

Abstract
Background: Malignant glioma is one of the most intractable diseases in the human body. Rho-kinase (ROCK) is overexpressed and has been proposed as the main cause for the refractoriness of the disease. Since efficacious treatment is required, this study investigated the effect of inhibition of ROCK isoforms. Materials and Methods: The short hairpin RNA transcription vector was transfected into the RT2 rat glioma cell line and the characteristics of the cells were investigated. The effect of nimustine hydrochloride (ACNU) anti-neoplastic agent on cells was also measured. Results: Inhibition of ROCK isoforms did not alter cell growth. Cell cycle analysis revealed that ROCK1 down-regulation reduced the G0 phase population and ROCK2 down-regulation reduced the G2/M phase population. When ROCK1-down-regulated cells were exposed to ACNU, they demonstrated susceptibility to the agent. Conclusion: The roles of ROCK1 and ROCK2 may be different in glioma cells. Furthermore, the combination of ROCK1 down-regulation and an anti-neoplastic agent may be useful for the therapy of malignant glioma.
- Glioma
- ROCK isoforms
- shRNA
- cell cycle
- ACNU
Malignant glioma is the most common primary neoplasm in the central nervous system and is intractable to many therapies. Despite recent advancements in surgery, radiotherapy, chemo- and immunotherapy, the prognosis remains unfavourable. Control of local recurrence is the key to therapeutic success, because glioma rarely metastasizes to tissues other than the central nervous system and the cause of death is typically due to local recurrence (1). For this reason, the development of better local therapy is required (2). One therapeutic method is local gene delivery, since direct delivery of nucleic acid such as protein-coding DNA or translation-modifying siRNA into tumours has been established and is clinically available. Regarding glioma treatment, a clinical trial using gene therapy by introducing the suicide gene with herpes virus-derived thymidine-kinase (HSV-tk) commenced in 1992 (3). Subsequently, many gene therapy protocols have been approved and used clinically throughout the world.
Cell proliferation occurs by signalling through the G-coupled protein receptor or receptor tyrosine kinase (RTK) and phosphorylating transcription factors. This signalling is related to cell migration, adhesion, contraction and gene expression. Among the G-coupled proteins, small GTPase Rho controls various signalling pathways by switching between inactive and active GDP-bound forms. Rho-kinase (ROCK) is a protein serine/threonine kinase and one of the Rho downstream targets (4, 5). ROCK is known as a regulator of actin stress fibre contraction, cytoplasm cleavage and smooth muscle contraction (6). Serotonin (7), thrombin (8), norepinephrine (9), and endothelin (10) regulate ROCK expression. In malignancies, ROCK participates in cell proliferation in glioma, breast cancer and lung cancer (11-13). Other than the ROCK, genes such as EGFR, p53 (14), FGFR2 (15) and PTEN (16, 17) have also been reported as factors controlling glioma proliferation. However, this study focused on ROCK, since this signalling pathway is a major target of various therapies (12, 13, 18). Modifications by phosphorylation of intracellular signal cascades have changed cellular adhesion, proliferation and restoration (19-21). The current study attempted to investigate the effect of down-regulation of the ROCK signal on cell proliferation using a modification of transcription with a short interfering RNA (siRNA). ROCK has two isoforms (6), ROCK1 and ROCK2, both with high homology and similarity of 65% of the protein level and 92% of the kinase domain (22). The functional difference of the ROCK isoforms has not been precisely determined in gliomas. These isoforms may play different roles in signals for proliferation, and accordingly, this study aimed to elucidate them. Lastly, in order to determine whether local siRNA transduction gene therapy can be used as a treatment for glioma in combination with a chemotherapeutic agent, the effect of ROCK1 down-regulation was measured in combination with a chemotheraputic agent, 1-(4-amino-2-methyl-5-pyrimidinyl) methyl-3-(2-chloroethyl)-3-nitrosourea: nimustine (ACNU). ACNU is a water-soluble alkylating drug that is widely used for glioma patients in Japan. This study addressed the hypothesis of whether ROCK modifications can confer sensitivity of glioma cells to ACNU.
Materials and Methods
Cell lines and animals. Rat glioma cell lines 9L (23), C6 (24) and RT2 (25) were used in the study. These cells were cultivated in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% foetal bovine serum. Adult male Fischer 344 (CDF) rats (50-70 g) were obtained from Sankyo Labo Service (Tokyo, Japan). All animal experiments were performed under permission from the Animal Care Committee at Jikei University School of Medicine.
Detection of ROCK isoform proteins by immunoblotting. Expression of both ROCK1 and ROCK2 in rat glioma cell lines was measured by immunoblotting. The exponentially growing cells were lysed by CHAPS buffer (Cell Signaling Technology Japan, Tokyo, Japan). After adjustment of the amounts of protein, an equal volume of SDS sample buffer, containing Tris-pH 6.8, 2% SDS, glycerol, β-mercaptoethanol and bromophenol blue, was added. After boiling, 10 μg samples were loaded on 7.5% SDS-PAGE gels (1.5 M Tris-HCl, 10% SDS, 30% bis-acrylamide, 10% ammonium persulfate, TEMED) and transferred to PVDF membranes (Amersham Biosciences, Little Chalfont, UK). Membranes were blocked with Block Ace (DS Pharma Biomedical, Osaka, Japan) and reacted with anti-ROCK1 mouse monoclonal antibodies (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) or anti-ROCK2 rabbit polyclonal antibodies (Santa Cruz) at a 1000-fold dilution. Specimens further reacted with respective secondary antibodies and the signals were detected by ECL (GE Healthcare, Little Chalfont, UK). The same membranes were reacted to anti-β-actin antibody to demonstrate equal loading of the samples (26).
Construction of short hairpin (sh) RNA transciption vectors to ROCK isoforms and transfection into rat glioma cells. Short hairpin RNAs (shRNA) against ROCK1 and ROCK2 were designed by the siRNA target finder (Ambion Inc., Austin, TX, USA) and corresponding DNAs were synthesised (Fasmac, Atsugi, Japan) and then ligated into the BamHI and HindIII sites of pSilencer 5.1-H1 Retro (ROCK1; Ambion) or pSilencer 3.1-H1 hygro (ROCK2; Ambion) mammalian transcription vector. After several preliminary experiments, the most effective target sequences were chosen. The target for ROCK1 was 5′-ACGCTCCGAGACACTGTAGT-3′ and that for ROCK2 was 5′-TACACAGAGATGTGAAGCCT-3′. For controls, pSilencer 5.1-H1 Retro or pSilencer 3.1-H1 hygro with a scrambled sequence were also used. These vectors transcribed shRNAs and the products functioned as short interferential RNA after cleavage. RT2 cells were chosen for the following studies because all glioma cell lines expressed almost equal amounts of ROCK1 and ROCK2 molecules and cell growth of RT2 was the most vigorous and rapid among the three cell lines. The vectors were transfected into cells by electroporation at 250 V with 975 μF and the transfected cells were selected by 150 μg/ml of puromycine (ROCK1; Cayla-InvivoGen France) or 3 μg/ml of hygromycine (ROCK2; Invitrogen, Carlsbad, CA, USA). Expression of ROCK1 and ROCK2 proteins was analysed by densitometry after immunoblotting (27).
Cell growth and doubling time in vitro. Cell growth was determined by counting the cell numbers after plating. Ten thousand cells were plated into a 60 mm dish and the total number of cells after proliferation was counted. The doubling time was calculated by the growth curve (28).
Cell cycle analysis. Cell cycles of the transfected cells were measured by a flow cytometer (FACScan; Becton Dickinson, Franklin Lakes, NJ, USA) after propidium iodide (PI) staining. A total of 105 cells were dispersed with trypsin, suspended in PBS, fixed with 75% ethanol and stained with 0.03 μM of PI with 180 units of RNaseA for 30 minutes. Populations in each cell cycle phase were analysed by Cell Quest Pro ver 5.2.1 (Becton Dickinson) (16).
Cell sensitivity assay to antineoplastic agent. A total of 104 of wild-type and transfected cells were plated in 96-well plates and exposed to varying doses (3.9 μM-8 mM) of ACNU (Daichi-Sankyo, Tokyo, Japan). After 72 hours of culture, the cells were fixed with 5% glutaraldehyde and stained with 0.01% methylene blue. Dye was eluted by 0.4 N hydrochloric acid after washing the plates. The absorbance was measured at 595 nm on a microplate reader (Bio-Rad, Hercules, CA, USA). IC50 (50% inhibitory concentration) values were subsequently calculated (29-31).
Results
ROCK expression in rat glioma cells. To confirm the expression of ROCK isoforms in various glioma cells, the expression of ROCK1 and ROCK2 was examined in 9L, RT2 and C6 cells (Figure 1). Both isoforms were ubiquitously expressed among these cell lines.
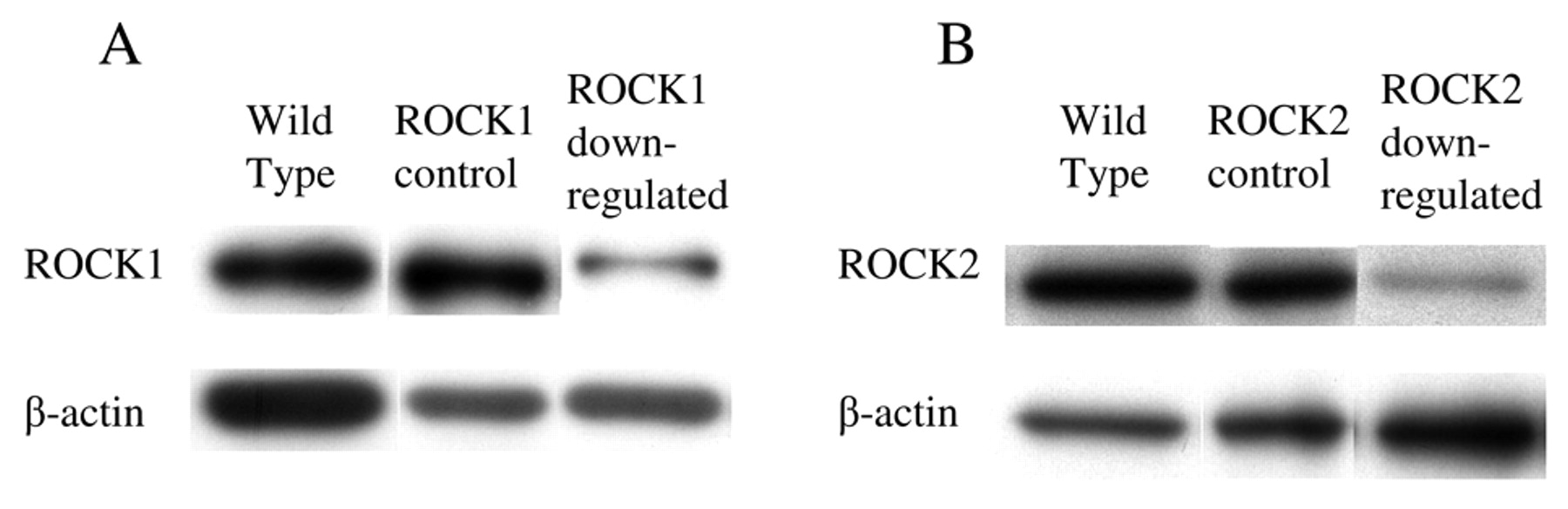
ROCK expression after shRNA transduction in RT2 cells. All cell lines expressed the ROCK isoforms. To investigate each isoform, ROCK1 and ROCK2 molecules were targeted with short hairpin RNA. After transfection of the respective vectors and selection with hygro- or puromycine, the expression of each isoform was compared. Transfectants with shRNA-containing vectors suppressed ROCK1 or ROCK2 expression, while control transfectants with scramble sequences did not alter their expression (Figure 2). There was a 57% decrease in ROCK1 and a 40% decrease in ROCK2 expression.
Cell proliferation. The doubling time of RT2 wild-type cells was 8.76 hours. When the control and target sequences for ROCK isoforms were transfected and cell proliferation was assessed, the doubling time of control cells for ROCK1, control cells for ROCK2, and those for the ROCK1- and ROCK2-down-regulated cells were 8.20 hours, 9.72 hours, 9.00 hours and 9.49 hours, respectively.

Expression of ROCK1 and ROCK2 in rat glioma cell lines.
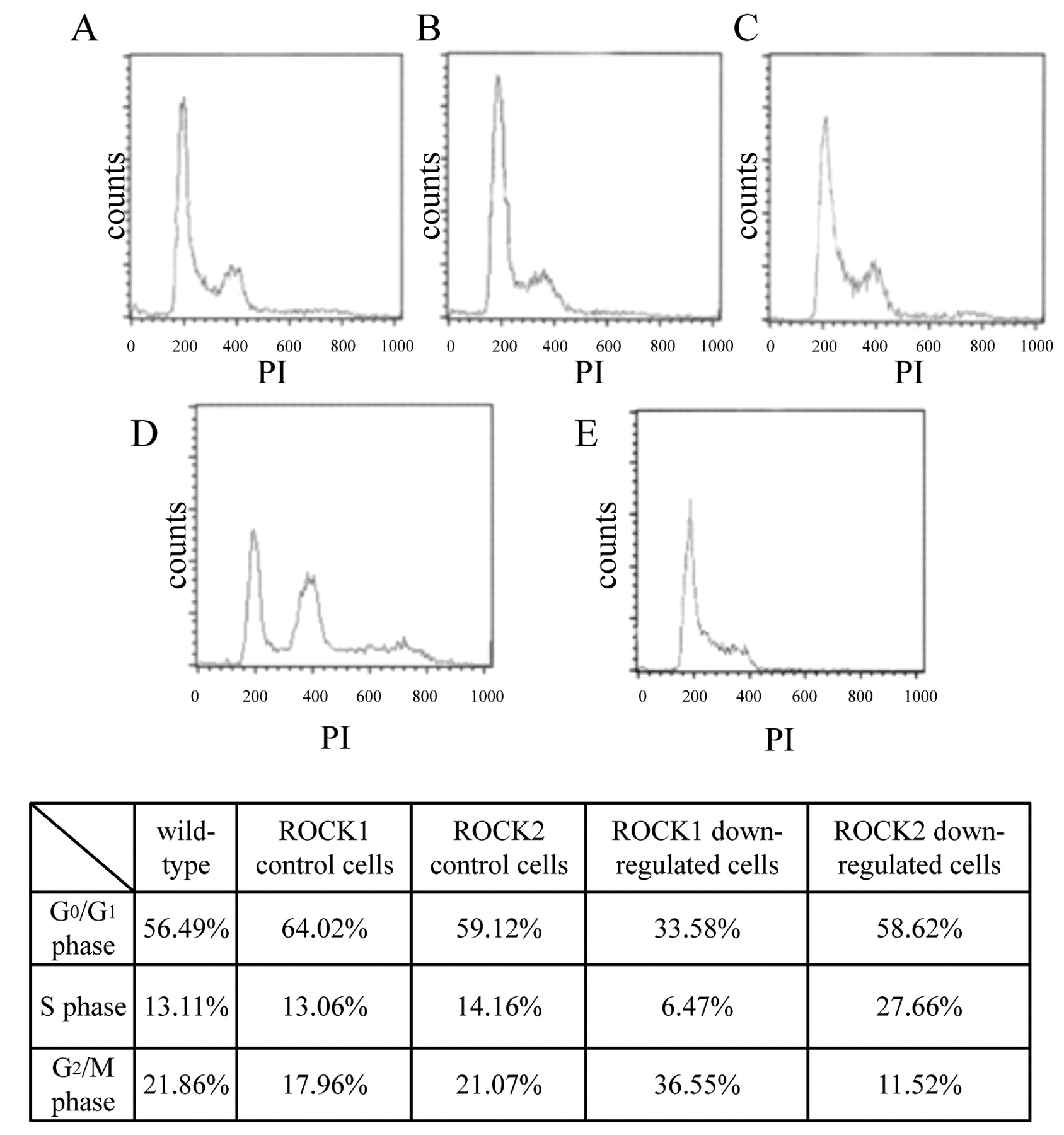
Cell cycle population analysis of transfected RT2 cells. Since the cells in which the ROCK isoform was inhibited did not show any change in cell proliferation, the effects on cell cycle were investigated. The cell population of the G0/G1, S, and G2/M phases of wild-type RT2 cells was 56.49%, 13.11%, and 21.86%, respectively. The cell cycle of the scrambled control sequences was 64.02%, 13.06% and 17.96%, respectively, for ROCK1 and 59.12%, 14.16% and 21.07%, respectively, for ROCK2. Transfection of the scrambled sequences did not alter the populations (Figure 3). In contrast, down-regulation of ROCK1 reduced the G0/G1 population (from 64.02% to 33.58%). Correspondingly, the G2/M phase population slightly increased (from 17.96% to 33.58%). On the other hand, down-regulation of ROCK2 reduced the G2/M phase population (from 21.07% to 11.52%). These results may indicate the different roles of the ROCK isoforms in the cell cycle.
Effect of ROCK modulations on the sensitivity of glioma cells to ACNU. Down-regulation of the ROCK isoforms exerted an influence on the cell cycle in glioma. Since it altered the cell cycle, its effect on drug sensitivity was evaluated. The cytotoxic assay with ACNU revealed that the control cells themselves had slightly lower IC50 values for the drug (Figure 4). IC50 values of wild-type RT2 cells, control for ROCK1 and control for ROCK2 cells were 464.86 μM, 299.35 μM and 286.90 μM, respectively. When ROCK2 was down-regulated, this modulation did not significantly alter the IC50 value (341.88 μM). However, when ROCK1 was down-regulated, it conferred sensitivity to the drug (IC50 was 171.10 μM, p<0.001 compared to ROCK1 control).
Discussion
In the present study, the different functions of the ROCK isoforms in the cell cycle of glioma cells were demonstrated and down-regulation with shRNA to ROCK1 increased sensitivity to exposure to the anti-neoplastic agent ACNU.
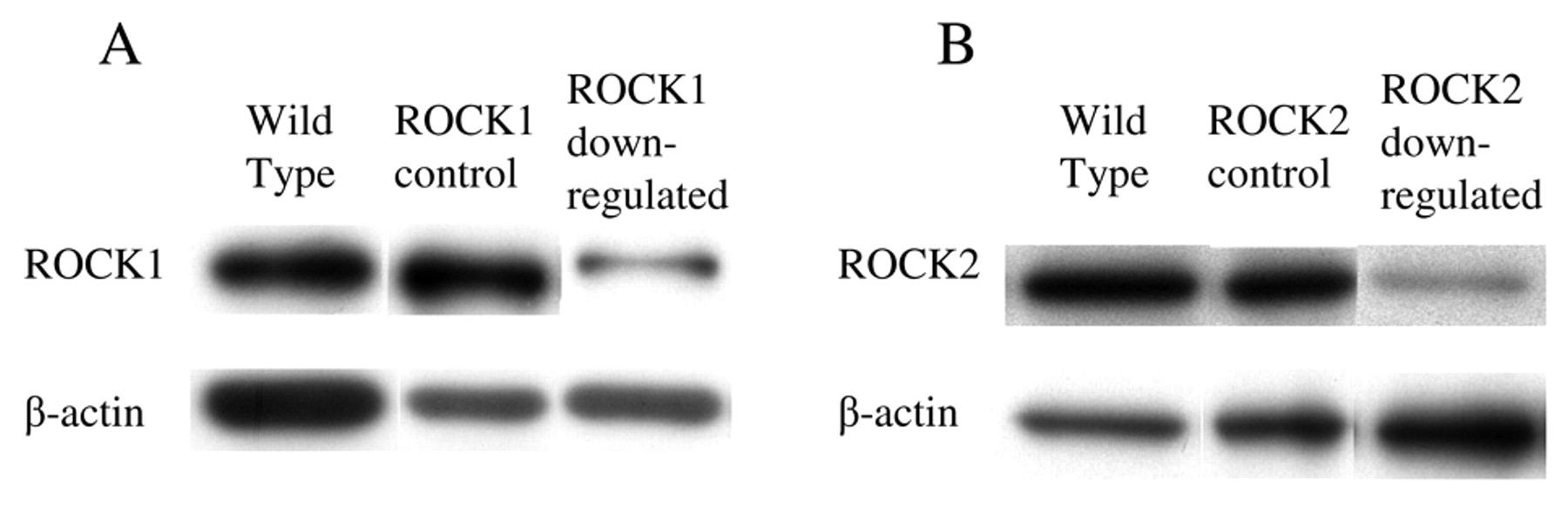
Inhibition of ROCK1 and ROCK2 in RT2 cells after shRNA Expression of the ROCK isoforms in 9L, RT2 and C6 was analysed by transfection. ROCK1 and ROCK2 in RT2 cell were down-regulated after immunoblotting (A: ROCK1; B: ROCK2). Both isoforms were expressed transfection of shRNA transcription vectors. A: ROCK1; B: ROCK2. in all three glioma cell lines tested.
Previous studies demonstrated that ROCK participates in cell proliferation. ROCK is expressed ubiquitously in the human body and has two isoforms (6, 22) with important roles in cellular function. This was true for malignant glioma cells. In the present study, the shRNA transcription vector for the ROCK isoforms was constructed and the respective down-regulated cells were obtained. However, no clones of transfection with either ROCK1 or ROCK2 shRNA vectors were grown, suggesting an essential function of the Rho kinase activity of ROCK in cell growth. Recent studies have revealed that delivery of plasmid DNA transcribing shRNA is superior to siRNA because of its effective interference and convenient production (32).
In the present study, shRNA was used for inhibition of ROCK molecules. Several regions of both ROCK isoforms were disrupted and it was found that the Rho-binding domain of ROCK1 (target: 1514-1533 of cDNA) and the kinase domain of ROCK2 (target: 844-863 of cDNA) were the most favourable. Although the target regions were selected beforehand, down-regulation of each isoform expression was moderate. However, the effects were sufficient to alter the cell cycle.
A subdermal tumour of RT2 cells was implanted and established in the syngenic Fischer 344 rat and the expression of ROCK1 and ROCK2 was compared. Immunohistochemical examination showed that ROCK1 was expressed strongly in several cells, demonstrating a patchy and isolated pattern in the tissue. In contrast, ROCK2 was diffusely and weakly expressed throughout the specimens (data not shown). In general, ROCK2 is expressed strongly in normal brain cells (22), however tumourigenic transformation has been found to suppress ROCK2 (27). This may be attributed to the weak expression of ROCK2 in established RT2 cells. In any case, localisation as well as the role of ROCK isoforms in the cell cycle was different in glioma cells. There have been various reports on how ROCK relates to the cell cycle, but it is not clear how ROCK1 and ROCK2 affect the cycle in glioma cells (33-35). The present study revealed that the G0/G1 phase population decreased after down-regulation of ROCK1 and the G2/M phase population decreased after down-regulation of ROCK2. These down-regulations did not significantly alter the doubling time of cell growth. These results may be useful in further analysis of the participation of ROCK molecules in the cell cycle.

Effect of down-regulation of ROCK1 and ROCK2 on the cell cycle. ROCK1 down-regulation decreased the G0/G1 phase population and increased the G2/M phase population. ROCK2 down-regulation decreased the G2/M phase population. The horizontal axis indicates the amount of DNA, and the vertical axis indicates the cell number. A: RT2 wild type; B: ROCK1 control cells; C: ROCK2 control cells; D: ROCK1 down-regulated cells; E: ROCK2 down-regulated cells.
Regarding therapy for glioma, sole inhibition of ROCK1 or ROCK2 did not change the cell growth. Moreover, when implanted into syngenic Fisher 344 rat subdermally or intracranially, both down-regulated cells were tumourigenic (data not shown). These findings suggest that tumours may not be suppressed unless both ROCK1 and ROCK2 isoforms are simultaneously down-regulated.
ACNU is a nitrosourea derivative that bridges between DNA and inhibits DNA division without arresting the cell cycle (36). Since the drug is one of the most common chemotherapeutic agents used in Japan (37, 38) with a good track record of patients who have been clinically treated, its combination effect with shRNAs was investigated. Down-regulation of ROCK1 augmented the efficacy of ACNU. Due to the cross-linking ability of ACNU, it was presumed that the effect of ACNU on G2/M phase cells, which were increased by ROCK1 down-regulation though the drug, was not cell cycle specific. Since the IC50 value in response to ACNU decreased and ROCK1 down-regulation increased the sensitivity of the cells, the combination of shRNA transfection gene therapy for ROCK1 with systemic ACNU treatment may be effective for patients bearing malignant glioma. Involvement of the ROCK isoform in the cell cycle of malignant glioma is important and further studies on this area are warranted.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Sensitivity of ROCK down-regulated cells to ACNU. Wild-type, ROCK1 and ROCK2 down-regulated cells were exposed to ACNU for 72 hours. After exposure, these cells were fixed, stained with methylene blue, and then the dye was eluted by HCl. Absorbance was measured at 600 nm. The value of the 50% inhibitory concentration (IC50) of wild-type cells was 464.8 M. Transfections of control vectors demonstrated increased sensitivity. Compared to the control cells, down-regulation of ROCK1 decreased the IC50 value. ROCK2 down-regulation did not change the sensitivity. N=4; Bars: standard deviation. ○: wild type RT2; □: ROCK1-control; ▵: ROCK2-control; ▪: ROCK1-down-regulated; ▴: ROCK2-down-regulated.
- Received June 8, 2010.
- Revision received June 29, 2010.
- Accepted July 6, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved