

Abstract
Background/Aim: To investigate the effects of inorganic and organic arsenic compounds on human T-lymphoblastoid leukemia cells. Materials and Methods: Cell proliferation was analyzed by 3-(4,5-dimethylthiazol-2-yl)-2,5¬diphenyltetrazolium bromide (MTT) assay. Apoptotic cell morphology was examined by cell staining with Hoechst 33342. Cellular caspase-3/7 activities were measured after arsenic treatment. Results: The inhibitory concentration by 50% (IC50) values of As2O3 towards MOLT-4 and daunorubicin- resistant MOLT-4/DNR cell proliferation were 0.87 and 0.92 μM, while the values for arsenic acid were 69.1 and 116.6 μM, respectively. These arsenic compounds also inhibited mitogen-induced proliferation of human peripheral blood mononuclear cells. Six organic arsenic compounds did not inhibit leukemia cell proliferation. As2O3 and arsenic acid induced apoptotic cell morphology and increased caspase-3/7 activity in the leukemia cells. Ascorbic acid and buthionine sulfoxide enhanced, while N-acetyl-L-cysteine abated, the suppressive effects of inorganic arsenic compounds on leukemia cell proliferation. Conclusion: As2O3 and arsenic acid inhibit proliferation and induce apoptosis in MOLT-4 and daunorubicine-resistant MOLT-4/DNR cells via glutathione-depletion and subsequent caspase-3/7 activation. Organic arsenic compounds have no inhibitory activity on the leukemia cell proliferation. Inorganic arsenic compounds are suggested as useful agents for treatment of T-lymphoblastoid leukemia.
- Arsenic trioxide
- arsenic acid
- organic arsenicals
- T-lymphoblastoid leukemia
- daunorubicin resistance
- apoptosis
As2O3 is an effective and relatively safe drug in the treatment of acute promyleocytic leukemia (1, 2). It has been reported that the effects of As2O3 are not limited to acute promyleocytic leukemia cells but can be observed in various other cell lines of myeloid (3, 4) and lymphoid origin (4), and drug-resistant sublines (5). The action mechanisms of As2O3 in acute promyleocytic leukemia and other malignancies were thought to involved on inhibition of growth and induction of apoptosis (6-8). Glutathione depletion enhances As2O3-induced apoptosis in cancer cells, suggesting that an increase in oxidative stress is related to the antineoplastic efficacies of As2O3 (9). Furthermore, organic arsenic-containing compounds and their selective sensitivity in targeting under division endothelial cells inhibiting angiogenesis, raise the possibility for designing better targeted antineoplastic arsenic-containing compounds with less toxicity to normal cells (10). Although the mechanisms regarding the anticancer actions of As2O3 have been suggested as described above, the inhibitory effects of many other organic and inorganic arsenic compounds on the cancer cell growth have been sparcely studied.
The success of chemotherapy in cancer treatment is frequently limited by intrinsic or acquired multidrug resistance due to increased expression of plasma membrane P-glycoprotein (11). This protein is an ATP-dependent transporter that effluxes a number of structurally unrelated anticancer agents out of the cells, thereby reducing their intracellular drug concentration, permitting cancer cells to survive against high concentrations of drugs that otherwise would be toxic. Inhibiting P-glycoprotein function with diverse pharmacological agents circumvents the multidrug resistance phenotype. MOLT-4, a human T-lymphoblastoid leukemia cell line, has been used extensively for studies of leukemia cell biology and antileukemia therapy (12). We have developed a multidrug resistant MOLT-4/DNR cell line from the MOLT-4 cell line, by exposing the parent cells to increasing stepwise concentrations of daunorubicin over 3 months (13). Multidrug resistance in MOLT-4/DNR has been proved to be closely related to the expression of P-glycoprotein and multidrug resistance 1 (MDR1) mRNA (13), and therefore, this subline should be a suitable model to investigate agents which overcome P-glycoprotein-related drug resistance. We have reported that As2O3 induces apoptosis on parent MOLT-4 cells and on the drug-resistant MOLT-4/DNR cells expressing functional P-glycoprotein via depletion of intracellular glutathione, and subsequent disruption of mitochondrial membrane and activation of caspase-3 (5). However, the effects of organic arsenic compounds and an inorganic arsenic acid on the growth of human T-lymphoblastoid leukemia cells expressing functional P-glycoprotein, or the molecular mechanisms of their action on these cells, have been sparcely investigated.
Thus, the present study was undertaken to evaluate antiproliferative effects of inorganic and organic arsenic compounds on human T-lymphoblastoid leukemia MOLT-4 cells and MOLT-4/DNR cells overexpressing P-glycoprotein, and provides an insight into their action mechanisms by examining cell growth and apoptosis in cells treated with such agents.
Materials and Methods
Reagents. RPMI-1640 medium and FBS were purchased from Gibco BRL Co. (Grand Island, NY, USA). As2O3, sodium arsenate, sodium methylarsonate, tetramethylarsonium, arsenobetaine, dimethylarsinic acid, trimethylarsine oxide, and arsenocholine were purchased from Tri Chemical Laboratories Inc. (Yamanashi, Japan). Chemical structures of the arsenicals used in this study are shown in Figure 1. Stock solutions of these agents were made at a concentration of 10 mM with ethanol or distilled water occasionally using alkali, and diluted to working concentrations by ethanol or distilled water before use. Daunorubicin was obtained from Sigma Chemical Co. (St. Louis, MO, USA). Stock solutions of daunorubicin were prepared at a concentration of 10 mM with distilled water. Ascorbic acid, buthionine sulfoxide, and N-acetyl-L-cysteine were obtained from Sigma-Aldrich. Cell proliferation Kit I and II (MTT) were purchased from Roche Diagnostics Co. (Indianapolis, IN, USA). Concanavalin A was obtained from Biochemistry Biotechnology Co. (Tokyo, Japan). [3H]Thymidine (5.55×1011Bq/mmol) was from GE Healthcare Co. (Tokyo, Japan). Hoechst 33342 solution was a product of Wako Chemical Co. (Tokyo, Japan). Hoechst 33342 solution (1 mg/ml) was diluted with distilled water to 1 mM, and used for the cell apoptosis assay.
Leukemia cell culture and cell proliferation assay. MOLT-4 and MOLT-4/DNR cells were maintained in RPMI-1640 medium containing 10% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin as described previously (5, 12, 13). The leukemia cells were washed and resuspended with the above medium to 5×104 cells/ml, then 98 μl of the cell suspension were placed in each well of a 96-well flat-bottom plate. Two microliters of the solution containing each test compound and 2 μl of daunorubicin-solution were added to yield final concentrations of 0.001, 0.01, 0.1, 1, 10 and 100 μM, respectively. The cells were incubated for 72 h in 5% CO2 at 37°C in a humidified chamber. After the incubation period, 15 μl of the MTT labeling reagent (final concentration 0.5 mg/ml) were added to each well and the plate was mixed on a microshaker, the cells were further incubated for 4 h. Subsequently, 100 μl of solubilization solution were added to each well, and the plate was mixed and allowed to stand overnight in 5% CO2 at 37°C. The spectrophotometric absorbance of the sample was measured on a microplate reader (Corona MT P-32; Corona Co., Japan) at 570 nm. A dose-response curve was plotted for each drug, and a concentration which yielded a 50% inhibition of cell growth (IC50) was calculated. IC50 values were determined from linear regression of at least four points at different concentrations of the agents.
Isolation of human peripheral blood mononuclear cells (PBMCs) and evaluation of arsenical effects in vitro. The present study was approved by the Ethical Committee of Tokyo University of Pharmacy and Life Sciences and written informed consent was obtained from the healthy volunteer included in the study. The study included one healthy female (24 years of age). Evaluation of the effects of arsenic compounds on proliferation of mitogen-activated PBMCs was carried out according to the methods described previously (14). In brief, 20 ml of venous blood were collected from the healthy volunteer. The blood was heparinized and loaded onto 3 ml of Ficoll-Hypaque (Nakarai Co., Japan), centrifuged at 1,300 × g for 20 min and PBMCs were separated as described previously (14). For the evaluation of PBMC sensitivity to the agents, cells were washed and resuspended in RPMI-1640 medium containing 10% fetal calf serum, 100,000 IU/l penicillin and 100 mg/l streptomycin to a final density of 1×106 cells/ml. Concanavalin A, as a T-cell mitogen, was added to each well to a final concentration of 5.0 μg/ml. Subsequently, 4 μl of a solution containing the test compounds were added to give final agent concentrations of 0.1-100 μM. The plate was incubated for 96 h in 5% CO2/air at 37°C. The cells were pulsed with 18.5 kBq/well of [3H]thymidine for the last 16 h of incubation and then collected on glass-fiber filter paper. The radioactivity retained on the filter was further processed for liquid scintillation counting. The mean of the counts for a duplicate of each sample was determined. The experiments were carried out five times, and the mean IC50 values of the test compounds were evaluated.
Cell staining with Hoechst 33342. Three milliliters of the cell suspension of MOLT-4 and MOLT-4/DNR cells at a cell density of 5×104/ml, as prepared above, were incubated in wells of a 6-well plastic plate in the presence of the indicated concentrations of the arsenic compounds for 48 or 72 h in 5% CO2/air at 37°C. After the culture, cells were washed twice in cold phosphate-buffered saline (pH 7.2), then 100 μl of 2% glutaraldehyde was added, and the cells were fixed for 24 h. The cells were then washed with phosphate-buffered saline, resuspended, and 10 μl of this cell suspension was stained with 2 μl of Hoechst 33342. Subsequently, the stained cells were analyzed with fluorescence microscopy.
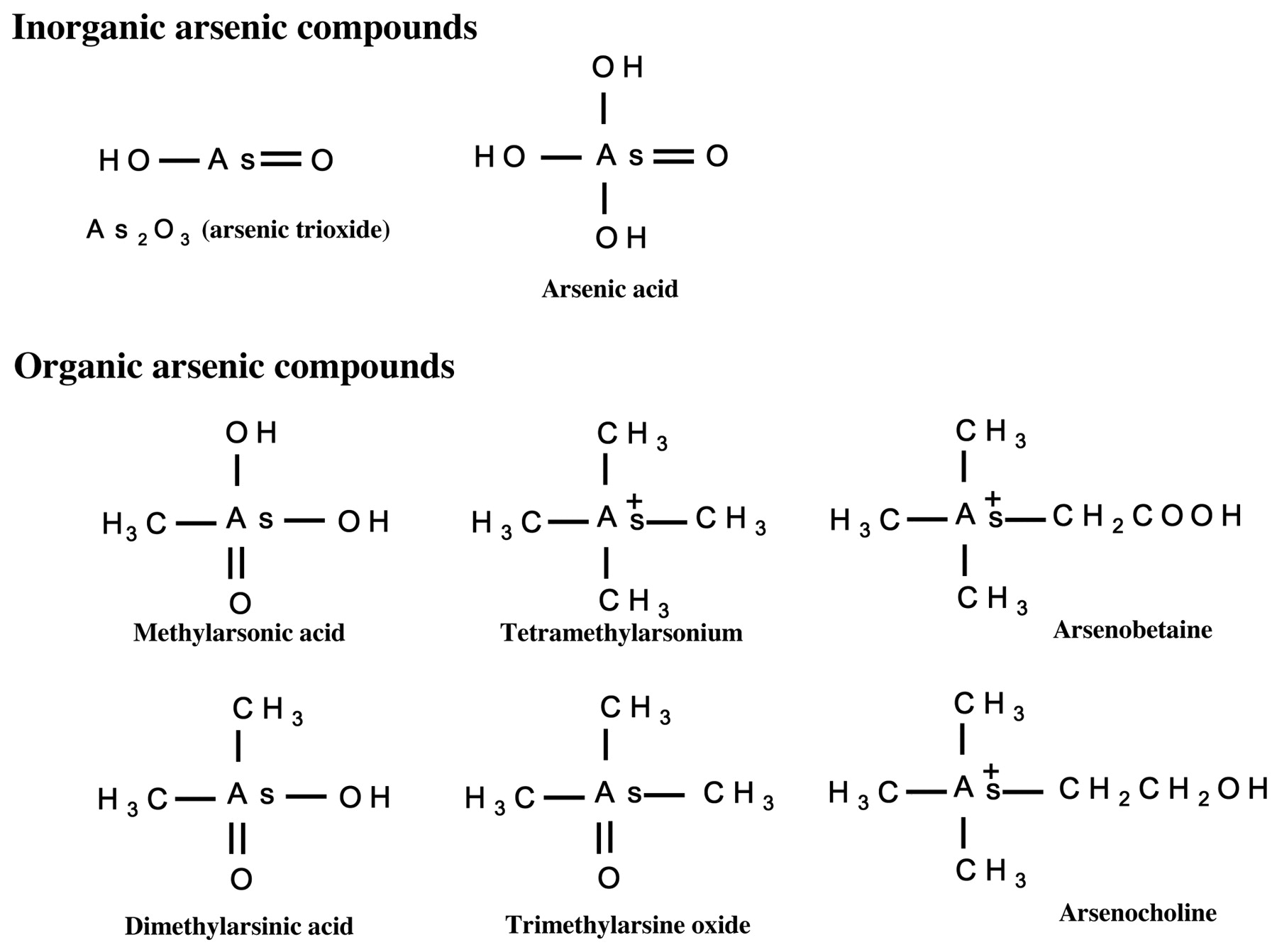
Chemical structures of arsenic compounds examined in this study.
Caspase-3/7 activity assay. Leukemia cells were incubated with the test agents for 3, 6, 12, and 24 h, respectively. Thereafter, 100 μl of Caspase-Glo 3/7 (Promega, Madison, WI, USA) reagent were added to each sample, and the cells were incubated for 30 min and assayed according to the manufacturer's instructions. The data were analyzed with GraphPad Prism 4.0 (San Diego, CA, USA).
Statistics. Leukemia cell proliferation rates in the absence and presence of serial concentrations of the test compounds were compared by Bonferroni test. Additive effects of serial concentrations of organic arsenic compounds with As2O3 were analyzed with Williams' test. Differences in caspase-3/7 activities of leukemia cells treated with As2O3 or arsenic acid, as compared to the enzyme activity in control cells, were analyzed by Dunnett's test. Effects of ascorbic acid, buthionine sulfoxide, and N-acetyl-L-cysteine on the suppressive efficacies of As2O3 and arsenic acid were analyzed by Dunnett's test. In all cases, p-values less than 0.05 were considered to be significant.
Results
Effects of inorganic and organic arsenic compounds on cell proliferation of human leukemia cells and PBMCs. Effects of two inorganic and six organic arsenic compounds on cell proliferation of MOLT-4 cells and MOLT-4/DNR cells were examined in vitro. These cells were incubated in the presence of serial concentrations of the agents for 72 h, and the proliferated cells were analyzed with MTT assay procedures. The IC50 value of daunorubicin for MOLT-4 cell growth was 0.7 μM, while the value for MOLT-4/DNR cell growth was 6.3 μM, i.e., 9 times higher. These observations were consistent with the data of our previous studies (5, 12, 13). Thus, daunorubicin was effective against parental MOLT-4 cells, but less effective against MOLT-4/DNR cells, as expected.
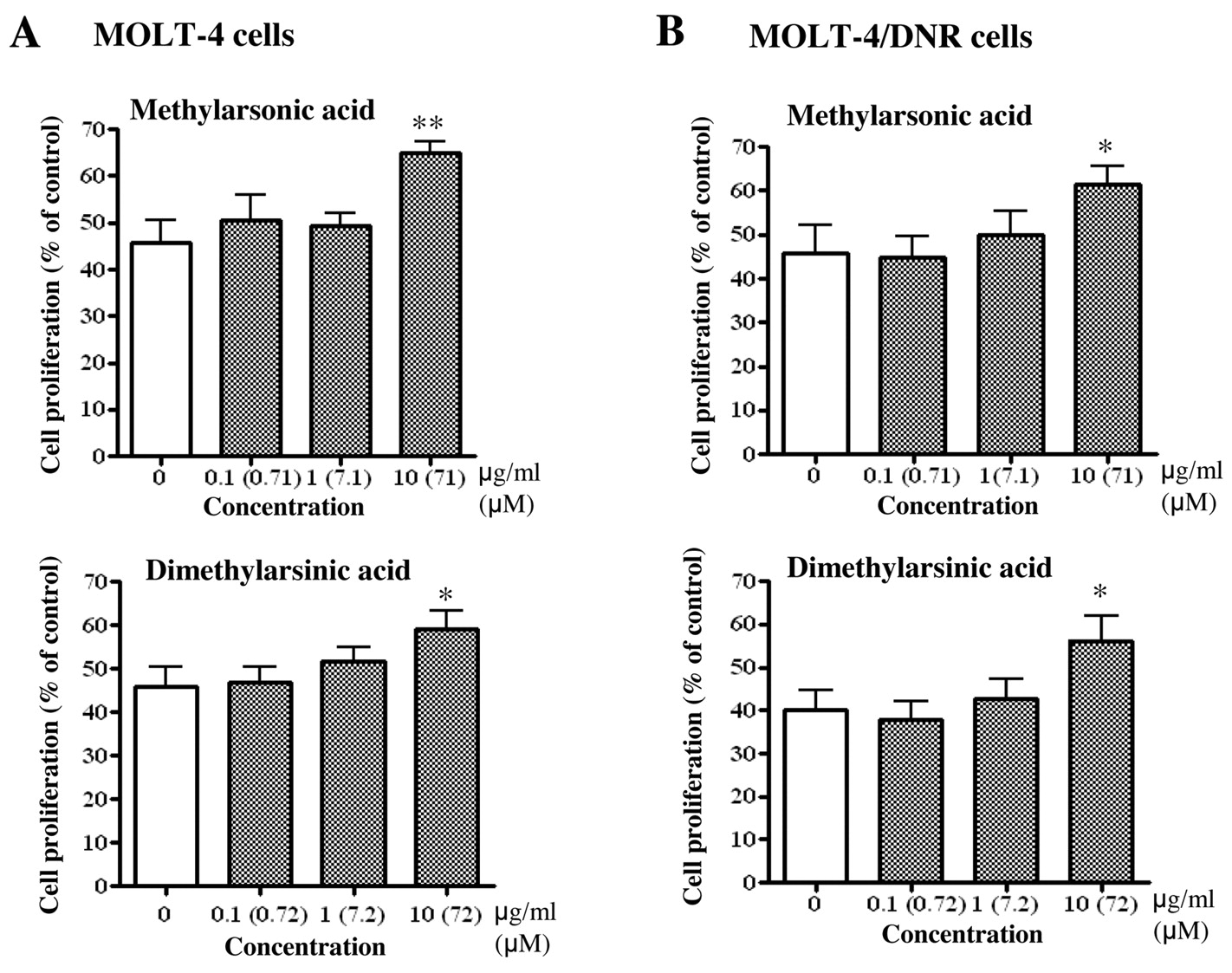
Inorganic arsenic compounds dose-dependently inhibited the proliferation of both MOLT-4 and MOLT-4/DNR cells (Figure 2). The IC50 values of As2O3 for the proliferation of MOLT-4 and MOLT-4/DNR cells were 0.87 and 0.92 μM, while the values for arsenic acid were 69.1 and 116.6 μM, respectively (Table I). The organic compounds methylarsonic acid, tetramethylarsonium, arsenobetaine, dimethylarsinic acid, trimethylarsine oxide, and arsenocholine did not present any inhibitory effects, or rather actually promoted proliferation, at the highest concentrations examined (112-148 μM) (Figure 2). Among these organic compounds, methylarsonic acid and dimethylarsinic acid have been detected in the blood of patients with promyelocytic leukemia, which were administered As2O3 for treatment (15). We then examined the effects of these organic arsenic compounds on the suppressive efficacy of As2O3 against proliferation of the leukemia cells. The suppressive efficacy of 1 μM As2O3 was significantly attenuated by 71 μM methylarsonic acid and 72 μM dimethylarsinic acid (p<0.025) (Figure 3). As2O3 and arsenic acid also inhibited proliferation of human PBMCs stimulated with concanavalin A. The IC50 values of As2O3 and arsenic acid for the concanavalin A-induced PBMC proliferation were 1.9 and 85.7 μM, respectively (Table I). The organic arsenic compounds examined in this study did not affect mitogen-induced proliferation of human PBMCs (Table I).
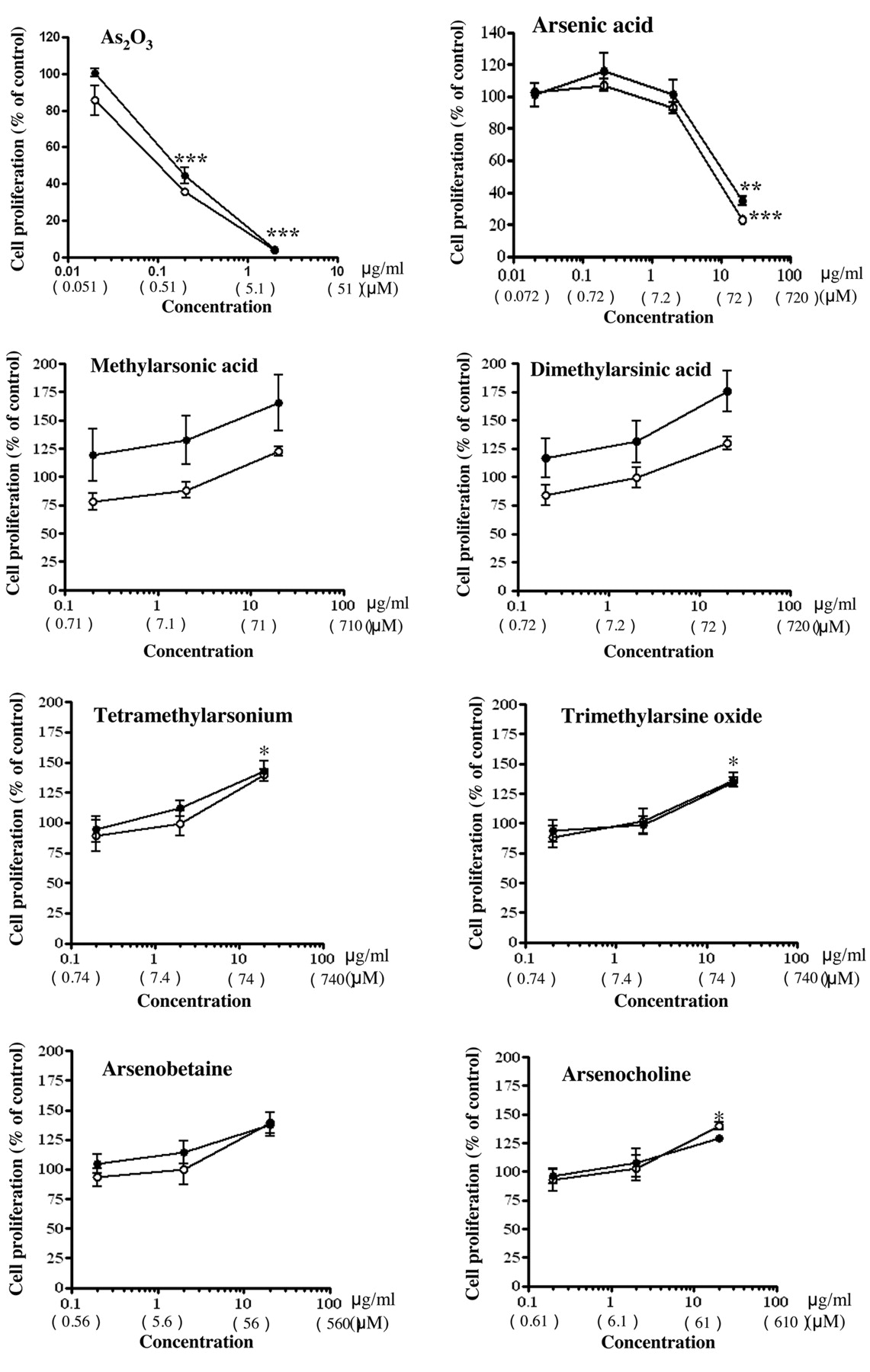
Effects of two inorganic and six organic arsenic compounds on in vitro proliferation of MOLT-4 and MOLT-4/DNR cells. Cell proliferation was examined by MTT assay, and the percentages of cell proliferation as compared to control were plotted. Values are the means of three independent experiments. Statistical significance was determined by Bonferroni test. *p<0.05, **p<0.01 and ***p<0.001 compared with each control.

Attenuation of growth inhibitory effect of 1 μM (0.2 μg/ml) As2O3 by organic arsenic compounds, methylarsonic acid and dimethylarsenic acid, on MOLT-4 (A) and MOLT-4/DNR (B) cell culture. Leukemia cells were cultured for 72 h in the presence of 1 μM As2O3 without or with serial concentrations (0.1-10 μg/ml, i.e., 0.71-72 μM) of methylarsinic acid (upper figures) or dimethylarsonic acid (lower figures), and the proliferated cells were analyzed by MTT assay. Values are the means of three independent experiments. Statistical significance was determined by the Williams' test. *p<0.025 and **p<0.005 compared with control.
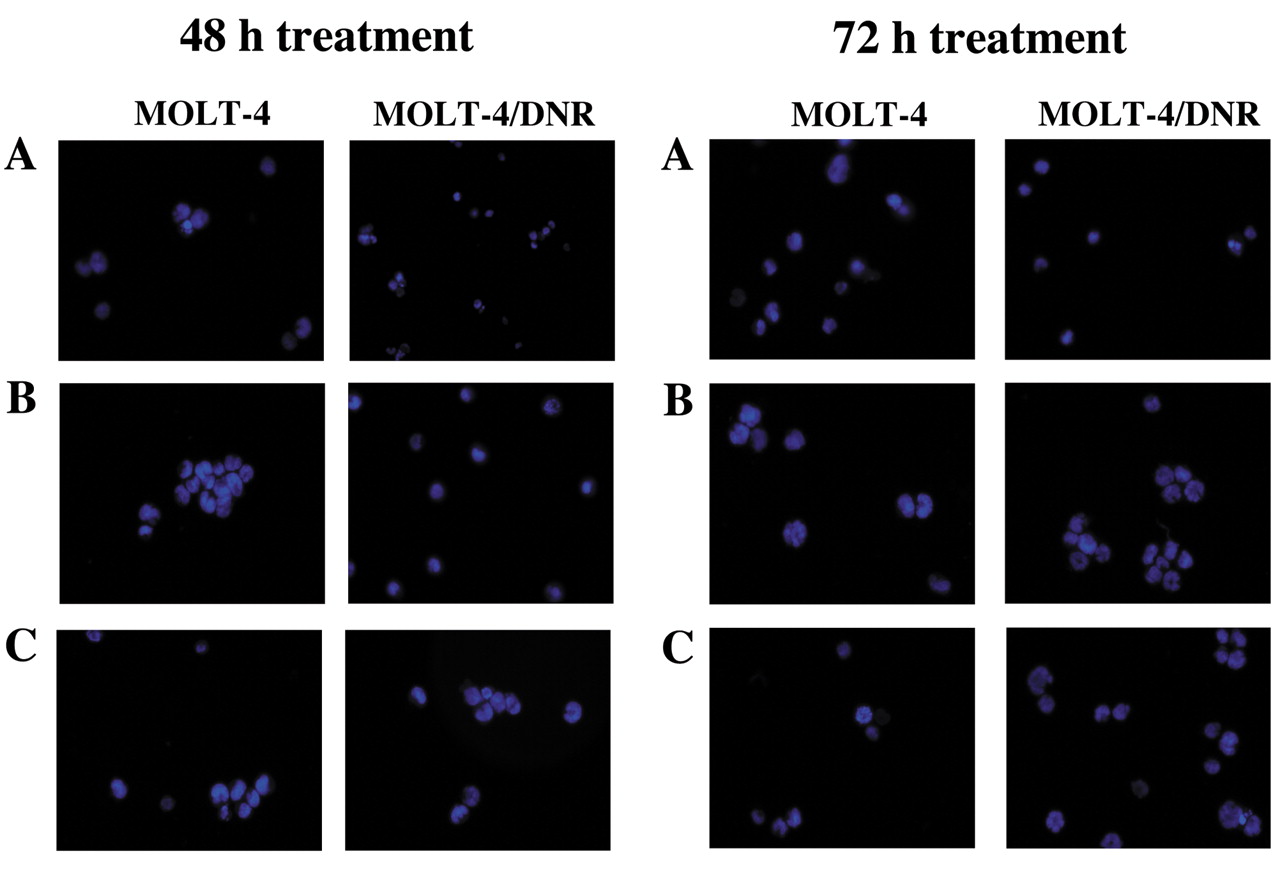
Apoptotic morphology and caspase-3/7 activity in leukemia cells treated with inorganic and organic arsenic compounds. MOLT-4 and MOLT-4/DNR cells were incubated in the presence of the arsenic compounds for 48 or 72 h, and the cells were stained with Hoechst 33342, followed by analysis with fluorescence microscopy (Figure 4). Treatment of cells with As2O3 at 1 μM (Figure 4B) or arsenic acid at 72 μM (Figure 4C) for 48 and 72 h induced apoptotic cell morphology such as atypical chromatin condensation and multinucleation, in both MOLT-4 and MOLT-4/DNR cells. Case pase-3/7 activities in MOLT-4 and MOLT-4/DNR cells treated with As2O3 or arsenic acid for 3, 6, 12, and 24 h are shown in Figure 5. Case pase-3/7 activity in MOLT-4 cells treated with 1 μM As2O3 for 6 h significantly increased (p<0.05), as did that of the cells treated with 72 μM arsenic acid for 12 and 24 h, as shown in Figure 5A (p<0.01). Similarly, the enzyme activities in MOLT-4/DNR cells treated with As2O3 or arsenic acid for 12 h increased significantly as shown in Figure 5B (p<0.01 and p<0.05, respectively).

Induction by As2O3 and arsenic acid of atypical chromatin condensation and multinucleation in MOLT-4 and MOLT-4/DNR cells, determined by staining with the fluorescent dye Hoechst 33342 at 48 h (left) and 72 h (right) after treatment. A: Untreated cells; B: As2O3-treated cells; C: Arsenic acid-treated cells.
Modulation of antiproliferative effects of As2O3 and arsenic acid by ascorbic acid, buthionine sulfoxide, or N-acetylcysteine. Antiproliferative effects of As2O3 and arsenic acid in the presence of ascorbic acid, buthionine sulfoxide, and N-acetylcysteine were examined, and the data are presented in Figure 6. In these experiments, the cells were treated with 1 μM As2O3 or 72 μM arsenic acid in combination with 37.5 μM ascorbic acid, 6 μM buthionine sulfoxide, or 4 mM N- acetylcysteine, respectively. Antiproliferative effects of 1 μM As2O3 on both MOLT-4 and MOLT-4/DNR cells were enhanced by ascorbic acid and buthionine sulfoxide, and the effects of ascorbic acid and buthionine sulfoxide in the MOLT-4/DNR cells were statistically significant as compared to the cells treated with 1 μM As2O3 alone (p<0.001 and p<0.01, respectively) (Figure 6B). Similarly, the antiproliferative effects of 72 μM arsenic acid on both MOLT-4 and MOLT-4/DNR cells were enhanced by ascorbic acid and buthionine sulfoxide (p<0.05-0.001) (Figures 6A and B). In contrast, N- acetylcysteine significantly attenuated the antiproliferative efficacies of both As2O3 and arsenic acid in MOLT-4 and MOLT-4/DNR cells (p<0.01-0.001) (Figure 6A and B).
Cells treated with either of 37.5 μM ascorbic acid, 6 μM buthionine sulfoxide, or 4 mM N-acetylcysteine alone did not show any significant changes in the percentage of MOLT-4 and MOLT-4/DNR cells as compared to the control cells (Figure 6A and B, left).
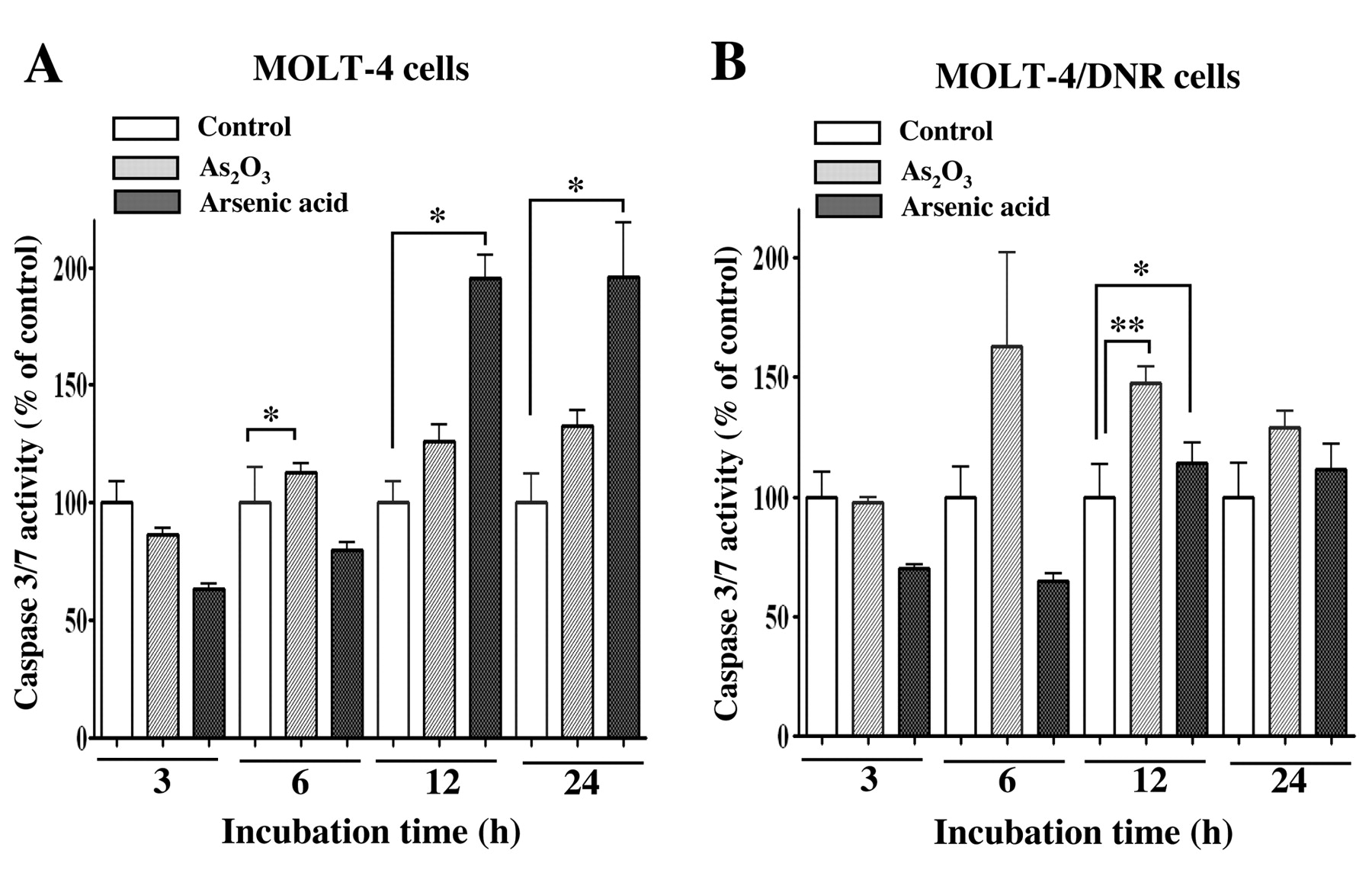
Changes in caspase-3/7 activity of MOLT-4 (A) and MOLT-4/DNR (B) cells after treatment with As2O3 or arsenic acid. Values are the means ±SD of three independent experiments. Statistical significance was determined by Dunnett's test. *p<0.05 and **p<0.01 compared with each control.
Discussion
Our previous study revealed that As2O3 induces apoptosis in cells of human T-lymphoblastoid leukemia MOLT-4 cell line and daunorubicin-resistant MOLT-4/DNR cell line via depletion of intracellular glutathione, and subsequent disruption of mitochondrial membrane potential and activation of caspase-3 (5). In contrast, the effects of arsenate and organic arsenic compounds on the growth of these cells, or the molecular mechanisms of their action on these cells, have been sparcely investigated. Thus, the present study was undertaken to evaluate antiproliferative effects of inorganic and organic arsenic compounds systematically on MOLT-4 and MOLT-4/DNR cells, and to provide an insight into their action mechanisms. The data provided above show that As2O3 and arsenic acid inhibit proliferation of both MOLT-4 cells and P-glycoprotein-expressing daunorubicin-resistant MOLT-4/DNR cells. The data also suggested that the inorganic arsenic compounds inhibit cell proliferation and induce apoptosis in both MOLT-4 and MOLT-4/DNR cells to almost the same extent, possibly via depletion of intracellular glutathione and subsequent activation of caspase-3/7. However, the six organic arsenic compounds examined in this study did not inhibit, but in fact rather promoted proliferation of MOLT-4 and MOLT-4/DNR cells.
Inhibitory concentration by 50% (IC50) values (μM) for the arsenic compounds examined in this study against the in vitro proliferation of MOLT-4 and drug-resistant MOLT-4/DNR cell lines, and peripheral blood mononuclear cells (PBMCs).
MOLT-4/DNR cell line was derived from parental MOLT-4 cells by exposure to a stepwise concentration of daunorubicin in our laboratory (13), and these cells were shown to be persistently resistant to the antiproliferative effect of daunorubicin. The IC50 value of daunorubicin towards MOLT-4/DNR cells was 9 times higher than that towards MOLT-4 cells. The resistance of MOLT-4/DNR cells to daunorubicin has been reported to correlate closely with the expression of functional P-glycoprotein (13). Multidrug resistance is recognized as one of the most common causes of failure for chemotherapy in treating cancer patients (11). P-Glycoprotein is an ATP-binding cassette transporter which hydrolyses ATP and extrudes cytotoxic drugs from mammalian cells (11). As described above, we revealed that both parental MOLT-4 and drug-resistant MOLT-4/DNR cells were equally sensitive to the suppressive effects of As2O3 and arsenic acid, suggesting that these inorganic arsenic compounds are not extruded by P-glycoprotein. In contrast to the effects of the inorganic arsenic compounds, the organic arsenic compounds examined here did not show any inhibitory effects on the in vitro proliferation of MOLT-4 cells, MOLT-4/DNR cells, and human PBMCs. Little is known about the effects of organic arsenic compounds on cancer cell proliferation to the best of our knowledge, and thus the present data would be the first report for such effects of such compounds. In general, organic arsenic compounds have less toxicity as compared to their inorganic counterparts, such as As2O3. Furthermore, we found a unique action of organic compounds including methylarsonic acid and dimethylarsinic acid, where these compounds are suggested to attenuate the inhibitory efficacies of As2O3 on T-lymphoblastoid leukemia cells. Methylarsenic acid and dimethylarsenic acid are found in the blood of patients with acute promyelocytic leukemia, after administration of As2O3 for treatment (15). Although the concentrations of these organic arsenic compounds detected in patients' blood (15) were less than those we used in this study, the influence of these compounds on the outcome of the As2O3 therapy should be examined in clinical cases in future studies.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Modulation of As2O3 and arsenic acid-mediated inhibition of MOLT-4 (A) and MOLT-4/DNR (B) cell proliferation by ascorbic acid, buthionine sulfoxide, and N-acetylcysteine. Cell proliferation was examined by MTT assay, and the percentages of cell proliferation as compared to controls were plotted. Values are the means±SD of three independent experiments. Statistical significance was determined by Dunnett's test. *p<0.05, **p<0.01 and ***p<0.001 compared with As2O3 or arsenic acid alone.
Our data showed that both As2O3 and arsenic acid inhibited proliferation of human PBMCs stimulated with a T-cell mitogen concanavalin A, suggesting that these inorganic compounds have immunosuppressive effects. The IC50 levels for these compounds towards mitogen-activated PBMCs are similar to those of these compounds for the proliferation of MOLT-4 and MOLT-4/DNR cells (Table I). These observations suggest that long-term clinical use of As2O3 should be carried out with careful consideration for possible occurrence of infectious complications in patients with acute promyelocytic leukemia.
As2O3 induced apoptotic cell morphology such as atypical chromatin condensation and multinucleation in both MOLT-4 and MOLT-4/DNR cells. These observations are consistent with our previous findings that this compound induces apoptosis in these cells (5). Here we also showed that arsenic acid, as well as As2O3, causes apoptotic cell morphology in MOLT-4 and MOLT-4/DNR cells. In addition, As2O3 and arsenic acid were shown to activate caspase-3/7 in the cells, which supports our conclusion that these inorganic compounds induce apoptosis in MOLT-4 and MOLT-4/DNR cells. Mitochondria are known to play a major role in apoptosis triggered by many stimuli (16). In the first stage of apoptosis, signal transduction cascades or damage pathways are activated. Subsequently, mitochondrial membrane function is lost, and then proteins released from mitochondria cause the activation of catabolic proteases and nucleases (17). Caspases are cysteine proteases that mediate apoptosis by proteolysis of specific substrates, and caspase-3 is considered a primary executioner of apoptosis (18).
In the present study, we showed that antiproliferative effects of As2O3 and arsenic acid in both MOLT-4 and MOLT-4/DNR cells were enhanced by ascorbic acid and buthionine sulfoxide, whereas the effects were attenuated by N-acetylcysteine. Thus the suppressive effects of As2O3 and arsenic acid can be potentiated by ascorbic acid, which reduces glutathione-dependent dehydroascorbate, and by buthionine sulfoxide, which inhibits synthesis of glutathione. In contrast, the effects of As2O3 and arsenic acid were attenuated by N-acetylcysteine, which promotes synthesis of glutathione, in both parental MOLT-4 cells and daunorubicin-resistant subline cells. The effects of As2O3 are consistent with those we have observed in our previous study (5), but the effects of arsenate on human T-lymphoblastoid cells are reported for the first time. As2O3 sensitivity of cancer cells has been reported to correlate with intracellular glutathione levels (19). Glutathione contents modulate the growth-inhibitory and apoptosis-inducing effects of arsenic compounds (19, 20). Cells expressing higher levels of glutathione or glutathione-associated enzymes are known to be less sensitive to As2O3 than cells expressing lower levels of these molecules (20). Moreover, cells with increased glutathione levels can be sensitized to As2O3 by agents that deplete intracellular glutathione (21). The decrease of glutathione level in cells acts as a potent early activator of apoptosis signaling (22). Depletion of glutathione is believed to induce the loss of mitochondrial membrane potential (23). Mitochondrial permeability transition resulting from intracellular thiol depletion is known to be a critical event in apoptosis (24, 25). Our previous study showed that As2O3 depletes intracellular glutathione in both MOLT-4 and MOLT-4/DNR cells (5). Thus, the present data, taken together with our previous observations, suggest that As2O3 and arsenic acid induce apoptosis via depletion of cellular glutathione and following activation of caspase-3/7.
In summary, the present study showed that inorganic arsenic compounds As2O3 and arsenic acid exhibit suppressive effects on the in vitro growth of both human T-lymphoblastoid leukemia MOLT-4 cells and P-glycoprotein-expressing daunorubicin-resistant MOLT-4/DNR cells. The data also suggest that As2O3 and arsenic acid inhibit cell proliferation and induce apoptosis in these cells via depletion of intracellular glutathione and subsequent activation of caspase-3/7. On the other hand, six organic arsenic compounds examined in this study did not inhibit, but rather promoted proliferation for both MOLT-4 and MOLT-4/DNR cells.
- Received September 8, 2011.
- Revision received October 25, 2011.
- Accepted October 27, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved