

Abstract
Background: Although zoledronic acid (ZOL) has been reported to show anti-tumour effects on various malignant tumours, few studies have reported the molecular mechanisms of resistance to ZOL. Materials and Methods: A drug-resistant cell line was developed by repeatedly treating human osteosarcoma cell line HOS with ZOL. Expression status of drug resistance-related molecules including ATP-binding cassette (ABC) and heat-shock protein 27 (HSP27) was confirmed in order to analyse molecular mechanisms of the drug resistance. Results: Repeated treatment with ZOL induced drug resistance with down-regulation of apoptosis in the resistant cell line. Although ABC expression was down-regulated, up-regulation of HSP27 in the resistant cell line was confirmed. The resistance was overcome by HSP27 silencing. Conclusion: Resistance to ZOL in osteosarcoma cells may be induced by a molecular mechanism different from conventional efflux pump-based resistance. Up-regulation of HSP27 expression may play a role in the development of ZOL resistance in HOS cells.
- Osteosarcoma
- drug resistance
- zoledronic acid
- heat shock protein 27
Bisphosphonates (BPs) are stable synthetic analogs of pyrophosphate whose clinical use has markedly increased in the past decade. Different side chains can be added to the central carbon atom to produce a range of BPs with varying clinical activity and potency. Osteoporosis is the most common indicator for the use of these compounds, but they are being increasingly used in the treatment of osteolytic bone diseases characterized by enhanced bone resorption such as Paget's disease. Recently, BPs have also been investigated as a treatment option for the hypercalcemia caused by malignancy and the bone pain caused by focal tumour-induced osteolysis (1, 2). Zoledronic acid (ZOL) is a new-generation nitrogen-containing BP and is one of the most potent inhibitors of bone resorption among the currently available BPs. Recent clinical studies have shown that, in addition to their potent antiosteoclastic effects, ZOL and other BPs induce apoptosis in cancer cells of several origins, including myeloma, breast cancer, prostate carcinoma, chondrosarcoma and osteosarcoma cell lines (3-5).
Osteosarcoma is the most common primary malignant bone tumour in children and adolescents. The first choice treatment for osteosarcoma is chemotherapy with multiple anticancer drugs such as doxorubicin, cisplatin, high-dose methotrexate and ifosfamide, which are used in combination with wide resection (6). In the last three decades, the use of neoadjuvant chemotherapy has increased the long-term survival rate of osteosarcoma patients from 10-20% to approximately 80% (6). However, patients who do not respond to these conventional chemotherapies have a poor prognosis. Therefore, new and alternative approaches to treatment for osteosarcomas are required. In this regard, several studies have reported the anti-osteosarcoma effects of BPs in vitro (7-9) and in vivo (7, 10), thereby suggesting that BPs are promising alternatives for osteosarcoma treatment.
Analysis of the molecular mechanisms underlying the resistance of cancer cells to chemotherapy is essential for overcoming drug resistance in tumour cells. The phenomenon whereby tumour cells develop resistance to many unrelated drugs is known as multidrug resistance (MDR). The possible mechanisms underlying resistance of cancer cells include overexpression of efflux pumps, inhibition of apoptosis, increased repair of DNA damage, mutations in key cell cycle checkpoint genes, and increased or altered drug targets (11). While osteosarcomas have been reported to exhibit an MDR phenotype, the clinical significance of the mechanism underlying this MDR is a matter of debate (12-14).
Despite the importance of BPs in the clinical management of cancer, few studies have reported the molecular mechanism underlying the development ZOL resistance in cancer cells (15,16); these studies have suggested that in addition to the conventional efflux pump mechanism, different types of molecular mechanisms are involved in the development of drug resistance in tumour cells. In this study, a ZOL-resistant osteosarcoma cell line was developed. It was found that tumour cells that were repeatedly treated with sub-lethal concentrations of ZOL developed marked resistance to the drug. Results of a cytotoxicity assay performed using a conventional chemotherapeutic agent doxorubicin combined with the result of expression analysis of the representative ATP-binding cassettes (ABC) suggested that the molecular mechanism underlying the resistance of the ZOL-treated tumour cells is different from the conventional drug resistance mechanism in osteosarcoma. The ZOL resistance appeared to be mediated by up-regulation of heat-shock protein 27 (HSP27), an anti-apoptotic molecule, in the resistant cells. Conversely, HSP27 silencing in the tumour cells reduced the ZOL resistance, thereby suggesting a close relation between HSP27 and ZOL resistance.
Materials and Methods
Cell culture and growth profile analyses. Human osteosarcoma cell line HOS was obtained from the American Type Culture Collection (17). The HOS cells were cultured at 37°C in Eagle's minimum essential medium (E MEM) supplemented with 10% fetal bovine serum (FBS) in a humidified atmosphere containing 5% CO2.
Cell proliferation assay. Cell proliferation was monitored as described in a previous study, with some modifications (18). The cells were seeded into 96-well plates at a density of 3000 cells/well. After 24 h incubation, the cells were treated with media containing the indicated reagents. After 48 or 96 h incubation, the viable cells were detected using a spectrophotometric CellTiter 96 aqueous cell proliferation assay (Promega, Madison, WI, USA).
Development of drug resistance. ZOL was kindly provided as the hydrated disodium salt (mol. wt. 401.6) by Novartis International AG (Basel, Switzerland). A ZOL-resistant cell line HOSR was developed from the parent cell line by exposing the cells to sub-lethal concentrations of ZOL (30-50 μM) for 96 h followed by incubating them in a complete medium lacking the drug for 10 days. ZOL-resistant cells were obtained after repeating this treatment protocol 10 times with increasing concentrations of ZOL.
Growth curve profile. The cells (1×104/well) were seeded into 6-well plates, and cell proliferation was evaluated at 1-7 days after culturing by cell counting in low power field using a photomicroscope.
Detection of apoptosis. The cells were seeded into 96-well plates at a density of 3000 cells/well and incubated for 24 h. After washing with phosphate-buffered saline (PBS) twice, medium containing the indicated reagents was added to the wells. After 48 h incubation, apoptosis was detected using ApoStrand™ ELISA Apoptosis Detection Kit (Enzo Life Sciences International, Inc., Plymouth Meeting, PA, USA).
Preparation of total RNA. Total RNA was extracted from the HOS and HOSR cells using RNeasy mini kit and RNase-free DNase Set (Qiagen, Valencia, CA, USA). The purity of the RNA thus extracted was assessed by measuring absorbance at 260 and 280 nm (A260/A280 ratios between 1.9 and 2.1 were considered acceptable). RNA concentrations were determined on the basis of the absorbance values at 260 nm.
Real-time reverse transcription-polymerase chain reaction. Quantitative real-time reverse transcription-polymerase chain reaction (RT-PCR) analysis was performed using Applied Biosystems 7500 real-time PCR system using TaqMan gene expression assays system (Applied Biosystems, Foster City, CA, USA). The TaqMan probes and primers for HSP27 (assay identification number Hs00356629_g1), ABC, sub-family C, member 1 (also known as multidrug resistance-associated protein 1 (MRP1)) (assay identification number Hs00219905_m1), ABC sub-family G member 2 (ABCG2) (assay identification number Hs00184979_m1), and ABC, sub-family B, member 1 (ABCB1) (also known as multidrug resistance protein 1 (MDR1)) (assay identification number Hs00184500_m1) were Assay-On-Demand gene expression products (Applied Biosystems). Human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene was used as an endogenous control (Applied Biosystems).
Immunoblotting analysis. Cell lysates (10 μg of protein/sample) were resolved using 5% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS–PAGE) and transferred to a polyvinylidene difluoride (PVDF) membrane. Immunoblotting was performed using an affinity purified mouse monoclonal antibody against HSP27 (Cell SignalingTechnology, Danvers, MA, USA). The blot was incubated with horseradish peroxidase (HRP)-conjugated immunoglobulin G (IgG) and HRP-linked secondary antibody (Cell Signaling Technology), and fluorescence detection was performed using the LumiGLO peroxidase chemiluminescent substrate kit (KPL, Gaithersburg, MD, USA).
RNA interference. RNA interference (RNAi) of HSP27 expression was induced using SignalSilence® HSP27 siRNA kit (Cell Signaling Technoogy). Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) was used as the transfection agent. The working concentration of small interfering (si) RNA in the experiments was 10 nM.
Statistical analysis. All the data in the present study are expressed in terms of mean±SD. Intergroup differences were analyzed using analysis of variance (ANOVA) and Dunnett's test.
Results
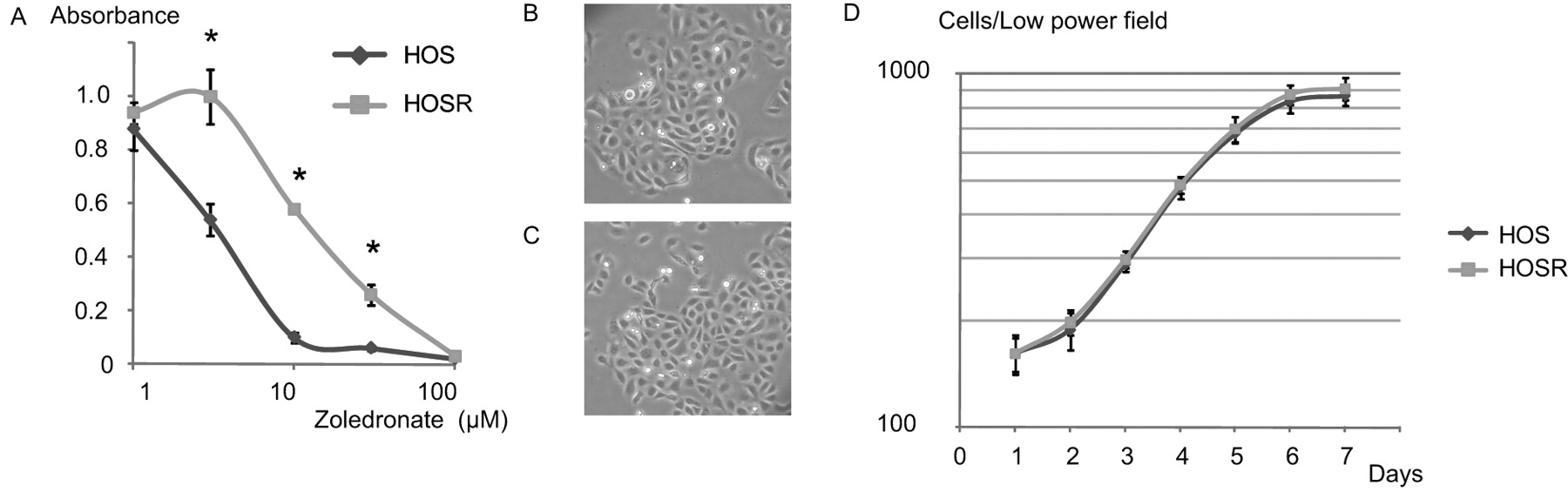
Development of drug resistance. To investigate drug resistance in HOS cells, a variant HOS cell line, HOSR, was developed that showed resistance to the cytotoxic effects of ZOL. After exposing the parent cell line to increasing concentrations of ZOL over several months, a subpopulation of cells was obtained that was able to withstand the effects of the drug at higher concentrations than the parent cell line. The ZOL responses of the parent HOS and HOSR cells in a cytotoxicity assay are shown in Figure 1A. The 50% inhibitory concentrations (IC50) were 3.3 and 13.2 μM in HOS and HOSR, respectively, showing that the variant cell line was approximately 4.0-fold more resistant to the cytotoxic effects of ZOL.

Development of cell line showing resistance to zoledronic acid (ZOL). A: ZOL sensitivity of the parent (HOS) and resistant (HOSR) cells. Both parent and resistant cells were exposed to the indicated concentrations of ZOL for 96 h, and the viable cells were qualified by a spectrophotometric assay. B and C: Morphologies of the parent HOS (B) and resistant HOSR (C) cells (×100). D: Growth curves of the two cell lines. Known numbers of single cells were plated into 6-well culture dishes. Cell numbers were counted daily by using a photomicroscope. The data points are the means±SD of 8 samples. *p<0.05, vs. control.
Morphological comparison of the two cell lines on the basis of microscopic examinations revealed no evident differences between the two cell populations (Figures 1B and C). In addition, growth curve analysis revealed no marked differences in the growth profiles of the two cell lines (Figure 1D).
Cross-resistance and expression of drug transporters. To characterize the drug resistance properties of the HOSR cell line against other drugs, the cross-resistance of this cell line to alendronate and doxorubicin was confirmed. The HOSR cells showed cross-resistance to alendronate, a representative BP (Figure 2A) (IC50: 4 and 13 μM in HOS and HOSR cell line, respectively). However, HOSR cells were more sensitive to doxorubicin, which is a representative clinical chemotherapeutic agent against osteosarcoma (Figure 2B), thereby suggesting that the HOSR cells respond differently at the biochemical level. Because the doxorubicin resistance of osteosarcoma was reported to be associated with members of the ABC superfamily (12, 14), the expression status of three representative members of ABC superfamily was confirmed by using real-time PCR. Real-time PCR did not reveal up-regulation of the expression of these genes in the resistant cells (Figure 2C), which supports the hypothesis.
Detection of apoptosis. ZOL-induced growth inhibition of osteosarcoma cells has been reported to be mediated through apoptosis (7, 9, 19), therefore the apoptosis induction status in the two cell lines was assessed. Significant inhibition of apoptosis induction in the resistant cell line was obtained (Figure 3A).
RNAi and confirmation of the role of HSP27 in the survival of the resistant cells. These findings suggest that the drug resistance of the HOSR cells was, at least partly, mediated by inhibition of apoptosis induction. HSP27 has been reported to participate in protection of cells from stress-induced apoptosis including the apoptosis induced by ZOL treatment (20, 21). Thus, it was hypothesised that HSP27 may be involved in the development of drug resistance in this cell line. The protein (Figure 3B) and RNA (Figure 3C) expression levels of HSP27 in the parents and resistant cell lines were investigated, and a marked up-regulation of HSP27 at both protein and RNA levels was detected. Next, the RNAi-induced expression of HSP27 was investigated and a marked down-regulation of HSP27 expression at both protein (Figure 3D) and RNA (Figure 3E) levels was detected. However, no marked down-regulation of the RNAi-induced HSP27 expression in the HOS cells was detected, probably because of the low baseline expression of HSP27 in this cell line. On the basis of these findings, the role of HSP27 in the drug resistance of the HOSR cells was confirmed. RNAi-induced silencing of HSP27 mRNA eliminated the drug resistance at a ZOL dose of 10-30 μM (Figure 3F). Considering the close association between drug resistance and ZOL-induced apoptosis and the previously-reported role of HSP27 in the prevention of apoptosis, it was hypothesized that up-regulation of HSP27 could lead to prevention of apoptosis and enhanced cell survival in the resistant cells. Thus the role of HSP27 in the prevention of apoptosis in the HOSR cells was confirmed. The rate of apoptosis in the HOSR cells pretreated with HSP27 siRNA was significantly greater than that in the cells treated with non-specific control siRNA (Figure 3G).

Analysis of drug resistance of HOSR cells. A: Alendronate sensitivity of the parent and resistant cells, indicating the cross-resistance of HOSR cells to the anti-tumour effect of alendronate. B: Cell proliferation assay using doxorubicin revealed that HOSR cells showed greater sensitivity to doxorubicin than HOS cells did, suggesting that the mechanism of bisphosphonate resistance differs from conventional ATP-binding cassette (ABC) transporter-based drug resistance. Real-time reverse transcription-polymerase chain reaction (RT-PCR) was performed to analyse RNA expression of the members of the ABC superfamily. C: Baseline expression levels of each gene in the parent cell line. The assays were performed in triplicate and representative data are shown here. *p<0.05, vs. control.
Discussion
Recent studies have shown that ZOL directly affects the proliferation and survival of osteosarcoma cells in vitro (7-9, 19, 22, 23). Although the molecular mechanism underlying ZOL activity is not fully understood, these studies suggest the effect of ZOL was at least partly mediated by cell cycle arrest and induction of apoptosis. These findings evoked interest regarding the in vivo biological effects of this agent. In fact, several studies performed in orthotopic mouse or rat osteosarcoma models showed that ZOL inhibited the growth of osteosarcoma cells in vivo (10, 24-26). Furthermore, the rate of incidence of complications for ZOL treatment is less than that for conventional chemotherapy for osteosarcoma. This finding, combined with the previously mentioned data, suggests that ZOL has potential clinical application for the treatment of osteosarcoma. However, prolonged contact of cancer cells with the drug will increase the chances of the development of drug resistance (27, 28). Since BP is selectively localized and retained in bone tumour (29), the possibility of drug resistance development is high.
Although a variety of molecular mechanisms for MDR have been reported (11), few studies have reported the molecular mechanism underlying the resistance to BPs. In one such study, a ZOL-resistant breast cancer cell line developed by continuous treatment of the cell with sub-lethal concentrations of ZOL (100 nM-8 μM) for 1 year showed up-regulation of ABC transporters including breast cancer resistance protein (BCRP) and lung resistance protein (LRP) (15). In addition, this model revealed up-regulation of the anti-apoptotic molecule BCL-2. Another drug-resistant osteosarcoma cell line showed up-regulation of farnesyl diphosphate synthase, which is an enzyme involved in the mevalonate pathway and is vital for the prenylation of small GTPases (16). Interestingly in this model, the cells were markedly susceptible to conventional anticancer agents, which is consistent with the current results. In the resistance model utilized in this study, sensitivity to doxorubicin was up-regulated with down-regulation of ABC transporters expression. Although the underlying reason for the difference in molecular mechanisms is unknown, the Authors believe that such variations can be attributed to the heterogeneity of the tumour cells and the differences in the cell lines and the methods used for inducing resistance. Consequently, the molecular mechanism of the development of ZOL resistance remains to be elucidated.
HSP27, a member of the small heat-shock protein family, is expressed in various types of cells in response to stress including heat shock, oxidative stress, hypertonic stress, and other forms of cellular injury (30). The molecule protects cells against stress by inhibiting apoptosis (20). Apoptosis, characterized by the condensation of nuclear chromatin, cytoplasmic shrinkage, membrane blebbing, nuclear fragmentation and formation of apoptotic bodies, is induced by a large variety of stimuli including cytokines, cytotoxic drugs, oxidative stress and ionizing radiation (31), some of which are also key phenomena in the induction of HSP27 expression. HSP27 inhibits apoptosis via both Fas ligand pathway and mitochondrial pathway (30). In addition to inhibiting the caspase activation pathway, it also promotes cell survival under stress by activating molecular chaperones, preventing cytoskeleton disruption, and modulating intracellular redox potential (30). The inhibition of HSP27 expression in osteosarcoma cells by anti-sense HSP27 was reported to reduce the rate of cell survival in vitro, thereby suggesting the important role of this molecule in the survival of osteosarcoma cells (32). Moreover, HSP27 overexpression is a poor prognostic factor in osteosarcoma patients (33), indicating that this molecule may enhance the survival of osteosarcoma cells in vivo.

{kind=link}
{kind=link}
{kind=link}
Analysis of the molecular mechanism underlying drug resistance. A: Detection of apoptosis induced by the indicated concentrations of zoledronic acid (ZOL) in the parent and resistant cell lines. Cells were treated with the indicated concentrations of ZOL for 48 h. Monoclonal antibody against single-stranded DNA was used to detect apoptosis. The intensities indicating the apoptosis rate were normalized using the intensity for the parent cell line. B and C: Differences between the heat-shock protein 27 (HSP27) expression levels in parent and resistant cell lines. Immunoblot analysis (B) and real-time reverse transcription-polymerase chain reaction (RT-PCR) (C) revealed up-regulation of HSP27 expression in the resistant cell line. D: Effect of RNA interference (RNAi) on HSP27 mRNA was confirmed by immunoblot assay. Lane 1, protein from cells that did not undergo small interfering (si)RNA treatment. Lane 2, protein from cells treated with specific siRNA for HSP27. Lane 3, Protein from cells treated with non-specific (NS) siRNA. E: Effect of siRNA on HSP27 expression was confirmed by using real-time RT-PCR. F: Cell proliferation of each cell line with silenced HSP27. Cells were pre-treated with specific or non-specific control siRNA and then the indicated concentration of ZOL was added. After incubation for 48 h, the viable cells were detected using a spectrophotometric cell proliferation assay. G: Detection of the inhibitory effect of HSP27 silencing on ZOL-induced apoptosis in the resistant cell line. Only specific siRNA for HSP27 facilitated ZOL-induced apoptosis. *p<0.05, vs. control.
Many studies have reported the close relationship between HSP27 and tumour cell susceptibility to conventional chemotherapeutic reagents. For example, in the herceptin-resistant model of breast cancer cells, the up-regulation of HSP27 promoted cell survival by stabilizing the epidermal growth factor receptor (EGFR) kinase Her2-related tyrosine, a herceptin target (34). In colon cancer cell lines, HSP27 regulates the resistance of colon cancer cells to 5-fluorouracil (35). In this study, HSP27 down-regulation reduced IC50 in a dose-dependent manner. In another study on breast cancer cells, administration of paclitaxel reduced the expression of HSP27 and enhanced the chemotherapeutic effect of doxorubicin (36).
The importance of the protective effect of HSP27 expression on cell survival, the critical role of HSP27 expression in survival of osteosarcoma cells, and the close relation between HSP27 and the development of drug resistance led to the investigation of HSP27 expression in the resistant cell line in the present study. A previous study also reported that the ZOL-induced expression of HSP27 protected cholangiocarcinoma cells from caspase activation (21), supporting the current findings on the down-regulation of apoptosis induction in the resistant cell line.
Conclusion
Drug resistance to the anti-proliferative activity of ZOL was induced by prolonged treatment of HOS cells with sub-lethal concentrations of ZOL in vitro. The underlying molecular mechanism was different from the conventional efflux pump mechanism of MDR. The up-regulation of HSP27 in the resistant cell line contributes to the development of drug resistance by inhibiting apoptosis. The findings of the present study may contribute to the understanding of the anti-neoplastic mechanisms of ZOL.
Acknowledgements
We are grateful to Ms. Mizuho Kosuge and Ms. Miyuki Murayama for their skilful technical assistance and to Ms. Minako Oshiro for her skilful secretarial assistance. This investigation was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science, Sports and Culture of Japan.
- Received March 31, 2010.
- Revision received May 20, 2010.
- Accepted May 26, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved