

Abstract
Background: Anti-proliferation and apoptosis inducing effects of green tea polyphenols (GTPs) were studied against human promyeolcytic leukemia HL-60 cells. Materials and Methods: Anti-proliferation activity of GTPs against HL-60 cells and cytotoxicity against normal cells, V79-4 were measured with the MTT assay. Nuclear fragmentation of GTP-treated cells was observed with a fluorescence microscope and flow-cytometric analysis was performed to observe the appearance of apoptotic bodies. Changes in apoptosis-related protein levels were investigated by Western blot analysis. Results: GTPs significantly reduced the HL-60 proliferation, however, they did not show significant inhibition in proliferation of normal V79-4 cells. GTP treatment induced apoptosis in HL-60 cells, evidenced by nuclei fragmentation and apoptotic body appearance. The expression level of Bcl-2 was decreased and caspase-3 was activated by GTP treatment. Conclusion: GTPs revealed anti-proliferative and apoptosis-inducing activity against HL-60 cells through the down-regulation of Bcl-2 and activation of caspase-3.
- Green tea polyphenols
- apoptosis
- Bcl-2
- caspase-3
Green tea has been the most consumed beverage next to water for many centuries in Asian countries. The beneficial effects of green tea are well known including cancer preventive activity and other medical applications (1-3). Green tea is chemically characterized by the presence of large amounts of polyphenolic compounds which may account for up to 30% of its dry weight. There are several polyphenolic catechins in green tea including (-)-epigallocatechin 3-gallate (EGCG), (-)-epigallocatechin (EGC), (-)-epicatechin gallate (ECG), (-)-epichatechin (EC), (+) catechin and (+) gallocatechin. Most of the medical activities of green tea seem to be associated with the epi- forms rather than the catechins (4).
The potential health beneficial effects of green tea and its constituent catechins have been extensibly studied (5-9). Many studies have focused on the antioxidative effects of green tea and its constituent catechins and it is suggested that its potential physiological effects for the protection and treatment of cancer and cardiovascular diseases come from its antioxidant activity (1, 10). It is reported that many antioxidant substances have anticancer or anticarcinogenic properties. For example, resveratrol in grapes and ellagic acid in green tea have been shown to protect cells from oxidative damage and cell death (11, 12). Despite there being many reports regarding green tea constituents, not many studies have been performed with green tea polyphenols (GTPs). The present study was therefore designed to investigate the induction of anti-proliferative activity of GTPs and to characterize the mechanism of its apoptosis inducing effect in human promyelocytic leukemia HL-60 cells.
Materials and Methods
Chemicals. Green tea polyphenols (GTPs) were purchased from LKT laboratories, USA. GTPs were solubilized in deionized water and the components of GTPs were analyzed by high-performance liquid chromatography (HPLC; Agilent Technologies, Japan) with reverse-phase C18 column. Mobile phase was CH3CN: H2O (13:8) mixture and flow rate was 1 mL/min. Samples were detected with ultraviolet light at 203 nm. All other chemicals were of the highest analytical grade and were purchased from Sigma, USA.
Cell culture, cell viability and cytotoxicity analysis. The human promyelocytic leukemia cell line HL-60 (ATCC CCL-240) was grown in RPMI-1640 (Gibco, BRL, USA). The Chinese hamster lung cell line V79-4 (ATCC CCL-93) was grown in Dulbecco's modified Eagle's medium (DMEM; Gibco, BRL, USA). Culture media was supplemented with 10% (v/v) heat inactivated fetal bovine serum, 2 mM glutamine and antibiotics. Cultures were maintained at 37°C in a humidified atmosphere of 95% air and 5% CO2. Cytotoxicity of GTPs was estimated by the MTT assay (13). HL-60 cells were treated with GTPs for either 48 or 72 h. V79-4 cells were treated with GTPs for 48 h. The data are expressed as a mean percentage of viable cells as compared to the respective control cultures.

HPLC identification of GTP constituents.
Nuclear morphology and flow cytometric analysis. HL-60 cells were treated with 0, 20, 50 and 100 μg/mL of GTPs for 24 and 48 h. Morphology of cellular nuclei were observed as described previously (14). HL-60 cells were treated with 0, 20, 50 and 70 μg/mL of GTPs for 48 h and used for flow-cytometric analysis as described previously (15).
Western blot analysis. HL-60 cells were treated with GTPs and subjected to Western blot analysis, as described previously (15). Blots were probed with mouse monoclonal anti-human anti-Bcl-2 (Oncogene Science, Cambridge, MA, USA), anti-caspase-3 (Transduction Laboratory, Lexington, KY, USA), and anti-PARP (Santa Cruz Biotechnology, Santa Cruz, CA, USA) antibodies. Immunoreactivity was detected using either an anti-mouse (Santa Cruz Biotechnology) or anti-rabbit (Amersham Biosciences, Buckinghanshire, UK) peroxidase-conjugated secondary IgG antibody and an enhanced chemiluminescence (ECL) detection kit (Amersham Biosciences).
Results
Anti-proliferative effect of GTPs. The component of GTPs used in this study was analyzed by using HPLC. As shown in Figure 1, various polyphenol components were identified. Among those, EGCG which is known as the main ingredient of green tea comprised 37.9% . The content of ECG and EGC were 18.9% and 14.2%, respectively. The content of EC was 11.0% and all other components constituted about 18.0% .
The anti-proliferative effect of GTPs on a promyelocytic leukemia cell line, HL-60, was evaluated with MTT assay. When cells were treated for 48 or 72 h with 0, 3, 10, 30, 100 or 300 μg/mL of GTPs, the relative cell proliferation progressively decreased in a dose-dependent manner, as shown in Figure 2. The IC50 value for GTPs on HL-60 cells was 49.5 μg/mL and 50.0 μg/mL at 48 and 72 h incubation, respectively. In order to observe cytotoxicity of GTPs on a normal cell line, the relative cell survival of a Chinese hamster cell, V79-4 upon treatment of GTPs was measured. As shown in Figure 3, there was no significant cytotoxic effects observed on 48 h incubation with GTPs.

Anti-proliferative activity of GTPs in HL-60 cells. HL-60 cells were incubated with 0, 3, 10, 30, 100 and 200 μg/mL of GTPs for 48 h (●) or 72 h (○). Cell survival was measured by the MTT assay. Data are presented as means±SD. The percentage of cell growth in the control group was designated as 100% .

Cell survival of a normal cells V79-4 after treatment of GTP. V79-4 cells were incubated with 0, 25, 50 and 100 μg/mL of GTPs for 48 h. Cell survival rate was measured by MTT assay. Data are presented as means±SD. The percentage of cell growth in the control group was designated as 100% .
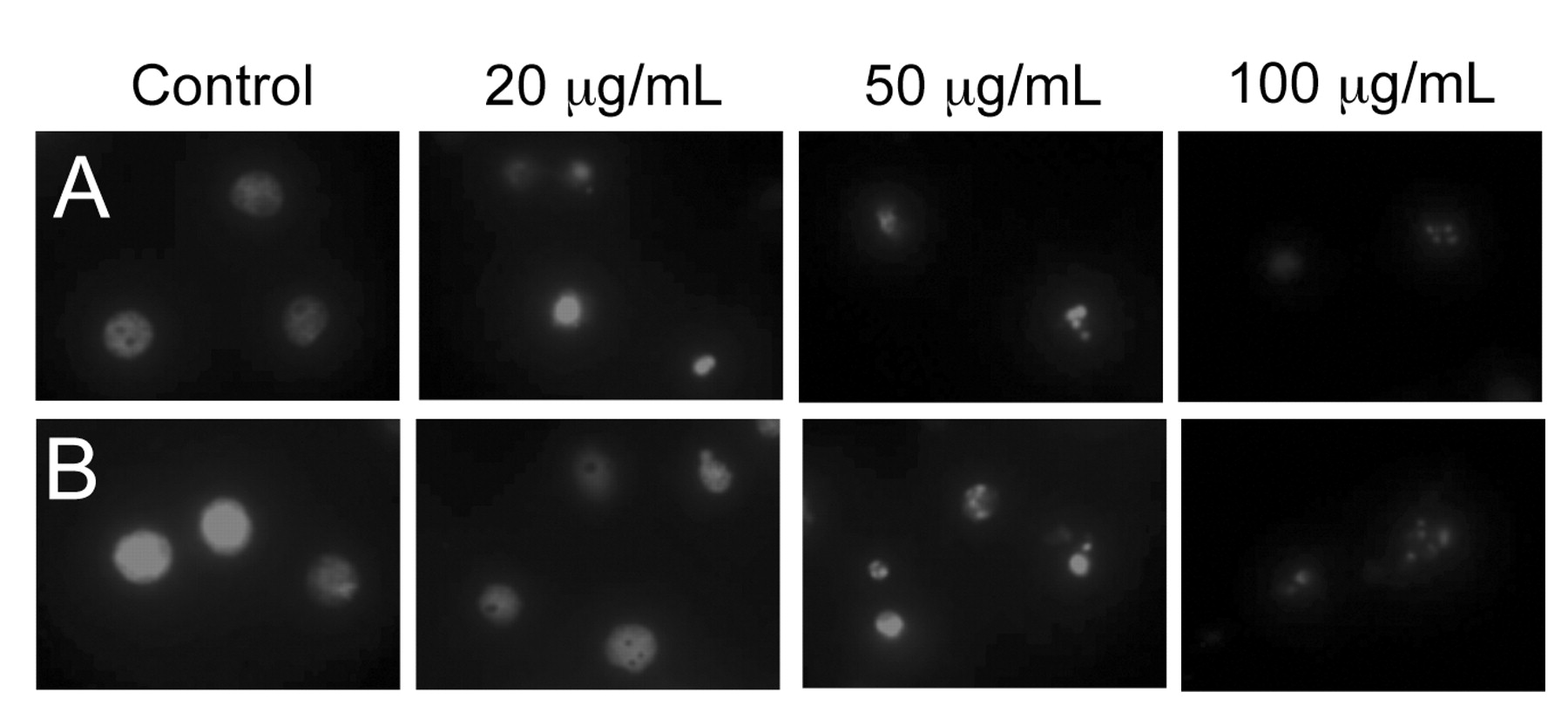
Induction of apoptosis by GTPs treatment. To determine whether GTP-meditated inhibition of growth and proliferation was associated with apoptosis, the GTP-induced nuclei fragmentation in HL-60 cells were examined. Cells were treated with various concentrations of GTPs and the nuclei were stained with propidium iodide. As shown in Figure 4, control cells exhibited intact nuclei, but cells treated with GTPs showed significant dose-dependent nuclear fragmentation. More fragmentation was observed in cells treated with GTPs for 48 h than cells treated for 24 h (compare Figure 4A and B).
The induction of apoptotic bodies in GTP-treated HL-60 cells was further analyzed by flow-cytometric determination of DNA content. Histograms of DNA content obtained from propidium iodide-stained HL-60 cells showed that the percentage of cells with reduced DNA content dose-dependently increased. The percentages of apoptotic cells observed at 0, 4, 20, 50 and 70 μg/mL of GTPs were 3.45%, 15.5%, 21%, 65.7% and 91%, respectively (Figure 5). The profile for the GTP-induced increase in hypodiploid DNA content closely correlated with the nuclei fragmentation. In parallel to the increase in the number of cells with sub-G1 hypodiploid DNA content, there was a decrease in the number of cells with diploid DNA content.

Apoptotic bodies were induced by GTP treatment. HL-60 cells were treated with 0, 20, 50 and 100 μg/mL of GTPs for 24 h (A) or 48 h (B) and stained with propidium iodide. Cellular morphological changes were observed using a fluorescence microscope at the magnitude of ×600.

Flow cytometric analysis of GTP-treated HL-60 cells. Cells were stained with propodium iodide after treatment with 0, 4, 20, 50 and 70 μg/mL of GTPs for 48 h. DNA content analysis was performed using a flow cytometry. Representative histograms are shown.
Decrease in Bcl-2 expression and activation of caspase-3 by GTPs treatment. To understand the molecular mechanisms by which GTPs induced apoptosis, we examined various apoptosis-related proteins. HL-60 cells were cultured in media containing 0, 4, 20, 50 and 100 μg/mL of GTPs for 48 h. After incubation, total protein was isolated and Bcl-2, caspase-3 and PARP [poly(ADP-ribosyl)polymerase] immunoreactivity levels were measured by Western blotting (Figure 6). The levels of the anti-apoptotic molecule Bcl-2 decreased. Pro-caspase-3 levels also decreased, implying that the levels of active caspase-3 had increased. To investigate the enzymatic activation of caspase-3, the cleavage of PARP, which is a caspase-3 substrate, was measured. When cells were treated with GTPs, a dose-dependent increase in the formation of the 85 kDa fragment and a decrease in the formation of the 116 kDa PARP were observed.
Discussion
Many health beneficial effects of GTPs have been suggested for a variety of disorders due to their antioxidative activity (10). Green tea catechin extract intake increased the life span of fruit flies through the increase in the activity of the antioxidant enzymes, superoxide dismutase and catalase (16). Oxidative stress induced by reactive oxygen species have been prevented by green tea polyphenol treatment in human microvascular endothelial cells (17). The polyphenolic fraction of green tea has been suggested to prevent UV radiation-induced skin cancer (18).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Changes in the expression of apoptosis-related proteins in response to treatment with GTPs. HL-60 cells were treated with 0, 4, 20, 50 and 100 μg/mL of GTPs for 48 h. Cell extracts were subjected to western blotting to determine immunoreactivity levels of Bcl-2, procaspase-3 and PARP as described in Materials and Methods. Representative Western blots are shown.
A significant number of naturally occurring substances have been shown to protect against experimental carcinogenesis. Thus, it is becoming increasingly evident that certain phytochemicals, particularly those included in the daily diet, have important cancer chemopreventive properties (19). Some anti-inflammatory chemopreventive agents have been found to suppress growth and proliferation of transformed or malignant cells through induction of programmed cell death or apoptosis (20). Several antioxidative compounds and extracts have been shown to protect against experimental carcinogenesis and to induce apoptosis of cancer cells. Extracts from B. platyphylla var. japonica showed antioxidative effects and apoptosis inducing effects (21). Growth inhibition and apoptosis inducing mechanism of GTPs are under extensive study. The growth inhibitiory effect of GTPs and its constituents have been reported. Among its constituents, EGCG and EGC displayed strong inhibitory effects against two lung cancer cell lines and evidence of apoptosis induction has also been suggested (9). The majority of the studies have been performed with the main ingredient, EGCG (22-25) and there are not many reports regarding anticancer or apoptosis-inducing effects of GTPs.
In this work, the apoptosis-inducing effects of GTPs on human preomyelocytic leukemia HL-60 cells have been investigated. The presented results demonstrated that GTPs inhibited cell growth inhibition in HL-60 cells but not in a normal V79-4 cells. In addition, apoptosis was induced in GTP-treated HL-60 cells. The induction of apoptotic cell death was accomplished by characteristic morphological and structural changes. Nuclei fragmentation and appearance of sub-G1 apoptotic cell population in GTP-treated cells consistently suggest the induction of apoptosis. It has been reported that apoptosis is not induced in caspase-3 null cells even after GTP treatment, which suggested the involvement of caspase-3 in apoptosis induced by GTP treatment (26). In this study, it has been shown that caspase-3 was activated and that a substrate of caspase, PARP, was proteolytically cleaved to low molecular weight fragments in GTP-treated HL-60 cells. Apoptosis induction effects of GTPs in other cancer cells were reported. GTPs inhibited the proliferation, arrested cell cycle progression and induced apoptosis of human breast cancer cells (27). GTPs showed growth inhibitory and anti-metastatic effects on mouse mammary carcinoma cells (28). In addition to caspase-3 activation, it was observed that the levels of Bcl-2, an anti-apoptotic protein, decreased upon treatment of GTPs, resulting in a decrease in the ratio of Bcl-2/Bax, one of the major events that regulate apoptosis. These findings suggest that GTPs exhibit growth inhibition and apoptosis induction activity in HL-60 cells through the down-regulation of Bcl-2 and activation of caspase-3.
Acknowledgements
This work was supported by the Seoul RNBD Program (10524) in Korea.
- Received August 14, 2008.
- Revision received December 17, 2008.
- Accepted January 26, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved