

Abstract
Background: We have previously reported Signal Transducer and Activator of Transcription 3 (STAT3) to be constitutively activated in aldehyde dehydrogenase (ALDH)+/cluster of differentiation-133 (CD133)+ colon cancer-initiating cells. In the present study we tested the efficacy of inhibiting STAT3 signaling in human colon cancer-initiating cells by ursolic acid (UA), which exists widely in fruits and herbs. Results: Our data demonstrated that UA inhibited STAT3 phosphorylation, and induced caspase-3 cleavage of ALDH+/CD133+ colon cancer-initiating cells. UA also reduced cell viability and inhibited tumor sphere formation of colon cancer-initiating cells, more potently than two other natural compounds, resveratrol and capsaicin. UA also inhibited the activation of STAT3 induced by interleukin-6 in DLD-1 colon cancer cells. Furthermore, daily administration of UA suppressed HCT116 tumor growth in mice in vivo. Conclusion: Our results suggest STAT3 to be a target for colon cancer prevention. UA, a dietary agent, might offer an effective approach for colorectal carcinoma prevention by inhibiting persistently activated STAT3 in cancer stem cells.
- Ursolic acid
- STAT3
- colon cancer
- cancer-initiating cells
Human colorectal carcinoma (CRC) is an important contributor to cancer mortality and morbidity. For men and women combined, mortality due to colorectal carcinoma is the second highest cause of cancer-related death in the United States (8). Although many advances have been made in the treatment of colon cancer, the five-year average survival rate of patients considering all stages was only 66% and more than 50,000 patients died of colon cancer in the United States in 2012. According to the American Cancer Society, there will be an estimated 102,480 new cases of colon cancer and 40,340 new cases of rectal cancer, while colorectal cancer will cause 50,830 deaths in the United States in 2013 alone (http://www.cancer.org/cancer/colonandrectumcancer/detailedguide/colorectal-cancer-key-statistics). The evidence for the existence of a very small population of cells called cancer-initiating cells that may be critical in tumor formation and resistance to chemotherapy and radiation is increasing. These cells have the ability to self-renew, driving tumorigenicity, recurrence, and resistance to treatment (19, 22, 24). Thus, it is very important to identify the signal transduction involved in colorectal carcinoma cancer-initiating cells and possible dietary agents which might target these cells for cancer prevention and therapeutic treatment.
The signal transducers and activators of transcription (STAT) protein family mediates cytokine and growth factor-directed transcription (7). STAT3 is persistently activated in many types of human cancer, and activated STAT3 is necessary not only for proliferation, but also for invasion in vitro and in vivo of human colorectal carcinoma (5, 6, 29). More importantly, STAT3 is also necessary for proliferation and survival of colon cancer-initiating cells (17). It, thus, may be a preventive and therapeutic target for human colorectal carcinoma.
Ursolic acid (UA), a triterpenoid, has been identified in apples, pears, and prunes, and appears to be protective against cancer (3). UA may be a potent inhibitor of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), and down-regulates the expression of apoptosis suppressor proteins, such as B-cell lymphoma-2 (BCL2) and BCL-XL in several cancer cell lines, including human colorectal carcinoma (3, 21). UA plays the role of an anticancer agent through several signaling pathways, including the STAT3 pathway (21). However, whether UA can inhibit human colorectal carcinoma cancer-initiating cells has not been reported to our knowledge.
Materials and Methods
Colon cancer cell lines. Human colorectal cancer cell lines (HT29, HCT116, SW480 and DLD-1) were purchased from the American Type Culture Collection (Manassas, VA, USA) and maintained in Dulbecco's modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 4.5 g/l L-glutamine, sodium pyruvate, and 1% penicillin/streptomycin. All cell lines were maintained at 37°C in a humidified incubator with 5% CO2.
Compounds. UA and other natural compounds (resveratrol and capsaicin) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Compounds were dissolved in sterile dimethyl sulfoxide (DMSO) to make 20 mM stock solution, stored at −20°C until use.
Isolation of colon cancer-initiating cells. The ALDEFLUOR kit (StemCell Technologies, Durham, NC, USA) was used to isolate subpopulations with high aldehyde dehydrogenase (ALDH) enzymatic activity as previously described (18). Briefly, cells were trypsinized to single cells using 0.05% trypsin and subsequently suspended in ALDEFLUOR assay buffer containing ALDH substrate (BODIPY®-aminoacetaldehyde, 1 μmol/l per 1×106 cells) and then incubated for 40 min at 37°C. For each sample, an aliquot of cells was stained under identical conditions with 15 mmol/l diethylaminobenzaldehyde (DEAB), an ALDH-specific inhibitor, as a negative control. In all experiments, ALDEFLUOR-stained cells treated with DEAB served as ALDH-negative controls. Antibody to human phycoerythrin (PE)-cluster of differentiation 133 (CD133) was purchased from Miltenyi Biotec (Auburn, CA, USA). ALDH+/CD133+ subpopulations were separated from SW480, HCT116, and HT29 colon cancer cells by a FACS Wantage SE flow cytometer (Becton Dickinion, Palo Alto, CA, USA). After sorting, ALDH+/CD133+ cancer-initiating cells were cultured in serum-free stem cell medium (MEBM; Clonetics Division of Cambrex BioScience, Walkerville, MD, USA) to maintain cancer-initiating cell characteristics. MEBM was supplemented with B27 (Invitrogen, Carlsbad, CA, USA), 20 ng/mL epidermal growth factor (EGF; BD Biosciences, San Jose, CA, USA), 4 μg/ml gentamycin, 1 ng/ml hydrocortisone, 5 μg/ml insulin and 100 μM β-mercaptoethanol (Sigma-Aldrich; St. Louis, MO, USA).
Cell viability assay. Colon cancer-initiating cells (3,000/well in 96-well plates) were incubated with the compounds (0, 5, 10, 25, 50 or 100 μM) in triplicate at 37°C for 72 hours. 3-(4,5-Dimethylthiazolyl)-2,5-diphenyltetrazolium bromide (MTT) viability assay was then performed and the absorbance was read at 595 nm. Half-maximal inhibitory concentrations (IC50) were determined using GraphPad Prism 5 (GraphPad Software, Inc, La Jolla, CA, USA).
Western blot analysis. ALDH+/CD133+ cells separated from HCT116 and SW480 colorectal cancer cells were treated with UA (10 μM or 25 μM) or DMSO for 24 h. DLD-1 colorectal cancer cells were serum-starved in media without FBS for 24 h before being treated with UA (10, 25, or 50 μM) for four hours and incubated with interleukin-6 (IL6, 25 ng/ml) for 30 min. The ALDH+/CD133+ HCT116 and SW480 cells, and DLD-1 colorectal cancer cells were lysed in cold RIPA lysis buffer containing protease inhibitors and subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The proteins were transferred to polyvinylidene difluoride (PVDF) membranes and probed with a 1:1000 dilution of antibodies (Cell Signaling Tech. Beverly, MA, USA) against phosphor-specific STAT3 (tyrosine 705; Y705), phosphor-independent STAT3, cleaved caspase-3, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Membranes were analyzed using Enhanced Chemiluminescence Plus reagents and scanned with a Storm Scanner (Amersham Pharmacia Biotech Inc, Piscataway, NJ).
Tumor sphere culture. The ALDH+/CD133+ subpopulations of HT29, HCT116, and SW480 colorectal cancer cells were plated as single cells in ultra-low attachment six-well plates (Corning, Lowell, MA, USA) at a density of 50,000 viable cells/well in duplicate. Cells were grown in a serum-free stem cell medium as described above in a humidified incubator (5% CO2) at 37°C. On the second day after seeding, the ALDH+/CD133+ cells were treated with DMSO, 10, 25 or 50 μM of UA, resveratrol or capsaicin. Tumor spheres were observed under a microscope 10 to 15 days later. For counting tumor spheres, the content of all wells was collected, pooled, and transferred onto a collagen-coated 6-well dish in differentiating medium (DMEM+10% FBS). Tumor spheres adhered under these conditions in approximately 24 hours, after which they were stained with crystal violet and counted under low magnification.
Mouse xenografts. All animal studies were conducted in accordance with the standard procedures approved by the Institutional Animal Care and Use Committee of Tongji Hospital, Huazhong University of Science and Technology, with the study approved number 00070801. HCT116 colon cancer cells (1×107 in Matrigel) were implanted s.c. into the flank region of 4- to 5-week-old female athymic nude mice (Slac laboratory animal Corporation, Changsha, Hunan province, China). After tumors developed (12 days), the mice were randomized into two groups and treated with 10 mg/kg UA or DMSO (six mice per group) i.p. daily for 13 days. Tumor growth was determined by measuring the major (L) and minor (W) diameters with a caliper. The tumor volume was calculated according to the following formula: tumor volume=0.5236×L×W2.
Results
Inhibition of STAT3 phosphorylation in ALDH+/CD133+ colorectal cancer-initiating cells by UA. To detect the effect of STAT3 phosphorylation in ALDH+/CD133+ colon cancer-initiating cells by UA, we separated ALDH+/CD133+ subpopulations from HCT116 and SW480 human colorectal cancer cell lines by flow cytometry, as previously described and then determined the level of phosphorylated STAT3 by western blots (17). ALDH+/CD133+ subpopulations from HCT116 and SW480 human colorectal cancer cells expressed phosphorylated STAT3 (Y705) when treated with DMSO alone (Figure 1). Such activated STAT3 plays a role in cancer cell proliferation, tumor growth, and reduction of apoptosis (14, 15, 17). We found that 25 μM of UA inhibited phosphorylation of STAT3 at Y705 in ALDH+/CD133+ subpopulations from HCT116 and SW480 human colorectal cancer cells (Figure 1). Additionally, UA selectively inhibited Y705 phosphorylation without affecting the total level of STAT3 (Figure 1).
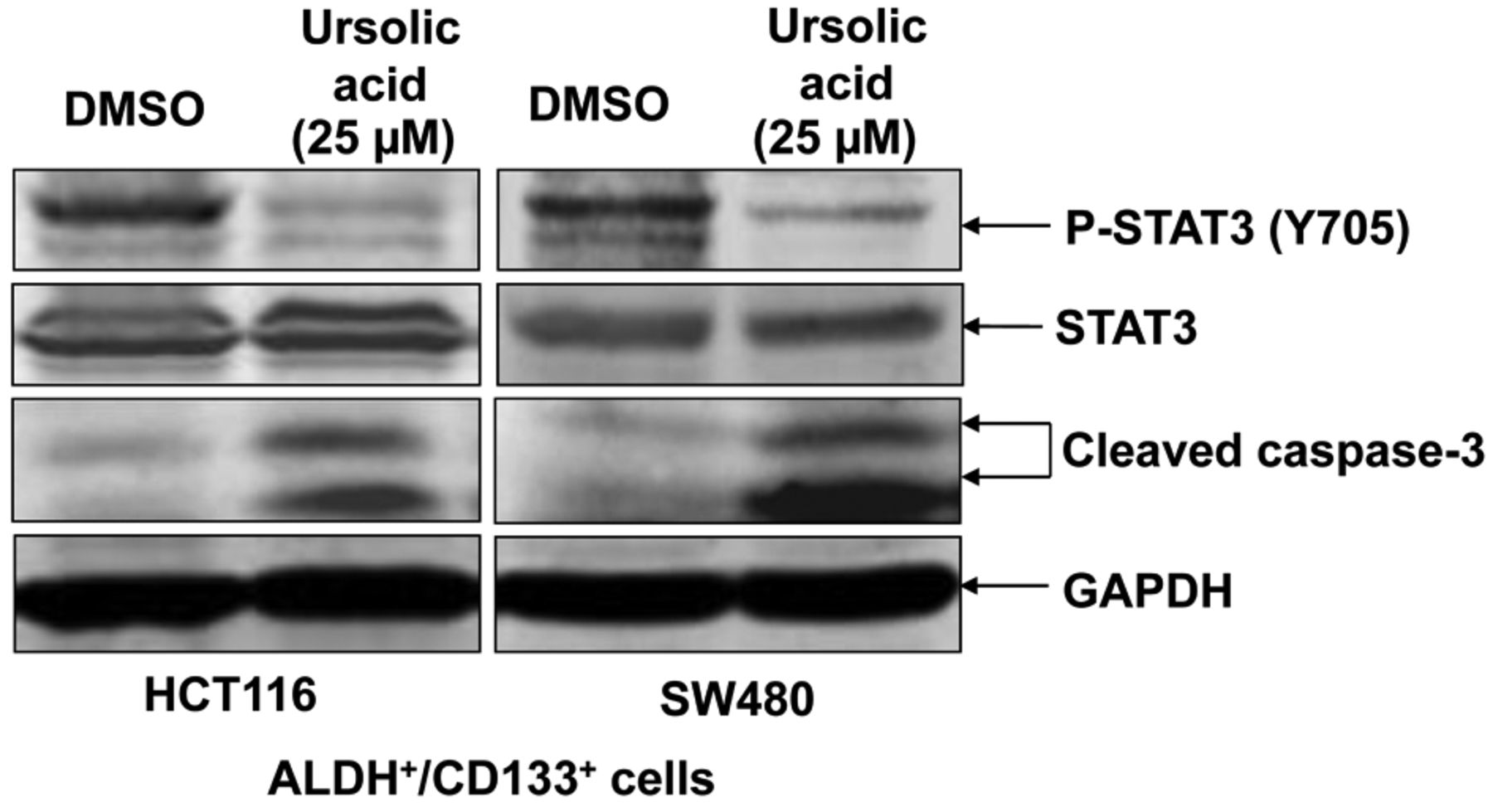
Ursolic acid (UA) inhibited signal transducers and activators of transcription-3 (STAT3) phosphorylation in and induced cleavage of caspase-3 of aldehyde dehydrogenase (ALDH)+/cluster of differentiation 133 (CD133)+ colon cancer-initiating cells. ALDH+/CD133+ cells were separated from HCT116 and SW480 colon cancer cells by flow cytometry and cultured in serum-free stem cell media to maintain cancer-initiating cell characteristics. ALDH+/CD133+ cells were treated with dimethyl sulfoxide (DMSO) or 25 μM UA for 24 h. Phosphorylation of STAT3 (Y705), phospho-independent STAT3, cleaved caspase-3, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were detected by western blot.
Inhibition of IL6-induced STAT3 phosphorylation in colorectal cancer cells by UA. To confirm the effect of UA on STAT3 activation, we then examined its effect on IL6-induced phosphorylation of STAT3 in colorectal cancer cells. DLD-1 colorectal cancer cells were treated with different concentrations of UA prior to incubation with IL6. There was a significant inhibition of phosphorylation of STAT3 in DLD-1 when treated with 10 μM of UA and complete inhibition of phosphorylation of STAT3 when treated with 25 and 50 μM of UA (Figure 2). UA did not affect the total levels of STAT3 (Figure 2).
UA reduced cell viability and induced cleavage of caspase-3 of ALDH+/CD133+ colorectal cancer-initiating cells. We next determined the effect of UA on colorectal cancer-initiating cell viability and apoptosis. Caspase-3 cleavage was induced in ALDH+/CD133+ subpopulations of both HCT116 and SW480 human colorectal cancer cells when treated with UA (Figure 1). The increased expression of cleaved caspase-3 indicates that UA may induce apoptosis in these subpopulations. Our data showed that UA also reduced the cell viability of colon cancer-initiating cells (Figure 3). Cell viability of ALDH+/CD133+ subpopulations of both HCT116 and HT29 was completely abolished by 25 and 50 μM of UA treatment (Figure 3), with a similar effect on SW480 cells (Figure 3). We also investigated the effects of other dietary compounds on colon cancer-initiating cells, namely of resveratrol and capsaicin. The IC50s of resveratrol and capsaicin are higher than UA, which means UA is more potent at inhibiting cell viability of colon cancer-initiating cells (Table I).
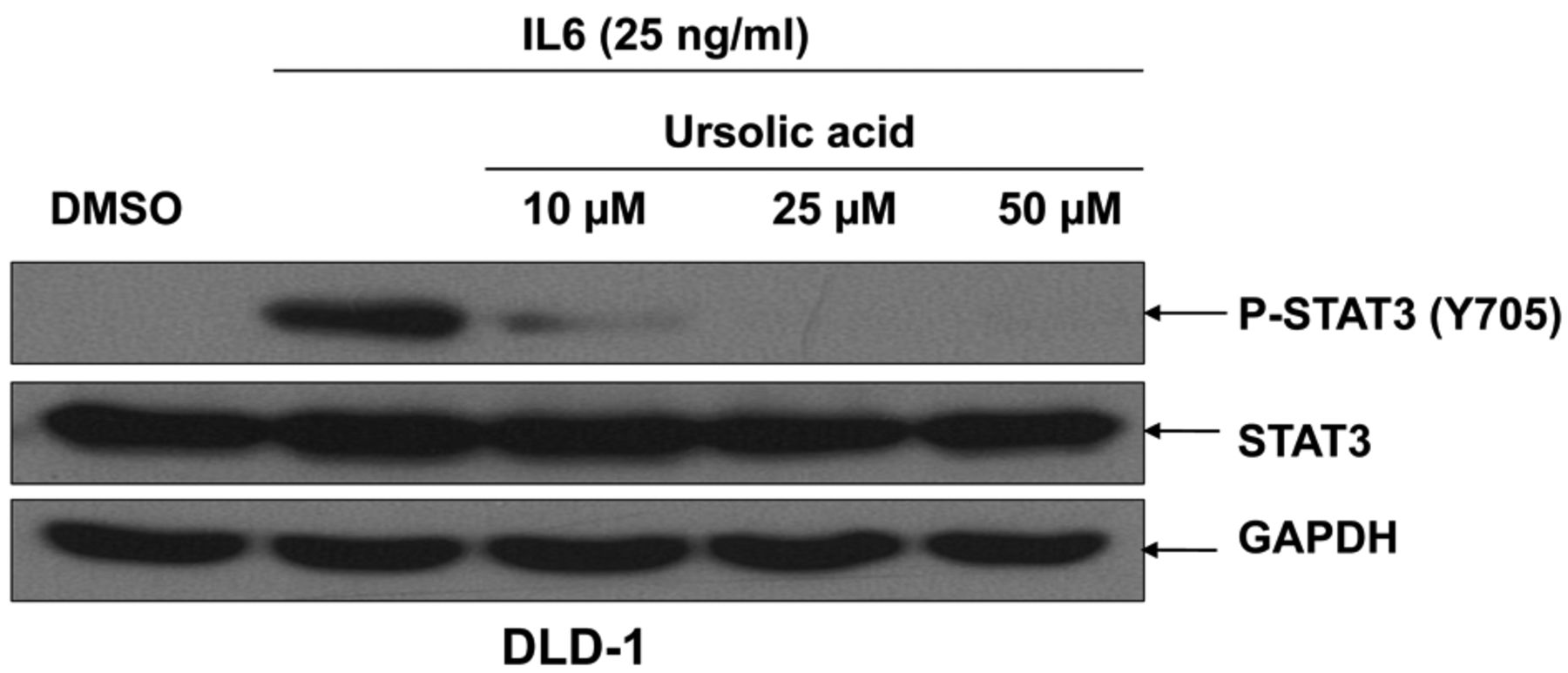
Ursolic acid (UA) inhibited interleukin-6 (IL6)-induced signal transducers and activators of transcription-3 (STAT3) phosphorylation in colon cancer cells. DLD-1 colon cancer cells were cultured for 24 h in Dulbecco's modified Eagle's medium (DMEM) and then serum-starved in DMEM without FBS for 24 h prior to treatment with UA. The cells were treated with the indicated doses of UA for four hours and incubated with IL6 (50 ng/ml) for 30 min. Phosphorylation of STAT3 (Y705), phospho-independent STAT3, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were detected by western blot.
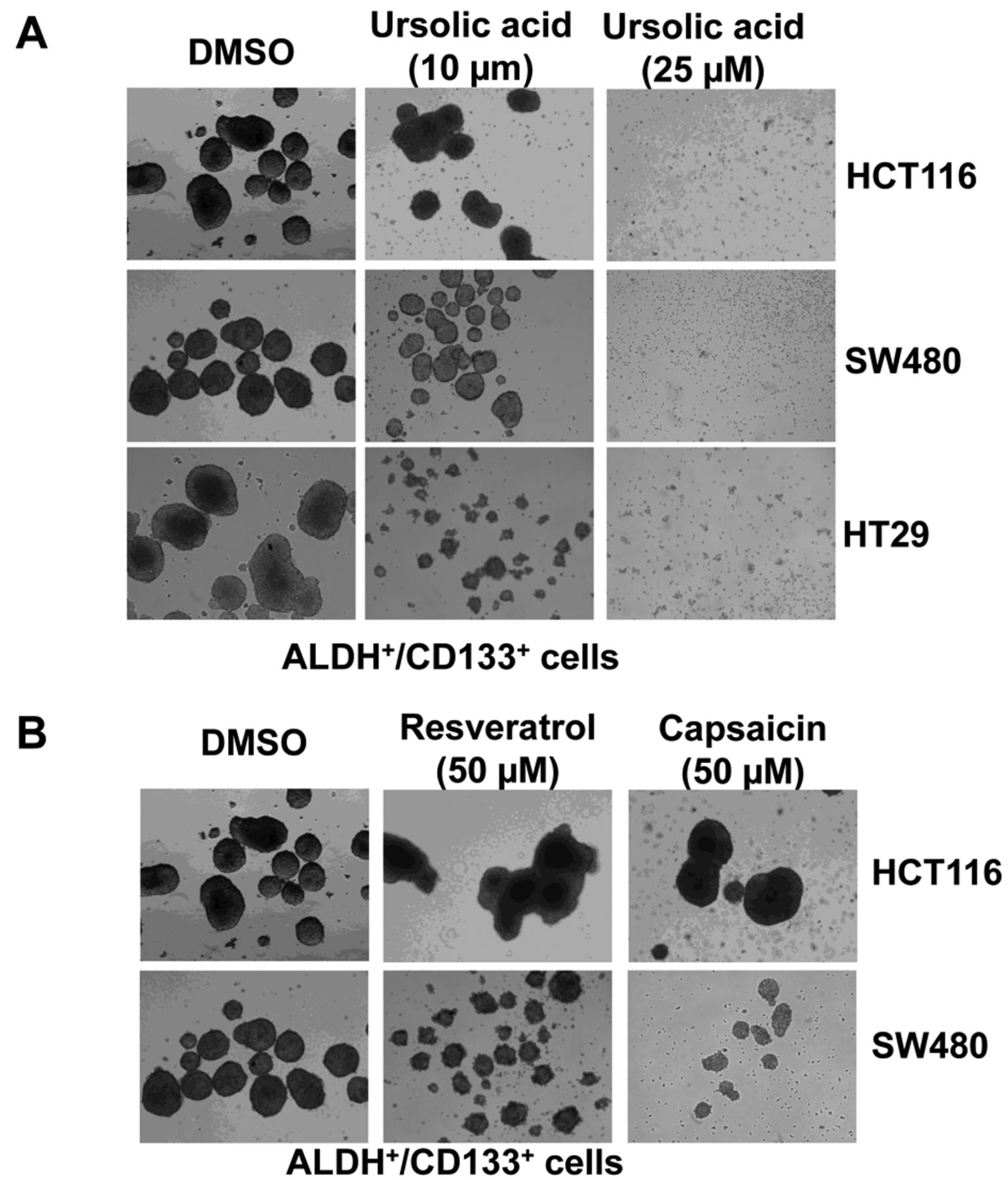
UA inhibited the tumor sphere-forming capacity of ALDH+/CD133+ subpopulations of HCT116, SW480 and HT29 human colorectal cancer cells. Furthermore, we investigated the efficacy of UA at reducing colorectal cancer-initiating cell survival and proliferation in anchorage-independent conditions and on the ability of these cells to form tumor spheres. Our results demonstrate that UA (25 μM) completely inhibited tumor sphere formation capacity in ALDH+/CD133+ subpopulations of HCT116, SW480 and HT29 human colorectal cancer cells (Figure 4A). In contrast to UA, treatment with as high as 50 μM of resveratrol or capsaicin of ALDH+/CD133+ subpopulations of HCT116 and SW480 cells led to decreases in tumor sphere-formation capacity, but not complete inhibition (Figure 4B).
UA suppresses tumor growth in vivo. Mouse xenograft experiments were performed to explore whether UA inhibits tumor growth in vivo. The tumor volume of mice given daily administrations of 10 mg/kg UA exhibited an overall decrease over the course of treatment (Figure 5). In contrast, the tumor volume of mice given daily administrations of DMSO increased over the course of treatment. Daily administration of UA thus resulted in reduced tumor volume in HCT116-xenografted mice compared with their DMSO-treated counterparts (Figure 5).

Ursolic acid (UA) inhibited cell viability of aldehyde dehydrogenase (ALDH)+/cluster of differentiation-133 (CD133)+ colon cancer-initiating cells. ALDH+/CD133+ subpopulations of HCT116, SW480, and HT29 colon cancer cells were seeded in 96-well plates (3,000 cells/well) in triplicate in a serum-free stem cell media. On the following day, ALDH+/CD133+ cells were treated with 5 to 50 μM of UA for 72 h. At the end of this course, 3-(4,5-Dimethylthiazolyl)-2,5-diphenyltetrazolium bromide (MTT) assay was used to determine cell viability. The viability of DMSO-treated cells was set at 100% and compared with the viability of UA-treated cells (*p<0.05).
Discussion
STAT3 is constitutively activated in many types of human cancer, including colorectal cancer (5, 7, 29). STAT3 activation plays a role in invasion in human colorectal carcinoma and is necessary for proliferation and survival of colon cancer-initiating cells (7-11). Colon cancer-initiating cells may play a critical role in tumor formation and resistance to chemotherapy and radiation due to their ability to self-renew, driving tumorigenicity, recurrence, and resistance to treatment (19, 22, 24). Inhibition of the STAT3 signal transduction pathway also inhibits cancer cell growth, which indicates its importance in signal transduction in tumor growth (16, 17). The STAT3 pathway may, thus, serve as a therapeutic target in the treatment of human colorectal cancer through targeting colon cancer-initiating cells.
UA is a triterpenoid compound, widely existing in medicinal herbs and fruits. While UA has been used in health products, it can also inhibit breast (9, 13, 23), lung (11), colon cancer (4, 21, 26), human prostate cancer (12), liver cancer (27), endometrial cancer (1, 2), melanoma (10) and human multiple myeloma (20). UA has been shown to act as an anticancer agent by targeting the STAT3 pathway in prostate cancer cells (13). UA has also been reported to have the capability to inhibit colon cancer cell growth, but whether it can be used to target STAT3 as a strategy to inhibit colon cancer-initiating cell growth has not been determined, to our knowledge.
To explore the effect of UA on the STAT3 signal transduction pathway in colon cancer-initiating cells, we used ALDH and CD133 together, as markers to isolate colon cancer-initiating cells as previously described (17). We first examined the effect of UA on the inhibition of STAT3 in colon cancer-initiating cells. We determined that ALDH+/CD133+ colon cancer-initiating cells expressed phosphorylated STAT3, and UA selectively inhibited STAT3 phosphorylation at Y705 without affecting the total level of STAT3. We then determined that UA treatment reduced cell viability of ALDH+/CD133+ colon cancer-initiating cells. In addition, UA was able to induce cleavage of caspase-3 in ALDH+/CD133+ colon cancer-initiating cells. Our data also show that UA was able to completely inhibit tumor sphere-formation capacity of ALDH+/CD133+ colon cancer-initiating cells. Two other natural and dietary compounds, resveratrol and capsaicin have also been reported to inhibit cancer cell growth. Capsaicin is the active component of chili peppers (28). Resveratrol is a phytoalexin produced naturally by several plants (25). However, the effects of resveratrol and capsaicin on cancer-initiating cells have not been reported. In this study, we found equal concentrations of resveratrol and capsaicin did not exhibit the same degree of inhibition of cell viability and tumor sphere-forming capacity in these cells; the inhibitory effects of UA were more potent. Finally, our data showed that UA was able to reduce tumor volume and growth, while tumor growth and volume increased in control mice. Taken together, our data show that the STAT3 pathway may play an important role in the molecular mechanism of UA-mediated suppression of colon cancer-initiating cells and thus may serve as a therapeutic target for treatment of colorectal cancer.
The half-maximal inhibitory concentration (IC50) of ursolic acid, resveratrol and capsaicin in aldehyde dehydrogenase (ALDH)+/cluster of differentiation 133 (CD133)+ colon cancer-initiating cells.
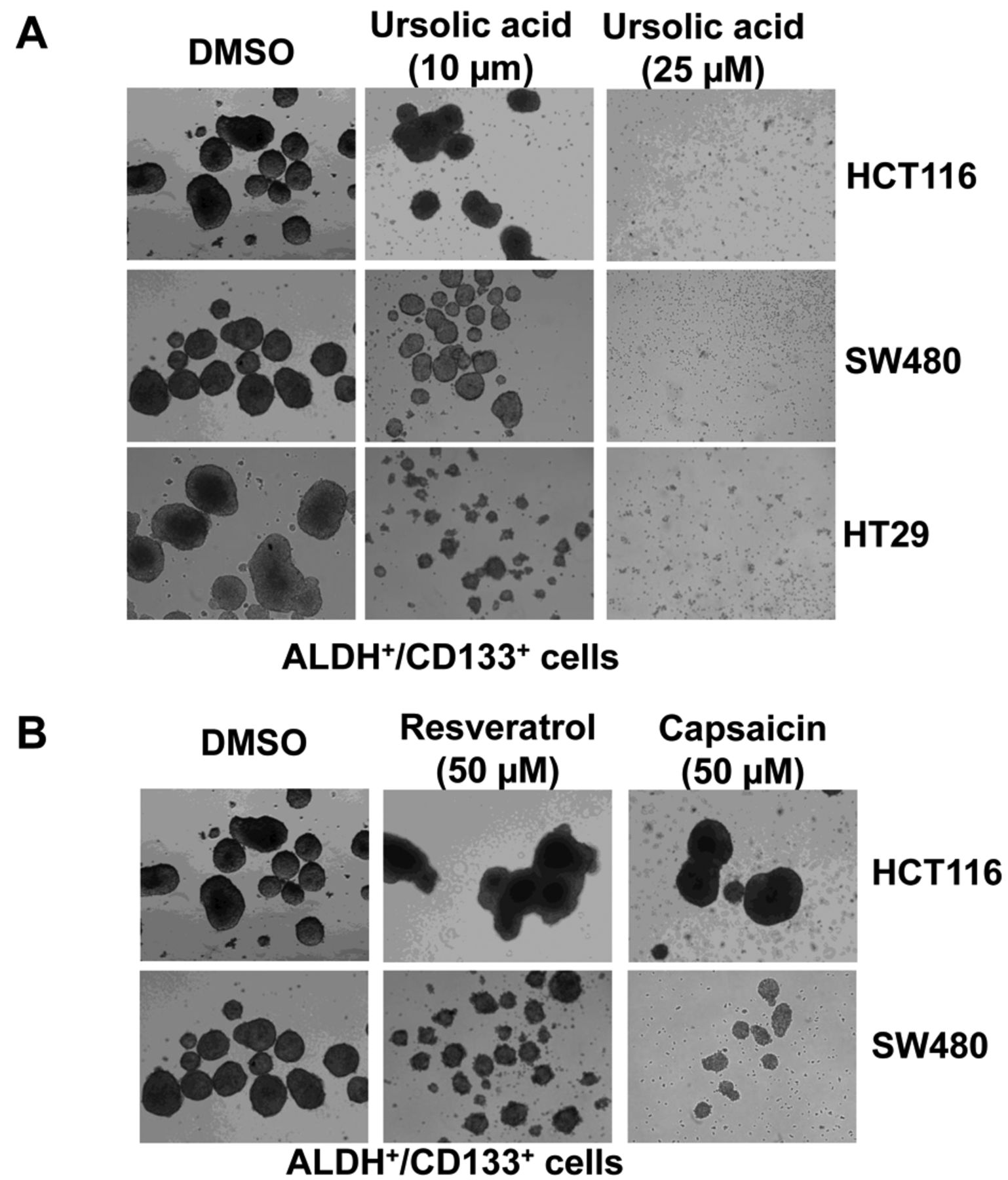
A: Ursolic acid (UA) inhibited tumor sphere formation of aldehyde dehydrogenase (ALDH)+/cluster of differentiation 133 (CD133)+ colon cancer-initiating cells. ALDH+/CD133+ cells were plated as single cells in ultra-low attachment six-well plates (Corning, Lowell, MA, USA) at a density of 50,000 viable cells/well. Cells were grown in a serum-free stem cell medium as described in the Materials and Methods. Twenty-four hours after seeding, the ALDH+/CD133+ cells were treated with 10 or 25 μM of UA. Tumor spheres were observed under a microscope 15 days post-treatment. B: Neither resveratrol nor capsaicin at so inhibited tumor sphere formation of ALDH+/CD133+ colon cancer-initiating cells as potently as UA.
In summary, this study demonstrated that STAT3 is activated in colon cancer-initiating cells and the STAT3 signaling pathway may play important roles in cell viability, tumor sphere-forming capacity, and tumor growth of these cells. UA was able to inhibit STAT3 phosphorylation, tumor sphere-formation capacity and reduce cell viability in colon cancer-initiating cells as well as tumor growth in mice. Since UA is available in a wide variety of foods and has anticancer properties against colon cancer-initiating cells, it may be a potential healthcare product that could be used daily, possibly to prevent colon cancer, as well as being a good drug candidate for the treatment of colon cancer. Eating fruits containing large quantities of UA might be a good way to prevent colon and other types of cancer by targeting cancer-initiating cells.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Ursolic acid (UA) suppressed growth of mouse tumor xenografts of colon cancer cells (*p<0.05). HCT116 colon cancer cells were implanted s.c. into the flank region of 4- to 5-week-old female athymic nude mice. After tumors developed, the mice were given i.p. dosages of 10 mg/kg UA or dimethyl sulfoxide (DMSO) daily over the course of 13 days. Tumor growth was determined by measuring the major (L) and minor (W) diameters with a caliper. Tumor volume was calculated according to the following formula: tumor volume=0.5236×L×W2.
Acknowledgements
This research was supported by National Natural Science Foundation of China (81001005) to Li Lin and the Fundamental Research Funds for the Central Universities, HUST: 0118540019 to Li Lin.
- Received August 1, 2013.
- Revision received September 18, 2013.
- Accepted September 19, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved