

Abstract
Background: Among the combinations of coamplified genes, Myc and ERBB2 or EGFR have attracted much attention for their relevance to cytogenetics, carcinogenesis and cancer therapy. Materials and Methods: Gene amplification of Myc, ERBB2 and EGFR were examined on 97 formalin-fixed and paraffin-embedded gallbladder carcinomas, by fluorescence in situ hybridization (FISH). Results: FISH revealed three tumors had Myc amplification. By dual color FISH, one of these three tumors had two populations of tumor cells with the coexistence of amplified Myc and ERBB2, and Myc and EGFR. The second tumor had a high level of amplification of Myc without amplification of ERBB2 or EGFR. The third tumor had a low level of coamplification of Myc and ERBB2. A case of polysomy 8 also demonstrated amplification of ERBB2 and EGFR. Conclusion: The present study shows that genomic instability due to Myc amplification may cause specific amplification of EGFR and/or ERBB2.
- Gallbladder
- adenocarcinoma
- fluorescence in situ hybridization
- Myc
- gene amplification
- ERBB2
- EGFR
Gene amplification is an important genetic change found in cancer cells. Myc, ERBB2 and EGFR are some of the representative protooncogenes that function as oncogenes mainly by gene amplification. Myc located on 8q24 encodes a transcription factor which promotes cell growth and proliferation, inhibits terminal differentiation and sensitizes cells to apoptosis. It has been suggested that Myc can promote genome instability through gene amplification, gene rearrangement and karyotypic instability (1). ERBB2 and EGFR, which are located at chromosome bands 17q12-q21 and 7p12, encode plasma membrane glycoproteins. They are members of the family of tyrosine kinase growth factor receptors (TKGFR) (2). Previous studies using combined immunohistochemistry (IHC) and fluorescence in situ hybridization (FISH) have shown ERBB2 and EGFR to be mainly overexpressed by gene amplification, especially high level amplification, in various solid tumors such as breast (3), stomach (4, 5), colon (6), esophagus (7), lungs (8, 9) and biliary tract carcinomas (10).
Epithelial cancer frequently has multiple amplifications and particular amplicons involving nonsyntenic genes tend to occur together. Among the combinations of coamplified genes, Myc and genes coding TKGFRs such as ERBB2 or EGFR have attracted much attention for their relevance to cytogenetics, carcinogenesis and cancer therapy. The coamplification of these genes was previously examined in gastric cancer and was frequently amplified; surprisingly, it was found that the coamplification of Myc and ERBB2 or EGFR was not incidental (11).
Recently, Paterson et al. examined the physical structure of coamplification of regions 8p12 and 11q13 in cancer cell lines and found a variety of arrangements and concluded that the mechanisms producing coamplification of non-syntenic oncogenes are not simple (12). This encouraged us to perform an examination of Myc amplification in gallbladder cancer, which had demonstrated ERBB2 and EGFR to each be amplified by 10% (10).
Materials and Methods
Tissue samples. A total of 97 gallbladder carcinomas obtained from consecutive surgeries performed at the Department of Surgery, Yamanashi Medical University, between 1986 and 2006 were examined. The patients included 26 males and 71 females ranging from 31 to 89 years of age (mean age, 66.0 years). The conditions of the patients were assessed according to the system for staging primary tumor/regional lymph nodes/distant metastasis (TNM) described in the AJCC Cancer Staging Manual (13) and histological classification according to The World Health Organization Classification of Tumors (14). The 97 patients were classified using the TNM classification system as follows: Stage I, 16 patents; Stage II, 21 patients; Stage III, 21 patients; and Stage IV, 39 patients. Histologically, the specimens were classified into 71 cases of tubular adenocarcinoma (wel-differentiated in 33 cases, moderately differentiated in 24 cases and poorly differentiated in 14 cases), 5 cases of papillary carcinomas and one case of mucinous carcinoma. This laboratory study was approved by the Institutional Review Board at the University of Yamanashi and written informed consent was obtained from all patients.
IHC. Serial sections (4 μm) cut from representative formalin-fixed, paraffin-embedded tissues placed onto MAS™-coated slides were used for hematoxylin-eosin staining, IHC detection and FISH analysis. IHC detection of Myc was carried out on all 97 primary tumors. The expression of ERBB2 and EGFR protein by IHC and their gene amplification by FISH had been previously examined in 89 of these cases and are described elsewhere (10).
A rabbit polyclonal antibody against human Myc (Santa Cruz Biotechnology, Inc, San Diego, CA USA; working dilution, 1:30), a polyclonal antibody against the internal domain of the human ERBB2 protein (Nichirei, Tokyo, Japan; working dilution, 1:100) and a monoclonal antibody against the internal domain of human EGFR (Novocastra Lab, Newcastle, UK; Working dilution, 1:20) were used. For the detection of Myc and EGFR, a high-temperature antigen unmasking technique was used.
The IHC analyses were reviewed by two examiners (SS, KN) who were unaware of the gene amplification data. For an evaluation of Myc staining, only nuclear immunostaining significantly higher than that of the control cells of the normal gallbladder mucosa were considered to be positive. Positive staining was further classified as 2+ and 1+ according to the intensities. For the evaluation of ERBB2 and EGFR positivity, each tumor, or portion of tumor, was scored according to the criteria recommended by Dako (Glostrup, Denmark) for the HercepTest, except that the number of positive cells was not taken into consideration.
FISH. A FISH analysis of Myc amplification was applied to all primary tumors. Primary tumors with positive immunohistochemical staining (2+ and 3+) for either ERBB2 or EGFR and those with Myc amplification were analyzed for ERBB2/EGFR amplification using FISH.
For FISH probes, P1 artificial chromosome clone RP1-80K22 specific for Myc and bacterial artificial chromosomes RP11-62N23 and RP11-339F13 specific for ERBB2 and EGFR, respectively, were used after labeling with SpectrumOrange™ using a nick translation kit (Vysis, Downers Grove, IL, USA). To detect gene amplification, a SpectrumGreen™-labeled pericentromeric probe (Vysis), which was specific to the chromosome on which the gene was located, was cohybridized in order to standardize the chromosome number. For the simultaneous hybridization of Myc/ERBB2, Myc/EGFR and ERBB2/EGFR, a Myc probe and an ERBB2 probe labeled with SpectrumGreen™ were also used.
Amplification was defined as the presence in >60% of tumor cells in a visual field (using a ×40 magnification objective) of more than two times as many gene signals than centromere signals of the respective chromosome. When either >9 gene signals or tight clusters of at least 10 gene signals were observed, the amplification was considered to be at a high level.
Statistical analysis. Agreement among observers in the interpretation of immunostaining of Myc was evaluated by kappa (κ) statistics as described elsewhere (15). A chi-square for independence test was employed to study the relationship between gene amplification and protein overexpression of Myc and the molecular features of Myc and clinical stages. The frequency of coamplification observed was compared with the frequency expected by chance using the Fisher exact test.
Results
Status of ERBB2 and EGFR. An overexpression of ERBB2 and EGFR was found in 14 and 11 out of a total of 97 cases, respectively. In nine of 14 ERBB2-overexpressing tumors and in nine of 11 EGFR-overexpressing tumors, the respective gene amplification was observed by FISH analysis (10). FISH images of high level gene amplification appeared as two distinct types: large clustered signals (LC) and multiple scattered signals (MS) as reported in previous FISH studies (6, 10, 11).
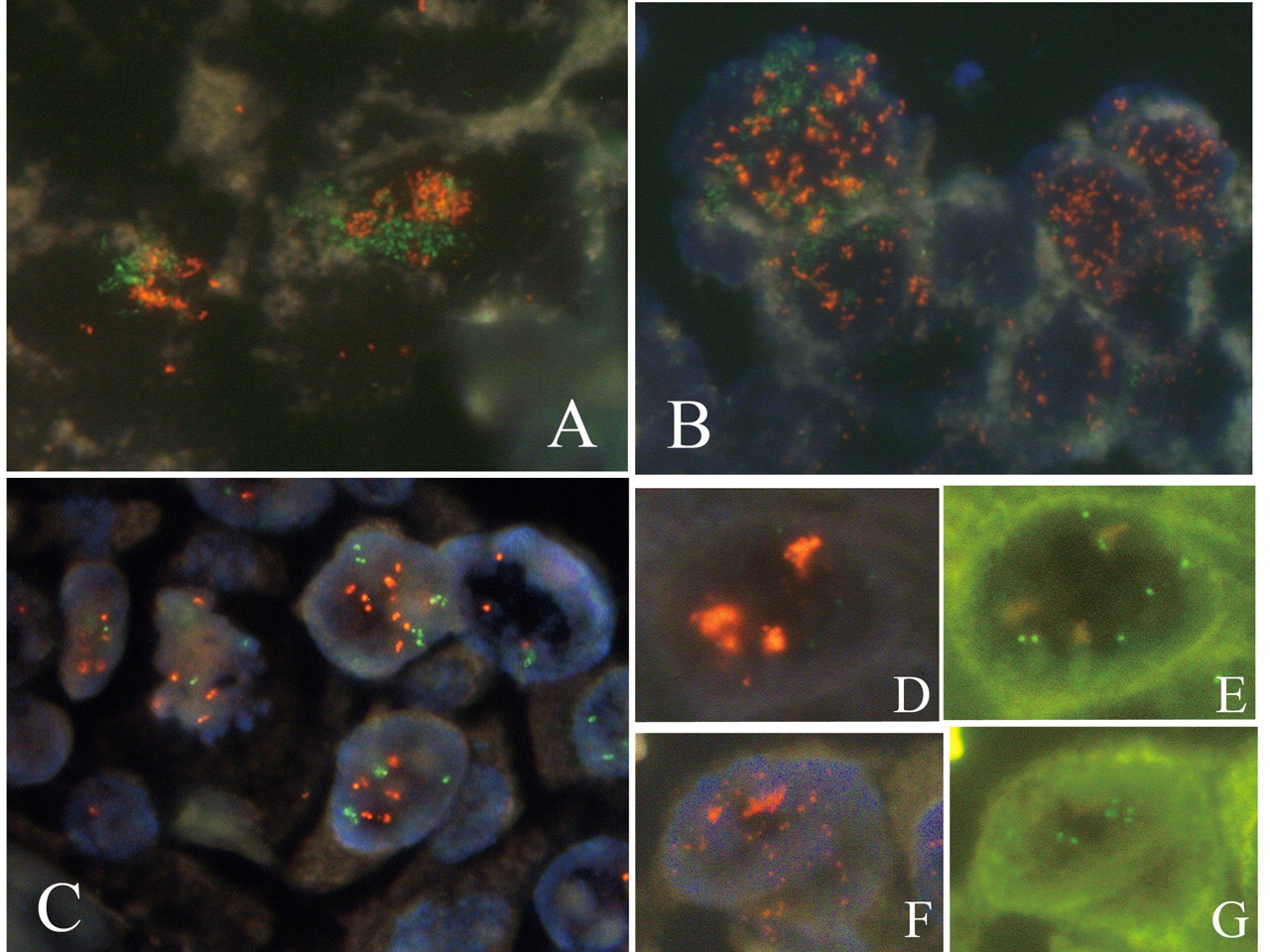
Coamplification, and therefore the co-overexpression of ERBB2 and EGFR in a single tumor was confirmed in two cases. Case 1 was a fungating tumor, with invasion limited to the mucosa. Immunostaining of ERBB2 and EGFR was demonstrated in the cell membranes in approximately 30% and 10% of tumor cell populations, respectively. Occasionally, immunoreactive cells for each were found to be intermingled as shown in Figure 1A and B; however, ERBB2 and EGFR were not found to be co-overexpressed in single cells. The amplifications of ERBB2 and EGFR, which precisely corresponded to protein overexpression on a cell by cell basis, appeared as LC and MS types, respectively (Figure 1C). Case 2 also had coamplification of ERBB2 and EGFR in the LC and MS type respectively. However, each of the amplified cell types was found to occupy separate areas of the tumor.
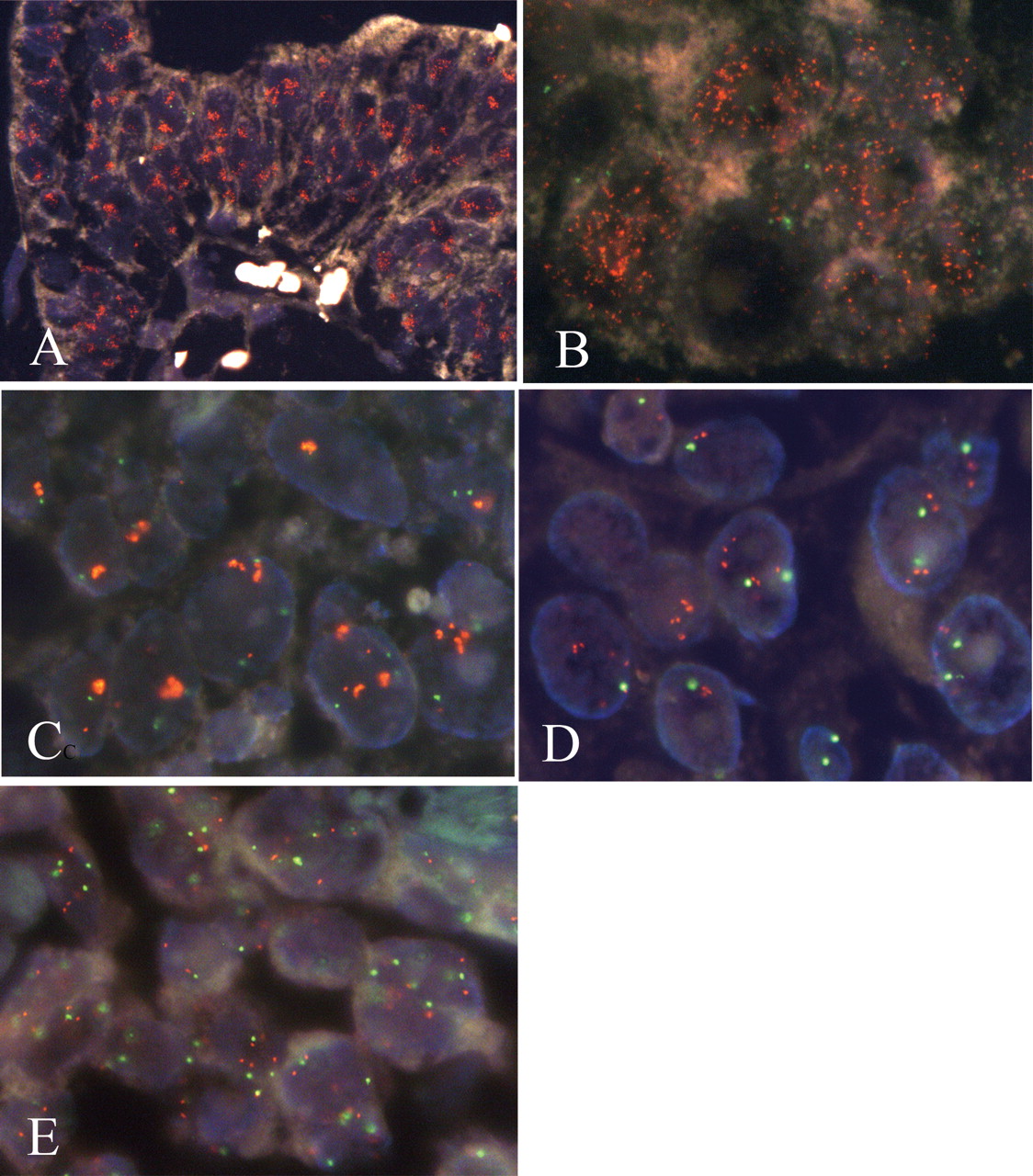
Status of Myc. The interobserver agreement of IHC of Myc was ‘fair’ (κ=0.36). The frequencies of the positive overexpression examined by two observers were 42% and 53% . FISH analysis of Myc amplification was applied to all cases. Successful probe hybridization was achieved in 81 out of 97 cases (84%). Gene amplification of Myc was found in three (3.7%) tumors and their clinicopathological data are shown in Table I. Case 1 showed 2+ immunostaining of Myc, in which distinct nuclear immunostaining of Myc protein was found in the tumor cells. FISH revealed that Myc was amplified predominantly by LC but some by MS as shown in Figure 2A and B, respectively. In the second tumor (case 3), cancer nuclei showed 1+ immunostaining of Myc. According to the FISH analysis, approximately 50% of the tumor cells had several clustered signals of Myc as shown in Figure 2C. The remaining tumor nuclei showed one or two copies of Myc and of centromere 8. Case 4 was another case with Myc amplification in which FISH showed from 1-7 additional Myc signals in comparison to the number of centromere 8 signals as shown in Figure 2D. An increased copy number of Myc was observed in the other 22 tumors; however, they also had the same number of centromere 8 signals, therefore, they were considered to demonstrate polysomy 8. In all cases with polysomy 8, the number of Myc genes was less than five, except for case 2 in which 6-9 Myc signals were found (Figure 2E). In 56 cases, no more than two signals of Myc were found in the tumor cell populations and thus they were considered to be disomies of chromosome 8.

IHC on serial sections alternately stained for ERBB2 (A) and EGFR (B), and dual-color FISH (C) of ERBB2 and EGFR (case 1). ERBB2-positive cells were negative for EGFR and EGFR-positive cells were negative for ERBB2, even though they were situated close together. Cancer nuclei with ERBB2 amplification were identified as clustered green signals and cells with EGFR amplification as multiple scattered orange signals.
Pathological details and results of IHC and FISH analysis of 3 tumors with Myc amplification.

Results of dual-color FISH of Myc (orange signals) and centromere 8 (green signals). Myc is amplified as clustered signals in A (case 1) and C (case 3), numerous scattered signals in B (case 1), or a moderately increased number of signals in D (case 4) and E (case 2). In case 2 (E), a similar number of centromere 8 signals in comparison with Myc signals are visible in most nuclei.
Coamplification of Myc and ERBB2 or EGFR. In case 1, dual-color FISH demonstrated the coexistence of amplified genes in individual nuclei for ERBB2 and Myc, and EGFR and Myc. Amplified signals representing the Myc and ERBB2 genes formed LC opposed to each other as shown in Figure 3A, and amplified Myc and EGFR were found as MS in the nuclei as shown in Figure 3B. In both the LC and MS, however, the overlapping signals of coamplified genes were only incidental. In case 4, no close association of amplified signals of Myc and ERBB2 were observed in dual-color FISH (Figure 3C). In case 2, cancer nuclei with polysomy 8 showed amplified ERBB2 and EGFR in LC and MS form respectively as shown in Figure 3D-G.
{kind=link}
{kind=link}
{kind=link}
Results of dual-color FISH. Cancer cells with coamplification of ERBB2 (orange signals) and Myc (green signals) in LC (A) and those with coamplification of EGFR (orange signals) and Myc (green signals) in MS (B) were found in case 1. In all nuclei, green signals and orange signals only occasionally overlap. Fewer than ten signals of ERBB2 (orange) and Myc (green) are visible scattered in the nuclei in case 4 (C). LC of ERBB2 (D) and MS of EGFR (F) with increased number of Myc (E and G) were found in case 2. (D and F, triple-band filter; E and G, SpecrumGreen™-specific filter).
Statistics. Although all three tumors with Myc amplification had protein overexpression, Myc amplification and/or polysomy 8 were not statistically related to the overexpression of Myc (data not shown). There was no correlation between the overexpression of Myc protein and the pathological stages (data not shown). The expected numbers of cases with coamplification of Myc/ERBB2/EGFR, Myc/ERBB2 and Myc/EGFR were 0.026, 0.28 and 0.28, respectively, whereas the observed number of cases with coamplification were 1, 2 and 1, resepectively. The differences between the expected and observed values were not significant; however, this does not necessarily prove that they are incidental.
Discussion
The present study identified very interesting cases, especially case 1, with regard to the cytogenetics and combinations of amplified genes. Although FISH has limited resolving power on a tissue section, it demonstrated clonal heterogeneity of gene amplification within single tumors, a combination of the coamplified oncogenes and the intranuclear topologies of the amplified genes. This suggested several possibilities concerning the cytogenetic mechanism of gene amplification, especially the coamplification of nonsyntenic oncogenes.
Firstly, in mammalian cells, high-level amplification is detected in two major forms of metaphase structures: homogeneously staining regions (HSR) and centromere-free extrachromosomal double minute chromosomes (DM) (16). An analysis of interphase nuclei with FISH shows that the amplified gene of interest in HSR appears as LC, while those in DMs appear as MS (17, 18). As a result, the MS and LC signals found in this study would usually, although not exclusively, correspond to the amplified genes in DM and HSR, respectively. Therefore, Myc and ERBB2 appear to be amplified in HSR, and Myc and EGFR in DM in case 1, Myc in HSR in case 3, and ERBB2 and EGFR in HSR and DM, respectively, in case 2.
Secondly, the cytogenetic mechanism leading to non-syntenic gene coamplification remains to be clarified. In a previous study on gastric cancer in which coamplification of Myc and ERBB2 or EGFR were observed, a case in which numerous overlapped FISH signals of Myc and ERBB2 were scattered in almost all amplification-positive cancer cells was identified (11). Therefore, the coamplification of non-syntenic genes could occur through translocational junction and subsequent rearrangement. In the present study, however, the coamplification of Myc and ERBB2, or EGFR found in case 1 are hard to explain as the result of amplification of pre-existing translocation junctions. This is because (i) Myc amplification-positive populations contained both amplified and non-amplified subpopulations of ERBB2 or EGFR; (ii) in nuclei with coamplified Myc and ERBB2 or Myc and EGFR, the distribution of the coamplified signals did not overlap; and (iii) Myc was amplified both in the HSR type and DM type structures, although the coamplification of ERBB2 was exclusively associated with the HSR type and EGFR with the DM type. If translocational fusion of Myc and ERBB2 or of Myc and EGFR preceded amplification, then coamplification of Myc and ERBB2 in DM and coamplification of Myc and EGFR in HSR should have appeared. Alternatively, Myc may have been amplified, yielding two clones with HSR and DM and then each clone acquired secondary amplification of ERBB2 and EGFR, respectively. However, it is still unknown why subsequent amplification of ERBB2 and EGFR occurred faithfully in the same structure as Myc amplification.
Recently, Paterson et al. examined the physical structure of the coamplification of regions 8p12 and 11q13 in five breast cancer cell lines using dual-color FISH with many BAC probes. These regions are frequently coamplified in breast cancer. They found a variety of rearrangements: 8p12 and 11q13 fragments overlapping in the same location of the chromosomes in two cell lines, entirely independent amplification of the two regions on separate chromosomes in one cell line and separate amplification of 8p and 11q sequences in distinct sites on the same rearranged chromosome in two lines. They concluded that in the latter three cell lines, the coamplification of 8p and 11q most likely reflects the selection of clones that have amplified both foci (12). They also analyzed the coamplification frequencies of representative genes on 8p12 and 11q13 of 171 breast tumors using a CGH array. Most of the associations of amplified genes on 8p12 and 11q13 reached statistical significance; however, the number of cases with a gain for both loci was only about 1.5-2 times that expected by chance. Therefore, they concluded that the results were consistent with the coamplification arising by selection rather than as a direct result of prior fusions. Unfortunately, in the present study, due to the small size of the cohort, the statistical analysis was not definitive in determining whether the amplification of Myc and ERBB2 and Myc and EGFR were incidental.
The combination of the coamplified genes also seems to support the mechanism suggested above. Cancer progression occurs in a process in which different clones or subclones are produced by genetic instability and at the same time are subject to selective forces. One of the cellular effects caused by the action of Myc is the destabilization of the cellular genome (19), which could facilitate the accumulation of subsequent genetic or chromosomal aberrations. It is also likely that coamplification of Myc and either ERBB2 or EGFR, may result in a growth advantage. In case 1 of the current study, Myc, ERBB2 and EGFR were overexpressed; however, these are the results of gene amplification and there is probably no benefit that a possible translocation preceding the amplification would have given to the tumor. Being coexpressed with Myc, both EGFR and ERBB2 were found to be amplified in mutually exclusive cancer cells in case 1. This observation could be explained by ERBB2 and EGFR sharing the same downstream signal transduction systems and therefore the action of the two molecules might be functionally redundant.
Case 4 had low level amplification of Myc and case 2 showed polysomy of chromosome 8 on which Myc is located. Although there are fundamental cytogenetic differences between gene amplification and polysomy, the copy numbers of Myc genes per cell in these two tumors were similar. There are some clinical data showing that the gene copy number, irrespective of amplification or aneusomy, is associated with the biological behavior of tumor cells. For example, in non-small cell lung carcinomas, the copy number of EGFR genes per cell significantly correlates with the levels of EGFR protein expression (20). Furthermore, copy numbers are important in predicting overall survival following tyrosine kinase inhibitor therapy in patients with advanced carcinoma (21). Reportedly, in breast cancer an increased copy number of Myc genes is significantly associated with poorer outcomes (22). It is possible that in cases 4 and 2, a highly increased number of Myc genes may have induced gene instability, which caused associated gene amplification of ERBB2 and ERBB2 and EGFR, respectively.
Coamplification of Myc and ERBB2 has been extensively examined in breast cancer; however, its frequency still remains controversial (23-25). Interestingly, while patients with breast cancer with coamplification of Myc and ERBB2 had worse outcomes when treated with chemotherapy alone, the addition of monoclonal antibodies against ERBB2 reversed this trend. This has been suggested to be due to cancer apoptosis induced by Myc because the antiapoptotic signals from ERBB2 are blocked (26).
This is apparently the first FISH study examining Myc amplification in gallbladder cancer. Myc amplification was found in only 3.7% of the gallbladder cancer cases but this information might be useful for “tailored” target therapy to Myc in the future. In addition, a genetic analysis using DNA or cDNA expression microarrays may be very convenient; however, caution should be exercised in the interpretation of the data from studies assessing gene amplification using “grind and bind” methodologies that fail to preserve the topology. In case 1, if the methods based on DNA extraction had been applied, then neither the specific combinations of Myc and ERBB2 or EGFR, nor the mutually exclusive amplification of EGFR and ERBB2, would have been clarified.
Acknowledgements
We are grateful to Dr. Johji Inazawa, Department of Molecular Cytogenetics, Medical Research Institute, Tokyo Medical and Dental University, for his helpful discussions.
Grant Supports: The Japanese Ministry of Education, Sports, Science and Culture, Grants-in Aid for Scientic Research C 19590342 (AO) and C 18590327 (YD) and for Young Scientists B 19790256 (SS)
Footnotes
-
Conflict of Interest Statement
We declare that we have no conflict of interest.
- Received June 13, 2008.
- Revision received November 20, 2008.
- Accepted November 27, 2008.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved