


Abstract
Background/Aim: Tumor suppressive microRNAs (miR) are frequently down-regulated during cancer development. The application of synthetic miR molecules restoring suppressed miR, therefore, opens up innovative possibilities in future anticancer therapy. The potential application, however, is limited by the instability of RNA molecules. The presented proof-of-principle study evaluates the potential of using synthetic chemically modified miR molecules as anticancer drugs. Materials and Methods: Chemically synthesized miR-1 molecules containing two 2′-O-RNA modifications, 2′-O-methyl- and 2′-fluoro-derivatives, introduced at different positions of the 3′-terminus, were transfected into prostate cancer (PC) cells (LNCaP, PC-3). Detectability was measured by quantitative RT-PCR. The effect of modifications regarding the growth inhibitory activity of miR-1 was investigated by cell growth kinetics with transfected PC cells. Results: All variants of synthetic modified miR-1 could be transfected into PC cells and were detectable by RT-PCR. Depending on the chemical modification, but especially on the position of the modification, the growth inhibitory activity of synthetic modified miR-1 was increased compared to synthetic unmodified miR-1. Conclusion: Synthetic miR-1 can be enhanced in its biological activity by modification of the C2′-OH group. This depends on the chemical substituent, the position and number of substituted nucleotides. The molecular fine-tuning of tumor suppressive miR like miR-1 may represent a promising approach for the development of multi-targeting nucleic acid-based drugs for cancer therapy.
- Prostate cancer
- cancer therapy
- chemically modified microRNA
- nucleic acid-based drug
- cell growth inhibition
MicroRNAs (miR) have been recognized as a class of regulatory RNA molecules that are frequently dysregulated in cancer cells. During cancer development, miR can function as oncogenes or tumor suppressors, which renders them interesting for therapeutic approaches. While administration of antagonistic miR targeting oncogenic miR conceptually corresponds to the application of pharmaceutical inhibitors (“loss” of miR functions), the restoration of down-regulated tumor suppressive miR by a specific miR substitution therapy has been little characterized so far (“gain” of miR functions) (1, 2).
Application of nucleic acid-based drugs opens up numerous possibilities in anticancer therapy. Apatorsen (OGX-427, OncoGenex Pharmaceuticals, Bothell, WA, USA), a 2′-methoxyethyl-modified antisense DNA oligonucleotide targeting the oncogenic factor heat shock protein 27, is one promising candidate of chemically modified nucleic acid-based drugs and is currently tested in several clinical trials in advanced cancer of the prostate, ovary, breast, bladder, and lung (3). Despite experimental data stating a good level of efficacy for numerous antisense oligonucleotide drug candidates in recent years, biological efficacy often has remained unproven in clinical trials and development has been terminated. A clear limitation of antisense technology is that antisense oligonucleotides specifically bind to one target mRNA only, and thus are more vulnerable to bypassing effector pathways and resistance mechanisms (4-6).
In 2005, Tsuda et al. performed a proof-of-concept study demonstrating that transfected chemically synthesized RNA molecules lead to an attenuation of ovarian cancer cell growth (7). The synthetic antisense RNA was directed against one individual target mRNA, however, this approach opened up the development of miR mimicking RNA molecules with the capacity to regulate several hundreds of genes. Since synthetic miR carry the same sequence as the endogenous miR, it is expected, that they target the same set of genes and control the same set of cellular responses (8). An important precondition for synthetic miR’s increased efficacy compared to antisense oligonucleotides is the introduction of miR molecules into the mammalian RNA interference (RNAi) pathway. Endogenous expression of miR includes the processing of double-stranded primary miR to precursor miR and miR/miR* duplexes, resulting in mature single-stranded miR interfering with protein factors of the RNA-induced silencing complex (RISC). Recent studies have shown that both double-stranded and single-stranded miR are incorporated into the RISC, however, single-stranded miR with lower efficacy (9, 10). In case of single-stranded RNA molecules transfected into cells, antisense as well as RNAi mechanisms have to be taken into consideration when evaluating the biological effects.
Apart from problems with uptake efficacy and possible off target effects, the poor in vivo stability of synthetic RNAs is a particular challenge in the application of oligonucleotide therapeutics. Therefore, chemical modification strategies have been developed to counteract cellular degradation and enhance the in vivo life time of the oligonucleotide drug. In addition, chemical modifications have shown to affect the binding affinity to the target. There is a plethora of chemical modifications to oligonucleotides that can be classified in three categories: (i) internucleotide modifications, (ii) sugar modifications, or (iii) nucleobase modifications. Among a large number of sugar modifications, the 2′-O-methyl (2′-OMe) nucleoside analog is a widely used modification that has been shown to increase nuclease stability and to enhance binding affinity to the target (11, 12). 2′-OMe modified oligonucleotides are well tolerated in siRNA (13, 14), and have been used in the first FDA-approved aptamer Macugen (14, 15). 2′-fluoro (2′-F)-RNA is equally popular as oligonucleotide drug, increasing the binding affinity to the target (16). Resistance against exonucleases apparently is not significantly increased, although 2′-F-RNA imparts resistance towards endonucleases (17). Like 2′-OMe, 2′-F-RNA modifications are well tolerated in siRNA and are components of the FDA-approved aptamer Macugen (15). Furthermore, fully 2′-OMe/2′-F-modified siRNA have been found to be both highly stable and more potent than standard siRNA (18).
In addition to the nature of modification, also the position in the oligonucleotide sequence requires consideration, when designing oligonucleotide therapeutics. The seed region (pos. 2-8) of the antisense strand of an siRNA guides the initial target recognition by the RISC, and modifications in this region would influence activity (19). This would apply similarly to target recognition by miR, such that modifications in the seed region may be risky. In general, modifications must take into account functional regions and should be preferentially restricted to peripheral regions. Accordingly, a number of modifications have been found to be well tolerated in the 3′-region of siRNA or miR mimics (20).
Regarding chemically modified miR for putative miR substitution therapy, the tumor suppressive miR-143 and miR-145, which belong to the same miR cluster, were examined more closely. miR within a cluster are regulated in parallel and control common as well as unique mRNA targets within several cellular pathways (21). After transfection of chemically modified miR-143 and miR-145 duplexes, cellular growth has been inhibited and radiosensitivity of the cancer cells has been restored (22, 23).
Anti-oncogenic miR-1 exhibits pivotal significance in tumor suppression as well and is frequently suppressed in cancer cells during malignant progression. miR-1’s activity has been attributed to activation of apoptosis and cell-cycle arrest as well as to inhibition of proliferation, angiogenesis, chemoresistance, and metastasis (24, 25). Moreover, miR-1 is an inhibitor of androgen receptor expression and activity (26, 27), and thus therapeutic modulation of miR-1 is basically specific for PC cells.
Molecular fine-tuning of tumor suppressive miR may represent a promising approach for the development of multi-targeting nucleic acid-based drugs for future therapy of PC as well as other entities. The aim of the study presented herein was to compare two 2′-O-RNA modifications, F and OMe, introduced at different positions of the 3′-terminus of chemically synthesized miR-1 regarding their impact on miR-1’s cell growth inhibitory properties.
Materials and Methods
RNA synthesis. Oligoribonucleotides miR-1syn and miR-1mod1 to miR-1mod10 and the non-modified nonsense RNA nmR (5′-UUCCGGAGCCGACAGUACAGUCUGGAUGGGAGAAGAUG-3′) were synthesized by the phosphoramidite method on a Pharmacia Gene Assembler Special (Uppsala, Sweden) at 1 μmol scale. The standard 5′-O-DMT-2′-O-TBDMS-P-cyanoethyl protected phosphoramidites adenosine (N-bz), cytidine (N-ac), guanosine (N-ibu) and uridine were obtained from ChemGenes (Wilmington, MA, USA). Also, the CPG-support 5′-O-DMT-2′-O-TBDMS uridine (1,000 A) and the 5′-O-DMT-P-β-cyanoethyl protected modified phosphoramidites 2′-methoxy-adenosine (N-bz) (Figure 1A), 2′-methoxy-uridine (Figure 1B), 2′-fluoro adenosine (N-bz) (Figure 1C) and 2′-fluoro-uridine (Figure 1D) were obtained from ChemGenes. CPG-support 5′-O-DMT-2′-O-methyl uridine (1000 A) was obtained from LGC Link (Bellshill, UK). 1-(Benzylmercapto)-1H-tetrazole (BMT, Biotech, Berlin, Germany) was used as activator. The water content of acetonitrile in coupling solutions was lower than 10 ppm (ACROS Organics, Geel, Belgium). The solutions of phosphoramidites, activator (BMT) and acetonitrile (MeCN) were kept over molecular sieve 0.3 nm. The coupling time for all natural and modified building blocks was 5 min. All syntheses were carried out “trityl-off”.
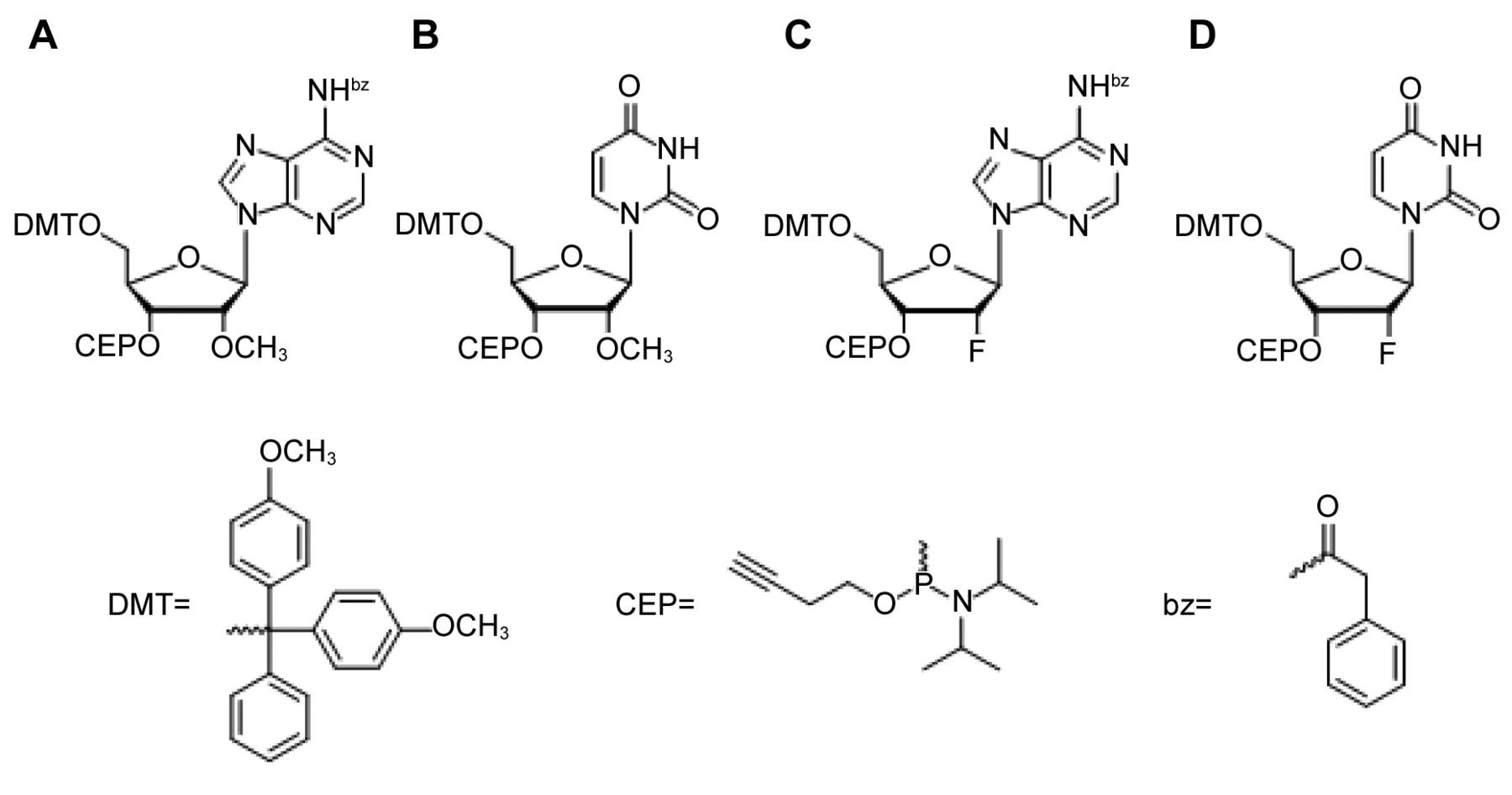
2′-Methoxy- and 2′-fluoro modified phosphoramidites used for chemical solid phase synthesis of oligoribonucleotides miR-1syn and miR-1mod1 to miR-1mod10 by the phosphoramidite method on a Pharmacia Gene Assembler Special (Uppsala, Sweden) at 1 μmol scale in the trityl-off mode.
The phosphoramidites, obtained from ChemGenes and LGC Link, were used as 0.1 M solution in acetonitrile for natural nucleosides and as a 0.15 M solution in acetonitrile for 2′-modified building blocks. The coupling time for all amidites was 5 min as described in the synthesis protocol (Table I).
Conditions for solid phase synthesis of miR-1syn, miR-1mod1 to miR-1mod10 and the nonsense-RNA nmR.
RNA deprotection and purification. The obtained RNA was cleaved from the support by incubation with a 1:1 mixture of concentrated ammonia and an ethanol-solution of methylamine (8.0 M) (Merck Millipore, Billerica, MA, USA) for 40 min at 65 °C in two 2 ml screw vials. Simultaneously the N-protecting- and the β-cyanoethyl-groups were cleaved off. After washing the support with ice-cold ethanol all supernatants were collected and lyophilized. The 2′-tert-butyldimethylsilyl protecting groups were removed by incubation of the obtained oligomer for 1.5 h at 55 °C with a mixture of triethylamine trihydrofluoride and dimethylformamide (3:1, v/v). The reaction was stopped by adding an aqueous ethanol solution (25%). RNA was precipitated with butanol and purified by polyacrylamide gel electrophoresis (PAGE) on a 15% denaturating polyacrylamide gel. RNAs were obtained by elution from the gel with 0.3 M sodium acetate (pH 7.0), followed by ethanol precipitation. RNA concentration was determined using the NanoDrop spectrophotometer (NanoDrop ND-1000; NanoDrop Technologies, Montchanin, DE, USA) at 260 nm and RNA samples were stored as solid at −20°C.
Cell culture. The human PC cell lines LNCaP and PC-3, all received from Cell Lines Service (CLS), Eppelheim, Germany, were propagated in RPMI 1640 medium supplemented with 10% fetal bovine serum, 1% pyruvate and 100 units/ml penicillin/streptomycin (all PAN Biotech, Aidenbach, Germany) at 37°C and 5% CO2 in a humidified atmosphere. Cells were passaged twice per week.
Transfection experiments. Electroporation was performed with an ECM 630 Electro Cell Manipulator and a 630B Safety Stand (BTX Harvard Apparatus, Holliston, MA, USA). 3.5×106 cells were suspended in 550 μl electroporation buffer (21 mM HEPES, 137 mM NaCl, 5 mM KCl, 0.7 mM Na2HPO4, 6 mM Glucose, pH 7.15) containing 2.5 μg of chemically synthesized miR species. Electroporation was performed in a 0.4 cm cuvette at 300 V and 1,500 μF followed by the complementation with 550 μl fetal bovine serum (PAN Biotech) and further incubation at 37°C and 5% CO2 in a humidified atmosphere. Control transfections were carried out using the nonsense RNA nmR.
RNA preparation. After incubation of transfected cells, total RNA was extracted using 500 μl peqGold TriFast reagent (PeqLab, Erlangen, Germany) according to the manufacturer’s instructions. Briefly, cells were lysed by 1.0 ml Trizol reagent, homogenized, and incubated for 5 min at room temperature. After addition of 200 μl chloroform, RNA samples were vortexed and centrifuged (12,000 × g). For RNA precipitation, the aqueous phase was transferred to a clean tube and RNA was precipitated by adding 0.5 ml of isopropanol. After a centrifuge step (12,000 × g) precipitated RNA was washed twice with 70 % ethanol, carefully dried and dissolved in 10-30 μl diethylpyrocarbonate treated water. RNA concentration was determined using the NanoDrop spectrophotometer (NanoDrop ND-1000; NanoDrop Technologies) at 260 nm and RNA samples were stored by −80°C.
Quantitative reverse transcription - polymerase chain reaction (RT-PCR). To perform the cDNA synthesis, 100 ng of total RNA were used with Superscript III Reverse Transcriptase (Life Technologies, Darmstadt, Germany) according to the protocol of Chen et al. (28). The required stem-loop primers were designed as follows: miR-1 stem-loop: 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCG CACTGGATACGACATACAT-3′; U6 stem-loop: 5′-GTCATC CTTGCGCAGG-3′.
Following, quantitative PCR was performed with SensiMix SYBR hi-ROX Kit (Bioline, Luckenwalde, Germany) on a CFX96 Real-Time System (Bio-Rad, München, Germany) with CFX Manager software (Bio-Rad). PCR primer sequences were as follows: miR-1 forward: 5′-GCCCGCTGGAATGTAAAGAAGTATG-3′; miR-1 reverse: 5′-GTGCAGGGTCCGAGGT-3′; U6 forward: 5′-CGCTTCGGCAGC ACATATAC-3′; U6 reverse: 5′-AGGGGCCATGCTAATCTTCT-3′. The cycling parameters were one denaturation cycle at 95°C for 5 min and 45 amplification cycles at 95°C for 10 s, 60°C for 20 s, and 72°C for 10 s, followed by a melting-curve analysis. For quantification, miR-1 signals were standardized to U6 RNA as reference and fold change was calculated according to the 2ΔΔCq method.
Cell proliferation assay. Cell growth was determined by using a CASY Cell Counter and Analyzer Model TT (Roche Applied Science, Mannheim, Germany). Adherent cells were treated with trypsin/ethylenediaminetetraacetic acid (PAN Biotech), and cell suspension was diluted in CASYton dilution buffer (1:100; Roche Applied Science). For the automated count of cells, 400 μl of the cell suspension were analyzed in triplicates applying a capillary of 150 μm in diameter and cell line-specific gate settings to discriminate between living cells, dead cells, and cellular debris (LNCaP: 6.0 μm/12.25 μm; PC-3: 7.2 μm/15.45 μm).
Statistics. Results of at least seven independent experiments were statistically analyzed using GraphPad Prism 7.0 (GraphPad, La Jolla, CA, USA). Analysis was performed using the Student’s t-test and significance levels were defined as *p≤0.05 (*), **p≤0.01, and ***p≤0.001. Data are expressed as mean±SD.
Results
A number of miR-1 versions have been synthesized, incorporating one to three modified nucleotides, either 2′-OMe or 2′-F-derivatives (Figure 2), at the 3′-terminus (Table II).

2′-O-methyl (2′-OMe) nucleoside (A) and 2′-fluoro (2′-F) nucleoside (B) analogs in chemically modified miR-1 species.
Name (column 1), sequence, modification, and position of the modification (column 2) of the 10 synthetic miR-1 species. In column 3, growth inhibitory activity of the synthetic miR is divided into inhibits growth stronger than unmodified miR-1syn (+) or has the same or lower growth inhibitory effect as unmodified miR-1syn (−).
Initially, dilutions of synthetic RNAs were analyzed by RT-PCR to determine the detectability and detection limits of modified RNAs. For this purpose, ten-fold serial dilutions ranging from 10−3 M to 10−10 M of the exemplarily selected 2′-OMe-substituted miR-1mod3 were analyzed by RT-PCR and compared to the corresponding dilution series of unmodified miR-1syn (Figure 3). The detection sensitivity was comparable for both synthetic RNA species. At the concentration of 10−3 M, threshold cycles of 23.15 (miR-1syn) and 22.14 (miR-1mod3) were observed. As expected, by diluting the RNA solutions by a factor of ten, the fluorescence signal curves shifted approximately 3.3 threshold cycles to the right. Detection of synthetic and chemically modified miR-1 molecules was feasible over a concentration range from 10−3 M to 10−6 M. At a concentration of 10−6 M or below, quantitative detection of RT-PCR products disappeared primarily by loss of linearity in fluorescence due to low concentrations of fluorescent agents. As a result of this, accurate quantification of synthetic miR-1 molecules was enabled up to concentrations of about 10−6 M.

Determination of detectability and detection limits of modified RNAs by chemically modified miR-1 RNA molecules. Ten-fold serial dilutions (10−3 M to 10−10 M) of unmodified miR-1syn (A) and 2′-OMe-substituted miR-1mod3 (B) were analyzed by quantitative RT-PCR.
The tumor suppressive activities of the 10 synthetic modified miR-1 candidates were tested by proliferation assays on electroporated PC cells. Electroporation efficiency was determined by applying preparations of total cell lysates to a miR-1-specific quantitative RT-PCR procedure. For data analysis, RT-PCR signals were normalized to control cells transfected with synthetic nonsense RNA (control=1.0), thereby eliminating cellular signals from endogenously expressed miR-1 (Figure 4). The relative amounts of modified miR-1 individually varied from 2-fold (miR-1mod8) to 1503-fold (miR-1mod3) compared to mock transfected control cells. Notably, the 2′-OMe-substituted species miR-1mod1 (246-fold) and miR-1mod3 (1503-fold) possessed higher, while the 2′-F-substituted miR-1mod8 (2-fold) and miR-1mod9 (5-fold) possessed lower intracellular levels than unmodified miR-1syn (185-fold).

Relative intracellular levels of miR-1syn, miR-1mod1, miR-1mod3, miR-1mod8, and miR-1mod9 in transfected cells. The corresponding miR were transfected into LNCaP cells, incubated for 24 h and the specific miR concentrations were determined by quantitative RT-PCR. Synthetic nonsense RNA transfected LNCaP cells served as control (Ctrl=1.0).
The tumor biological properties of modified miR-1 molecules were determined by cell growth kinetics over a period of 240 h. PC cells LNCaP and PC-3 were transfected individually with all species from miR-1mod1 to miR-1mod10, and cell growth rates were compared to miR-1syn transfected cells serving as control. The antiproliferative efficacy of unmodified miR-1syn compared to nonsense transfected control cells was similar to biological expressed DNA plasmid-encoded miR-1 which has been demonstrated in all setups with LNCaP and PC-3 cells (26).
To show representative results, one of the highest effects and one of the least effects of 2′-OMe- and 2′-F-substituted synthetic miR in LNCaP cells are depicted in Figure 5 (remaining results are not shown). In LNCaP cells, the 2′-OMe-substituted miR-1mod1 (Figure 5A) revealed no or weak enhancement of miR-1-mediated growth inhibition compared to unmodified miR-1syn. In contrast, 2′-OMe-substituted miR-1mod3 (Figure 5B) produced a significant greater cell growth suppression than unmodified miR-1syn. The 2′-F substituted miR-1 did not show such pronounced variations in the amplification of the growth-inhibiting effect. The slightest anti-proliferative effect of miR-1mod8 (Figure 5C) was almost as pronounced as the strong effect of miR-1mod9 (Figure 5D). These data were confirmed by comparable transfection series with PC-3 cells (data not shown).

Growth inhibitory activity of unmodified miR-1syn (△) and 2′-OMe-substituted miR-1 species (◇) in LNCaP cells compared to synthetic nonsense RNA (□) as control. 2′-OMe-substituted miR-1mod1 (A) and miR-1mod3 (B) as well as 2′-F-substituted miR-1mod8 (C) and miR-1mod9 (D) were incubated for 240 h and cellular growth wass assessed by cell counting. Results are expressed as the mean±SD of cell count. *p<0.05; ***p<0.001, as determined by the Student’s t-test.
Discussion
miR activities regulate cancer development by targeting up to hundreds of mRNA species, thereby controlling central cellular processes such as cell growth, cell motility, and epithelial-to-mesenchymal transition (24, 29, 30). Tumor suppressive miR become frequently down-regulated during cancer progression, and loss of anticancer pathway control enables malignant cell growth. Consequently, restoration of intracellular levels of tumor suppressive miR like miR-1 represents a new and promising concept for molecular cancer therapy (24, 31, 32).
The idea of using synthetic oligonucleotides to control the expression of specific genes dates back to the late 1970s (33, 34). Since then, a vast variety of chemical modifications has been developed, for one to counteract the poor extracellular and intracellular stability of synthetic oligonucleotides, and for second to increase the affinity to a specific target, which could possibly also potentiate biological effects (35).
The correlation of the biological efficacy of modified synthetic siRNA or miR molecules with the chemical nature of the substituent is poorly studied. 2′-OMe and 2′-F substitutions are the most commonly used modifications in synthetic RNA approaches in mammalian cells (11). Both substituents exhibit low toxicity, but 2′-OMe-substituted RNA molecules appear to be more sensitive to RNAse activity and thus more unstable in cells (36, 37). The biological activity of RNA molecules, however, does not seem to be affected by the modification. A complete 2′-F substitution of a green fluorescent protein (GFP)-specific siRNA was not only significantly more stable in the presence of RNAses, but also caused almost the same degradation of the target protein as the unmodified control RNA (18, 38). Sugar modifications by 2′-OMe and 2′-F also occur naturally in tRNA and rRNA in mammals. This suppresses autoimmune responses, which would also be advantageous over other modifications in the therapeutic use of synthetic RNA molecules (39, 40). Complex foreign RNA, e.g., virus RNA, does not have such modifications and usually elicits an immune response. Other substitutions include bicyclic RNA molecules in which the 2′- and 4′-positions of the ribose are linked by a methylene residue (locked nucleic acid; LNA) and RNA molecules with alkylation of the 2′-OH with aminoethyl, guanidinoethyl, cyanoethyl, and allyl groups (18, 41). Studies on these modifications mostly concern the stability of synthetic RNA to enzymatic degradation in cells and blood. However, biological effects have hardly been analyzed so far, which is why we chose 2′-OMe and 2′-F groups as substituents in this experimental study.
We tested a series of chemically synthetized C2′ OMe- and C2′ F-substituted miR-1 molecules regarding their antioncogenic potential to retard PC cell growth. The results provided evidence that modified miR-1 can be administered effectively into cancer cells and harbors antiproliferative effects. In our study, 2′-OMe-substituted miR-1 molecules possessed higher transfection efficiencies than 2′-F-substituted miR-1 molecules. There are little reliable data in the literature on the influence of the chemical nature of the substituent on the transfection efficiency of RNA molecules. Our study shows clear differences in intracellular concentrations of 2′-Me- and 2′-F-substituted miR molecules after transfection. This is not exclusively dependent on the transfection rate but suggests that it is also different. However, the lower concentration of F substituted miR molecules could also be due to their lower stability toward cellular RNAses (42). A general dependence of transfection efficiency on the substituent cannot be postulated. Transfection of differently modified 2′-OMe- and 2′-OMe-4′-thio-substituted siRNA showed little difference in biological efficiency, again indicating very similar transfection rates (43, 44).
Apart from a potentially significant influence of the substituent, the transfection method probably also has an impact on transfection efficiency. Vesicle-based transfections introduce high RNA concentrations into the cell, but these lead to comparatively low biological effects. By vesicle-free transfection, e.g., electroporation, less RNA enters the cell, but the biological effect is comparable (45, 46). We also observed this phenomenon. Modified miR transfected by liposomes caused significantly lower antiproliferative effects than comparable electroporation approaches (data not shown). Presumably, the intracellular release of RNA from the transfection vesicles is also an essential factor.
We found that the antitumor effects of miR-1 significantly increased by chemical modification of the C2′-position of several modified variants compared to unmodified synthetic miR-1syn. Anti-proliferative effects of modified miR-1 ranged from very slight growth-inhibitory effects to strongly reduced cell growth of 155% of the unmodified miR-1syn efficacy (Figure 5D).
In case of miR-1mod5 transfected into LNCaP cells and miR-1mod1, miR-1mod4, miR-1mod5, and miR-1mod7 transfected into PC-3 cells, growth inhibitory effects were less than that of unmodified miR-1syn. With the exception of miR-1mod1 all these miR contained two or three C2′-substituted nucleotides. Comparable cellular effects were described by Noguchi et al. in melanoma cells (47). The authors have observed that the addition of aromatic benzene-pyridine analogs to the 3′ terminus of synthetic miR-205 resulted in a complete loss of tumor suppression functions. These findings may point to a critical issue in the context of RNA based therapy approaches. Inappropriate substituents and substitution positions may neutralize the anticancer effect of tumor suppressive miR.
Comparison of 2′-OMe-substituted (miR-1mod2, miR-1mod3, miR-1mod6) and 2′-F-substituted (miR-1mod8, miR-1mod9, miR-1mod10) miR-1 molecules with C2′ modifications in corresponding positions revealed similar growth inhibitory properties. For the three 2′-F-substituted miR lower intracellular concentrations in transfected cells have been detected. This could be due to a lower transfection efficiency or to the fact that 2′-F-substituted RNA molecules have lower exonuclease stability compared to 2′-OMe-RNAs (17). Nevertheless, cell growth inhibitory effects comparable to those of 2′-OMe miR were achieved with 2′-F-substituted miR which indicate the high antiproliferative effect of tumor suppressive miR such as miR-1. It is possible that the efficiency of biologically active molecules such as microRNA does not depend to the same extent on the concentration as pharmacological inhibitors do.
Interestingly, the present study also demonstrated cell line-dependent differences in LNCaP and PC-3 cells transfected with miR-1mod7, miR-1mod8, and miR-1mod10. In LNCaP cells, these three modified miR caused significantly reduced cell growth compared to unmodified miR-1syn transfected cells, whereas in PC-3 cells only little or no growth attenuation was detectable (Figure 5C). Overall, transfection experiments suggest that LNCaP cells are more sensitive to chemically modified miR-1 than PC-3 cells. Out of the applied 10 chemically modified miR-1 species, seven variants inhibited LNCaP cell growth, but only four variants had a similar effect on PC-3 cells. Presumably due to well-known cellular and molecular differences between both cell lines, e.g. in androgen sensitivity, androgen receptor expression, and p53 status (48, 49), PC-3 cells are more resistant to anticancer treatment than LNCaP cells, as suggested by literature data (50-53). Additionally, one cannot exclude the possibility of varying transfection efficiencies utilizing chemically modified RNA molecules for the transfection of LNCaP and PC-3 cells.
In contrast to similar growth inhibitory properties of 2′-OMe- and 2′-F-substituted miR, our data demonstrate an influence of the substituent positions (Table III), which may further potentiate the miR-1’s anticancer capacity.
Overview of the growth inhibitory activity of the synthetic miR depending on the position of modification and the applied PC cell lines. Classification of the growth inhibitory activity (+ and –) as described in Table II.
Substituents on position 20 and position 21 primarily caused positive effects on modified miR-1 anticancer properties. Substitution of the nucleotide 22, however, failed, with one exception, to enhance miR-1’s anticancer activities. Thus, the terminal 3′-nucleotide may represent a critical nucleotide for the molecular mode of action of miR-1 and an unmodified 3′-ribose residue may appear essential for miR-specific cell growth inhibition. Since recent studies have shown the incorporation of single-stranded miR into the RISC (9, 10), this molecular model may be supported by our findings that nucleotide 22 represents a putative binding site for cellular factors, such as proteins of the RNAi machinery.
Conclusion
Our data demonstrated, for the first time, the feasibility of molecular tuning to enhance anticancer capabilities of tumor suppressive miR-1. Chemically synthesized and modified miR-1 introduced into PC cells enables an increased suppression of tumor cell growth compared to unmodified miR-1 molecules. The anticancer efficacy of modified miR-1 thereby depends on the position rather than the type of the modified nucleotide. Under certain circumstances, inappropriate substitutions may lead to attenuated miR-1 molecules with extenuated anti-proliferative properties. Considering the background of clinical applications, the molecular tuning of synthetic tumor suppressive miR may open up a new class of nucleotide-based drugs for anticancer therapy. By selecting well characterized microRNA possessing a high antiproliferative potential, tailor-made synthetic modified miR could be designed for the future therapy of different types of cancer.
Acknowledgements
The Authors thank Julia Drenckhan for her excellent technical assistance.
Footnotes
Authors’ Contributions
Conceptualization: S.M., M.B.S.; Methodology: B.A., S.M., M.B.S.; Formal Analysis: B.A., S.M., M.B.S.; Investigation: H.A., B.A., L.J.; Writing-Original Draft Preparation: S.M., M.B.S.; Writing-Review & Editing: S.M., M.B.S.; Supervision: S.M., M.B.S.
Conflicts of Interest
The Authors declare that they have no competing interests.
- Received February 18, 2023.
- Revision received March 9, 2023.
- Accepted March 20, 2023.
- Copyright © 2023 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.