

Abstract
The pharmacology of progestins includes actions initiated by various cellular targets, including classic receptors characterized as nuclear transcription factors (nPR), G-protein-coupled membrane receptors (mPR), enzymes, membrane channels and transporters. The effects initiated by targets other than nPR are termed non-genomic and there is an increasing recognition that these effects also play an important role in the regulation of cell growth. Materials and Methods: The nPR-positive breast cancer (MCF-7) and the nPR-negative uterine cervix cancer (C4-I) cell lines were exposed to progesterone (PG) and mifepristone (MF) during a culture period of 96 h. Daily cell count, cell cycle analysis and apoptosis assay were performed. Results: It was possible to separate the nPR initiated effects (growth stimulation) from the non-genomic effects (growth inhibition) in the MCF-7 cells. Below 1 μM PG treatment gave a small, but distinct increase in cell density which was effectively blocked by MF. Such an effect was absent from the nPR-negative C4-I cells. For a range of concentrations between 1 μM and 100 μM, the effect of both PG and MF developed over time and showed concentration dependency. The PG concentrations needed to reduce cell density by 50% (IC50) were 12.8±1.1 μM and 6.5±0.2 μM for the MCF-7 and C4-I cells, respectively. MF appeared to be equally or slightly more potent, with respective IC50 values of 6.9±0.5 μM and 5.3±0.3 μM. The cell density reduction was both a result of cell cycle arrest and apoptosis. The combination of PG and MF had a potentiated effect on cell density reduction, cell cycle arrest and apoptosis. Conclusion: The antiproliferative/cytotoxic effect of PG and MF in concentrations between 1 and 100 μM is of a non-genomic nature.
- Progestins
- antiprogestins
- cytotoxic
- breast cancer cells
- non-genomic effects
- mifepristone
- apoptosis
- cell cycle
Steroid receptors function as ligand-dependent transcription factors to control the expression of specific genes (1). The effect on gene expression or repression can be observed after 30 minutes (2). However, so-called non-genomic effects (or effects of extranuclear initiated signaling), first recognized due to their rapid onset, have also been identified. Numerous non-genomic effects of steroids have been observed and their mechanisms characterized (2-6). The successful treatment of the premalignant condition endometrial hyperplasia with the progestin levonorgestrel (7, 8) is, at least partly, of a non-genomic origin, based on the observation that the effect of levonorgestrel intrauterine system (IUS) was maintained, even with complete nuclear transcription factor progesterone receptor (nPR) down-regulation (9). The assumption of a non-genomic mechanism was also supported by in vitro studies wherein progesterone (PG) caused a concentration-dependent reduction of nPR-negative cells (10-12). Finally, mifepristone (MF), a potent receptor antagonist (13), was unable to reverse the antiproliferative effect of supraphysiological PG concentrations in nPR-positive cells (14). In addition to the nPR, membrane G-protein coupled progesterone receptors (mPR) have been characterized (15). However, other cellular targets such as adenylate cyclase (16), guanylate cyclase (17), mitogen-activated protein kinases (MAPK)/extracellular signal-regulated kinases (ERKs) (18, 19), may also be involved in non-genomic PG effects.
In the present study we employed human nPR-positive breast cancer cells, MCF-7 (20), to further characterize the non-genomic effect of PG and MF. Receptor negative C4-I cells were utilized as a control system for the nPR mediated effects.
Materials and Methods
Chemicals. PG and MF were purchased from Sigma Aldrich Chemical Company, St. Louis, MO, USA. All other chemicals used were of analytical grade.
Cell culture. Two human cell lines were employed. The breast cancer cell line MCF-7 was obtained from the European Collection of Cell Cultures (Salisbury, UK) and grown in Eagle's Minimal Essential Medium (Sigma Aldrich) supplemented with 10% (v/v) fetal bovine serum (Gibco Ltd., Paisley, UK), and 0.10 g/l streptomycin, 0.060 g/l penicillin G, 2 mM L-glutamine and 1 mM Na-pyruvate (Sigma Aldrich). The human carcinoma of the uterine cervix cell line C4-I was obtained from the American Type Culture Collection (Rockville, MD, USA) and cultured in RPMI-1640 medium (Sigma Aldrich) supplemented with 10 % (v/v) newborn calf serum (BioWhittaker Inc., Walkersville, MD, USA), 0.10 g/l streptomycin (Sigma Aldrich) and 0.060 g/l penicillin G (Sigma Aldrich). Phenol red was omitted from the culture media to avoid steroid-mimicking effects (21). The seeding density of MCF-7 cells was 4.3-4.8×104 cells/ml and 5.1×104 cells of C4-I cells (if not otherwise stated). After 24 h the cells were adherent, and medium including active substances were added and renewed at intervals of 24 h. Doubling times for the MCF-7 and C4-I cells were 36-37 h and 29-33 h, respectively. Before conducting the experiments we verified that the MCF-7 cells and the C4-I cells were nPR-positive and nPR-negative, respectively, with an assay described previously (11).
Cell cycle kinetics and apoptosis. The cells were harvested and cell cycle analysis and quantification of apoptosis were performed daily by flow cytometry employing a FACScan flow cytometer (Becton Dickinson Immunocytometry Systems, San Jose, CA, USA) equipped with an argon-ion laser (488 nm). The assay of cell cycle kinetics was carried out as described previously (22). Two different approaches were used to study apoptosis. The appearance of a hypodiploid DNA peak (sub-G1 fraction) indicates the presence of an apoptotic cell population (23). Induction of apoptosis was also determined by the JC-1 assay, wherein the changes in the mitochondrial membrane potential (Δψ) were detected, and performed as reported earlier (22).
Determination of IC50 values. The effect on cell densities was characterized by the concentrations (IC50) needed to cause 50% reduction in cell densities. The IC50-values were determined according to Chou (24).
Statistics. The results are presented as mean value±SEM if not otherwise stated.
Results
Effects of PG and MF on cell density. In the presence of physiological PG concentrations (up to approximately 1 μM) the MCF-7 cell densities showed a small but distinct elevation above the control values (Figure 1), but the effect diminished and disappeared for higher concentrations. The addition of MF blocked this response to PG (Figure 1). No such effect was detectable for the nPR-negative C4-I cells (results not shown).
Figure 2 shows that PG concentrations above 1 μM caused a steep fall in cell densities. The IC50 was 12.8±1.1 μM (n=3) after 96 h treatment. For concentrations above 1 μM MF had a virtually identical effect with an IC50 of 6.9±0.5 μM (n=3). MF reinforced the effect of PG with an apparent slight increase in the sensitivity to PG (IC50=5.6±0.3 μM, n=3).
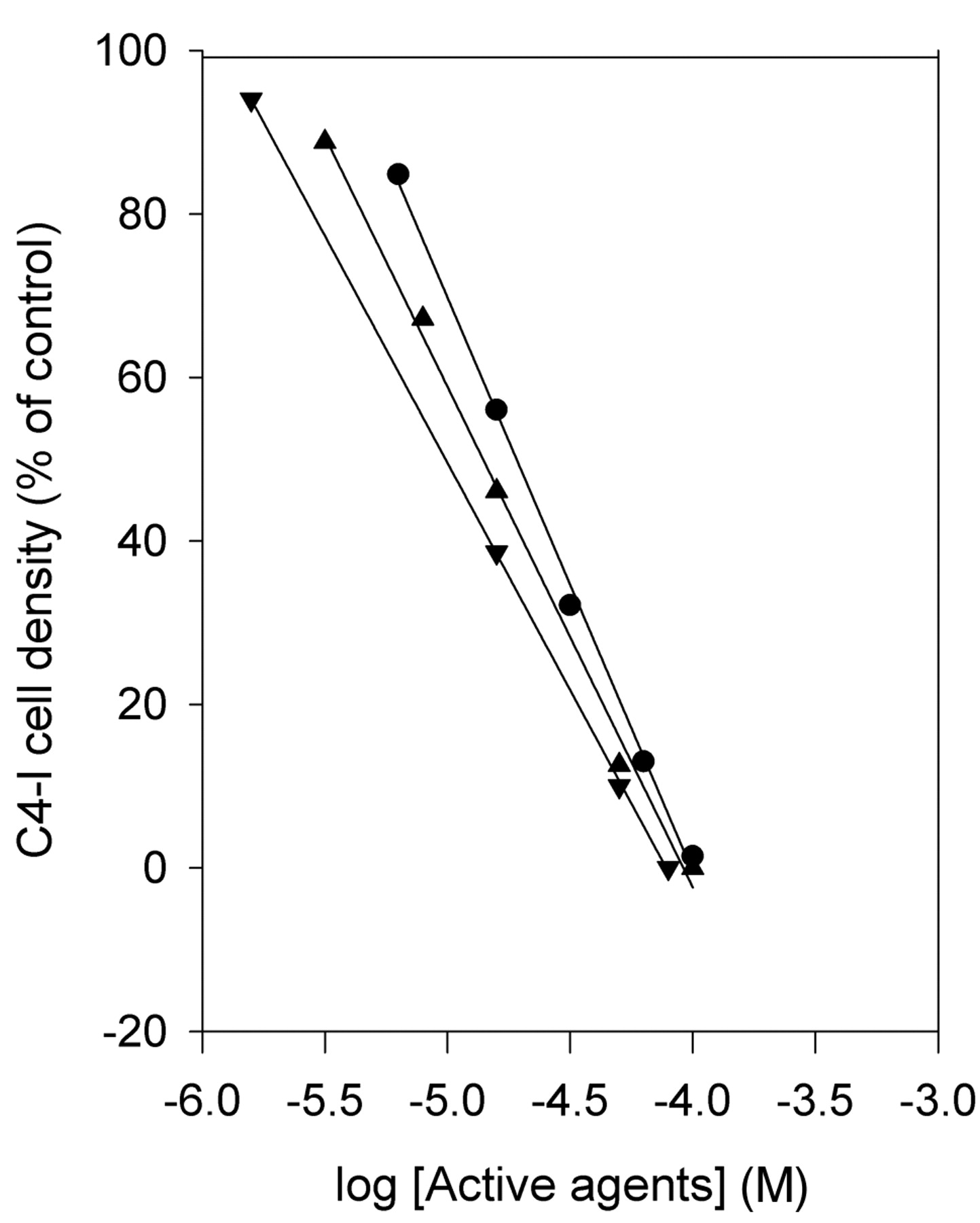
The effect of PG on the C4-I cell densities showed an identical pattern for concentrations above 1 μM (Figure 3) with an IC50 of 6.5±0.2 μM (n=3) after 96 h. MF showed an IC50 of 5.3±0.3 μM (n=3) and potentiated the effect of PG. The apparent IC50 for PG in the presence of MF was 3.5±0.2 μM (n=3).

The nPR initiated effect on MCF-7 cell densities by physiological concentrations of PG (<1 μM) in the absence (black columns) or presence of MF (red columns) 96 h after seeding. Mean values, n=3.
Effects of PG and MF on cell cycle kinetics. Flowcytometric analysis was performed for both MCF-7 and C4-I cells to determine if cell cycle arrest contributed to the cell density reduction. When the MCF-7 cells were seeded at a density of 4.3×104 cells/ml, both PG and MF caused a clear G1/G0 retardation after 72 h with a fractional increase of 12% and 16%, respectively (Table I). At a higher seeding density the fraction of the cell population which resided in the G1-phase was higher (Table II). After 72 h the relative retardation after PG was less pronounced (8%) or identical after MF (16%). A continuous development of the cell cycle retardation was observed during the culture period, in both the control and the exposed cells (Table II). In addition, a concentration-dependency of PG and MF was evident (data not shown). In these experiments the highest concentrations (95-159 μM PG and 70-116 μM MF) caused massive cell death after 96 h treatment (results not shown). This situation made impossible to perform reliable flow cytometric analysis.
The C4-I cells were seeded at a high density (56×104 cells/ml) to secure optimal assay conditions. A continuous development of the cell cycle retardation was observed during the culture period, in both the control and the exposed cells (Table III). Treatment with 32 μM PG or 23 μM MF resulted in no clear differences from the controls.
Apoptosis. A sub-G1 fraction was observed in both cell types after exposure to PG and MF (Table IV). The development of nuclear fragmentation was dependent both on the duration of culture and the concentration of the active agents. Table V showed that the development of apoptosis (JC-1 assay) was dependent on time after seeding in addition to concentrations of PG and MF. The highest tested concentrations (95 μM PG and 70 μM MF) caused extensive apoptosis after 96 h exposure and the combination of PG and MF reinforced the effect.

The non-genomic effect on MCF-7 cell density after treatment with PG (● - ●), MF (▴ - ▴), or a combination of PG and MF (▾ - ▾) 120 h after seeding. Mean values, n=3.
Discussion
A clear difference between the MCF-7 cells with classical nuclear receptors (20) and C4-I cells without was observed for low concentrations of PG. A small, but distinct, increase in MCF-7 cell density occurred with PG, but was blocked by MF. This was the expected response in an nPR-positive cell type since MF binds with high affinity to the receptors and effectively competes for binding (13).
Two types of PG receptors have been distinguished, nuclear (nPR) and membrane G-protein-coupled receptors (mPR). The concentrations needed to obtain half maximal binding (Kd) to nPR have been reported to be 1-4 nM (25, 26). The binding affinity for mPR was lower, with a Kd of about 150 nM (27). Receptors of both types have also been identified in MCF-7 cells (15, 20). Furthermore, MF displays no binding affinity for human recombinant mPR (28). These observations argue against a role of the nPR or mPR in the antiproliferative effect of PG and MF at high concentrations (>1 μM).
{kind=link}
{kind=link}
{kind=link}
The non-genomic effect on C4-I cell density after treatment with PG (● - ●), MF (▴ - ▴), or a combination of PG and MF (▾ - ▾) 120 h after seeding. Mean values, n=3.
MCF-7 cell cycle distribution 96 h after seeding. The effects of 32 μM PG or 23 μM MF are shown (n=4).
Above the PG concentrations which saturate nPR (five times the Kd, i.e. 5-20 nM), so-called non-genomic or extranuclear-initiated effects occur. Many cellular targets have been identified for the non-genomic PG effects, including cell growth and apoptosis. Examples are adenylate cyclase (16), guanylate cyclase (17), MAPK/ERK (18, 19), Src tyrosine kinase (29), phosphatidylinositol 3-kinase (PI3K), (30), P-glycoprotein (31-33) and multidrug resistance protein 5 (MRP5) (34-36).
In the present study, PG and MF exerted a very potent antiproliferative effect at concentrations between 1 and 100 μM. After 96 h the highest concentrations caused almost complete cell death of MCF-7 and C4-I cells. The values for IC50 were 7-12 μM for PG and 5-7 μM for MF. These values for PG are in agreement with previous reports (2.1 μM; 12) and calculated IC50 values based on previous results [4.5 μM (10) and 6.0 μM (11)] for the nPR-negative C4-I cells. In a recent study on the nPR-positive Ishikawa cells, somewhat higher values were found for PG and MF, with IC50 of 45 μM and 19 μM, respectively (14). However, in both MCF-7 and Ishikawa cells the combination of PG and MF (1-100 μM) reinforced the effect of each other suggesting a common target. Such an effect was also seen for the nPR-negative cell type C4-I. The present observation that MF was a potent inducer of apoptosis is supported by several in vitro studies (37-40).
MCF-7 cell cycle distribution after high seeding density (13.3×104 cells/ml). The effects of 32 μM PG or 23 μM MF are shown. Mean values of three independent experiments.
C4-I cell cycle distribution after culture with 32 μM PG or 23 μM MF. The cells were seeded at a density of 56×104 cells/ml. Mean value of three independent experiments.
Sub-G1 fractions. MCF-7 cells were seeded at a density of 13.3×104 cells/ml and C4-I cells at 56×104 cells/ml. The proportion of MCF-7 and C4-I cells in the sub-G1 fraction is given as the percentage of the G1/G0 phase cell population (mean value, n=3).
Mitochondrial membrane potential. The MCF-7 and C4-I cells were seeded at densities of 80.8×104 cells/ml and 11.2×104 cells/ml, respectively. Mean values of three independent experiments.
A possible common target is ATP-binding cassette (ABC) transporters which are inhibited by both PG and MF. Examples are P-glycoprotein (31-33, 41, 42) and MRP5 (34-36). Even if ABC transporters are potential targets which initiate an antiproliferative effect of PG and MF in the concentration range 1-100 μM, other molecular target(s) may exist as well. A role for P-glycoprotein in apoptosis has been suggested (43) by suppressing caspase activation independent of ATPase activity (44).
Clinical studies have shown that apoptosis is an important mechanism in the successful treatment of endometrial hyperplasia (45-47), a precursor of endometrial carcinoma. In contrast to the efficacy of PG and MF at inducing apoptosis, the cell cycle retardation was dependent on cell density and was clearly much higher after low- than high- seeding density, shown for MCF-7 cells in the present study. Cell density inhibition is itself presumed to retard the cell cycle, consequently the effect of the active agents diminishes.
If progestins and antiprogestins were to be introduced into clinical practice for the treatment of proliferative disorders, drug monitoring would be needed since the therapeutic index appears to be narrow (1-100 μM for PG and MF). Doses of MF, considered to be high (400-600 mg), resulted in peak plasma concentrations in a range from 4 to 12 μM, but with huge interindividual variations (48). The combination of PG and MF would be advantageous partly by blocking the nPR and by a more powerful non-genomic effects due to the synergistic interaction.
Acknowledgements
The study was supported by grants from the Norwegian Cancer Association. The technical assistance by Ragnhild Jæger and Natalia Smaglyukova is greatly appreciated.
- Received August 17, 2010.
- Revision received November 10, 2010.
- Accepted November 12, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved