data shown are representative of three independent experiments. shCD147: A375 cells transfected with CD147 shRNA lentivirus; shCtrl: A375 cells transfected with control lentivirus. *p<0.05.
We investigated the mechanisms underlying the MMP-inducing activity of CD147 in human MM cells. The supernatant of A375 cells transfected with control lentivirus induced high gelatinolytic activity of MMP2 (Figure 5, lane 4). This enzymatic activity was significantly lower in supernatant from A375 cells transfected with CD147 shRNA lentivirus (lane 5 Figure 5); the MW of CD147 in the supernatant was 20 kDa (lane 4, Figure 4). These results clearly demonstrate that CD147 expressed on cultured MM cells is cleaved and released into the supernatant and that it stimulates fibroblasts to induce MMP2 activity. The antibody used in this study reacts with the distal extracellular region containing the N-terminal Ig-like domain that is important for MMP induction (16).
According to Egawa et al., the MW of cleaved CD147 is 22 kDa (16). CD147 is a highly glycosylated protein with a MW ranging from 43-66 kDa; the MW of its protein portion is approximately 28 kDa (21, 22). As different forms of glycoprotein are produced by different modes of glycosylation depending on the tissue type (1), we attribute the discrepancy in the MW between the present and our earlier findings to different glycosylation modes or differences among tumors with respect to the cleavage sites.
In summary, we demonstrate that the cleaved form of CD147 plays a pivotal role in stimulating fibroblasts to induce MMP2 activity.
Acknowledgements
This study was supported in part by grant no. 21591438 from the Ministry of Education, Science and Culture of Japan.
- Received July 30, 2014.
- Revision received September 12, 2014.
- Accepted September 18, 2014.
- Copyright© 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved