


Abstract
Background/Aim: This study evaluated the effect of enrofloxacin antibiotic treatment on the ability of an attenuated Salmonella typhimurium (S. typhimurium) strain VNP20009 to control schwannoma growth in a preclinical mouse schwannoma tumor model. Materials and Methods: The antitumor efficacy of VNP20009 intratumoral (i.t.) injection was assessed in a syngeneic mouse-NF2 schwannoma model, with and without subcutaneous (s.c.) injection of enrofloxacin beginning on day-1 or day-8 post-VNP20009 injection. S. typhimurium colonization was assessed in excised tumor samples following antibiotic treatment. Results: I.t. injection of the VNP20009 significantly decreased the growth of schwannoma tumors in mice compared to PBS-treated controls. Treatment of mice with enrofloxacin on day-1 post-VNP20009 injection resulted in abrogation of VNP20009-mediated tumor growth control. In contrast, tumor growth in i.t. VNP20009-injected mice infused with enrofloxacin beginning on day 8 was significantly decreased compared to i.t. PBS-injected controls. Enrofloxacin significantly reduced the number of viable VNP20009 bacteria in excised tumor samples within one day of antibiotic infusion. Viable bacteria were either few or essentially eliminated at the end of the experiment in antibiotic-treated animals compared to VNP20009-only. Conclusion: Viable VNP20009 can persist for as long as 2.5 weeks following intratumoral injection of schwannoma, during which time tumor growth is retarded. Antibiotic treatment starting 1-day following i.t. VNP20009 abrogated bacterial tumor growth control, whereas initiation of antibiotics 8-days following i.t. VNP20009 was associated with control of tumor growth, albeit less than seen in animals unexposed to antibiotics.
While the potential for bacterial cancer therapy (BCT) has been of clinical interest for many decades (1), only one such therapy, utilizing a live attenuated strain of Mycobacterium bovis for the treatment of high-risk non-muscle-invasive bladder cancer, has been approved by the U.S. Food and Drug Administration (2). Despite many promising results of BCT using attenuated Salmonella typhimurium (S. typhimurium) strains in animal models (3-5), clinical studies have been disappointing (6, 7). One of the challenges is the safe delivery of bacterial inoculum in an amount sufficient to control rapidly dividing cancer cells in an immunosuppressive environment. A recent alternative approach has been to assess bacterial therapy for slow-growing benign tumors, such as schwannomas, which are mainly treated by means of operative resection and symptomatic management of pain because anti-cancer therapeutics generally are ineffective due to the slow replication rates of these tumors (8). The only generally accepted pharmacotherapy, bevacizumab, temporarily stabilizes growth in a subset of schwannomas by targeting the highly vascularized state of these tumors (9, 10). However, being genetically stable, slow growing and highly vascularized, schwannomas may provide a target for bacteria-mediated tumor cytotoxic and anti-angiogenic effects. Schwannomas also contain immunosuppressive type 2 macrophages and regulatory T cells (Tregs) (11), representing an immunological milieu potentially receptive to bacteria-mediated mechanisms that can overcome such a phenotype and thereby induce host anti-tumor immunity. We recently demonstrated that i.t. injection of S. typhimurium strain VNP20009 controls the growth of intrasciatic schwannomas in both a human xenograft and a mouse syngeneic model (12). VNP20009 injection increased immune cell infiltration into tumors, shifted the balance of resident macrophages from the tumorigenic M2 type to the tumoricidal M1 type, and inhibited tumor angiogenesis. VNP20009 was shown to invade tumor cells, elicit pro-immunogenic cytokines and inflammasome proteins, and induce tumor-cell apoptosis. VNP20009 also controlled the growth of uninjected subcutaneous (s.c.) schwannomas contralateral to VNP20009-injected tumors and rechallenge intrasciatic schwannomas (implanted after injection of VNP20009), stimulating the same immune responses in tumor cells as in the prior experiments. Growth of two additional neoplasms, peripheral nerve sheath tumors (MPNSTs) and meningiomas, were also controlled by i.t. VNP20009 injection (12).
The purpose of the current study was to assess whether persistent tumor colonization by VNP20009 is necessary for schwannoma growth control. The relevance of this question is underscored by the failure of VNP20009 to control growth of late-stage malignancies; investigators of these clinical trials hypothesized that the failure of intravenous VNP20009 treatment to control tumor growth may have been due to inadequate tumor colonization (7). In this study, we utilized enrofloxacin treatment starting either 1 or 8-days after intratumoral injection of VNP20009 to test the hypothesis that persistent bacterial colonization is necessary for VNP20009-mediated schwannoma growth control.
Materials and Methods
Tumor cell culture. Mouse 08031-9 schwannoma cells (obtained from Dr. Marco Giovannini, Univ. of California, Los Angeles, CA, USA) were maintained as previously described (13). We confirmed that the 08031-9 schwannoma cell line was free of contaminants, including mycoplasma.
Bacterial cell culture. We purchased the attenuated Salmonella enterica serovar typhimurium strain VNP20009 (with modified lipid A (msbB–) and purine auxotrophic mutation (purI–); ATCC YS1646, cat. #202165, Manassas, Virginia, USA). Bacteria were cultured in Luria-Bertani (LB) broth medium with low sodium (DifcoLaboratories, Detroit, MI, USA) at 37°C and 300 rpm overnight in aerobic conditions (14, 15). Cells were grown to the late-log phase (OD600=0.8), harvested by centrifugation at 5,000 rpm for 10 min, and washed twice with sterile 1× phosphate buffered saline (PBS) before injection into tumors.
Animals. All animal experimentation was approved by and conducted under the oversight of the Massachusetts General Hospital (MGH, Boston, MA, USA) Institutional Animal Care and Use Committee. Animals, FVB/N mice (obtained from Charles River Laboratories headquarters; Wilmington, MA, USA), were kept on a 12:12 light-to-dark cycle with ad libitum access to food and water.
Mouse schwannoma model and intratumoral injection of bacteria. The utilized mouse-schwannoma model was generated by s.c. injection of 08031-8 mouse schwannoma cells into the left flank of isoflurane-anesthetized syngeneic immunocompetent FVB/N mice (5-7-week-old males; Charles River Laboratories), as described (13, 16). We resuspended 08031-9 cells in PBS and mixed with Matrigel (1:1) (BD Biosciences, Bedford, MA, USA); then grafted 105 cells subcutaneously into mice. Tumors were measured with calipers. We estimated tumor volume using the equation W × L × L × π/6, where width (W) and length (L) corresponded to the two largest dimensions, as described (13). Once tumor reached 100 mm3, VNP20009 cells (1×104 cfu/mouse in 50 μl PBS) were injected percutaneously into tumors. Control animals were injected intratumorally with PBS alone. An insulin syringe was used for all i.t. injections of S. typhimurium VNP20009 and PBS.
Enrofloxacin treatment. On day 1 or day 8 post i.t. VNP20009 injection, schwannoma-bearing mice were infused subcutaneously with enrofloxacin (Sigma Aldrich, St. Louis, MO, USA, Cat. #PHR1513) at 5 mg/kg diluted in 100 μl saline every 12 h for 14 days (17).
Assessment of viable S. typhimurium in tumors. Animals were terminally anesthetized with isoflurane (3%) and sacrificed via cervical dislocation in a manner consistent with institutional regulations. Tumors were resected, homogenized, and serially diluted in PBS (10-fold), and plated on plain (LB) plates. After incubating plates overnight at 37°C, S. typhimurium colonies were counted. Dilution and resected tumor weight were included in calculation of the bacterial titer (CFU/g), as previously described (4).
Histological and immunohistochemical analysis. Animals were terminally anesthetized with isoflurane (3%) and sacrificed by cervical dislocation. Tumor tissues were removed, snap frozen, and stored at −80°C for block cutting and immunohistochemical staining. Primary antibodies against bacterial outer membrane protein A (ompA) (Biorbyt, St Louis, MO, USA, cat. #orb422682) were used for staining bacteria. Sections were dried at room temperature (RT) overnight, then fixed in pre-chilled acetone at 4°C for 10 min, allowed to dry, and stained immediately thereafter. Staining involved several steps. First, sections were washed in PBS, blocked with serum-free protein block (Dako, Carpinteria, CA, USA), and quenched for peroxidases in dual endogenous enzyme block (Dako). Next, sections were washed in PBS and incubated with primary antibody for 1 h at RT, washed again in PBS, and incubated with horseradish peroxidase-conjugated secondary antibody (Abcam, Boston, MA, USA, cat. #ab6721) for 30 min at RT. Then, sections were washed in PBS and incubated with 3,3′-diaminobenzidine (DAB) solution (Dako) followed by counterstaining in ethanol and xylene before mounting in Cytoseal (Richard Allan Scientific, San Diego, CA, USA). Finally, sections were covered with cover slips for microscopic visualization.
Data analysis. All group values are presented as mean±standard error of the mean (SEM). We used repeated measures ANOVA to compare outcomes across groups (18) with Bonferroni post hoc corrections for pairwise comparisons. We considered p<0.05 as significant. We utilized Microsoft Excel for statistical analyses.
Results
I.t. injection of attenuated S. typhimurium suppressed tumor growth in a murine syngeneic schwannoma model. As previously described (13, 16, 19), we produced subcutaneous murine schwannomas, utilizing the 08031-9 cell line, that developed in syngeneic immunocompetent (FVB/N) mice. Thirteen days after tumor-cell implantation, schwannomas were injected under direct visualization with attenuated S. typhimurium strain VNP20009 or PBS (control). Tumor growth was assessed over the subsequent 18 days via direct measurement. Replicating our previous findings (12), i.t. injection of VPN20009 resulted in decreased tumor size compared to the control group (Figure 1) by approximately 50% after one week and 66% by the end of the experiment (day 32).
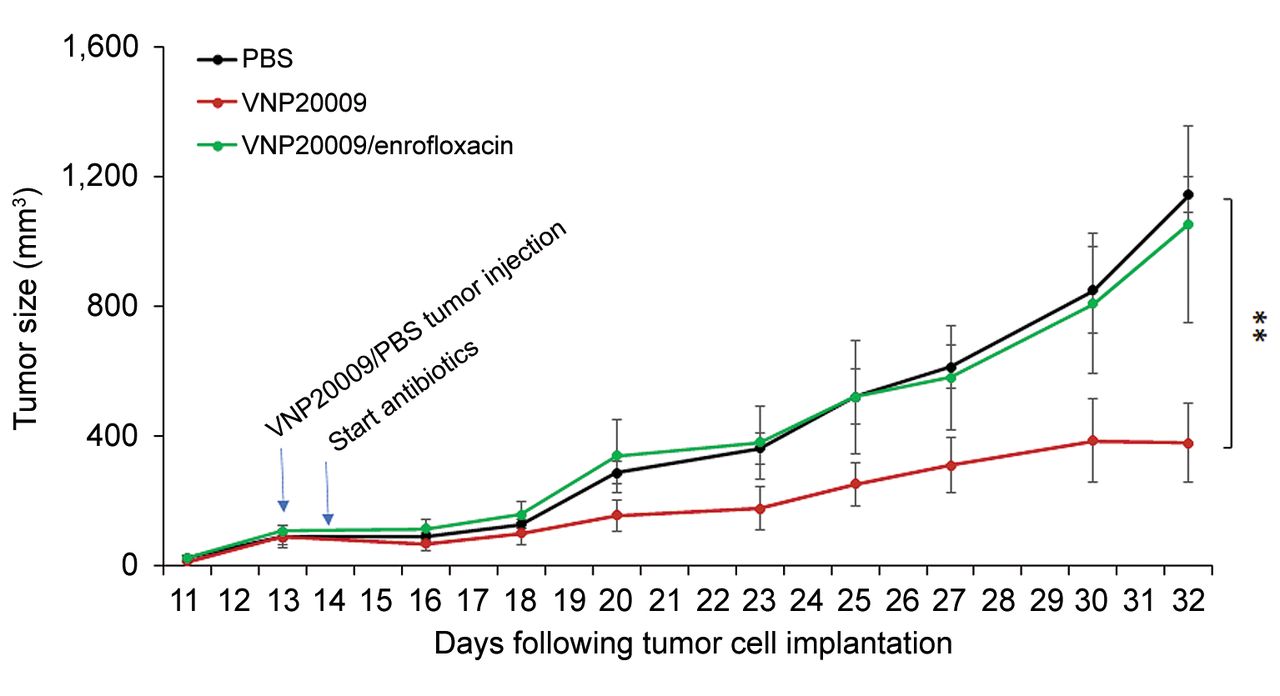
Early infusion of enrofloxacin abrogated S. typhimurium-induced tumor growth retardation of schwannoma tumor development in syngeneic mice. Volume of injected tumors over time by treatment (PBS: N=6 Mice; i.t. VNP20009 injection on day 13 post implantation with or without enrofloxacin infusion starting one day later: N=5 Mice/Group). Repeated measure ANOVA was utilized to compare the tumor volumes between the different groups. Data are shown as mean±SEM. **p<0.01.
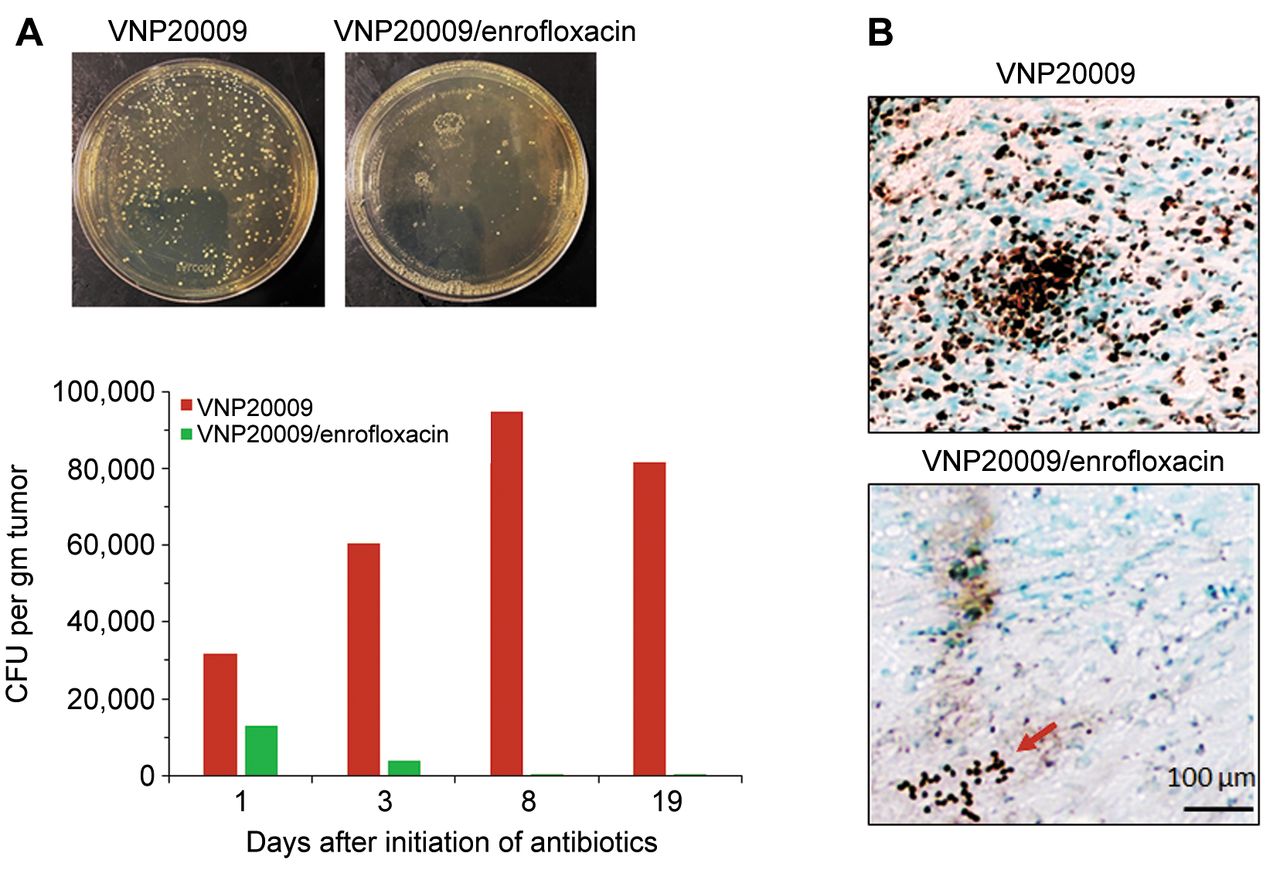
Early infusion of enrofloxacin abrogated S. typhimurium-induced tumor growth control and eliminated viable intratumoral VNP20009 cells. In the subcutaneous schwannoma bearing mice, infusion of enrofloxacin was initiated one day after i.t. injection with VNP20009. Tumor growth in the antibiotic-treated group was no different from that in the PBS-injected controls (p=0.98) (Figure 1). The S. typhimurium counts in tumors excised from the VNP20009 injected, enrofloxacin-treated group were 16%, 8%, and 0.3% compared to tumors from the VNP20009-injected, antibiotic-untreated mice on days 1, 3, and 7, respectively, after onset of enrofloxacin treatment (Figure 2A). No bacteria were detected in tumor tissues at the end of the experiment (day 18 post enrofloxacin treatment) but were present in tumors from non-antibiotic treated mice. The reduction in S. typhimurium counts in tumors that were injected with VNP20009 in the presence of enrofloxacin was further confirmed by immunohistochemical staining for bacterial outer membrane protein A (ompA) in tumor sections collected at the end of the study (day 32). While VNP20009-injected tumors removed from mice that did not receive antibiotics showed abundant bacterial staining, only scant bacterial rods were observed in i.t. VNP20009-injected tumors excised from mice subsequently treated with s.c. enrofloxacin (Figure 2B).

Early infusion of enrofloxacin eliminated viable VNP20009 cells from the implanted tumors. A) Colony forming units (CFUs) of bacteria present in the implanted schwannoma tumors following surgical removal on days 1, 3, 7, and 18 (N=2/group/time point) after initiation of enrofloxacin infusion, homogenization in PBS, and serial plating on LB plates. Images of LB agar plates represent the bacterial CFUs collected from tumors on day 18. B) Immunohistochemical staining of bacterial ompA in the tumor sections collected on day 18 shows abundant bacterial brown positive staining in the VNP20009 injected tumors, while only scant bacterial colonies were detected in the mice received both i.t. VNP20009 and enrofloxacin infusion. Red arrow indicates positive staining.
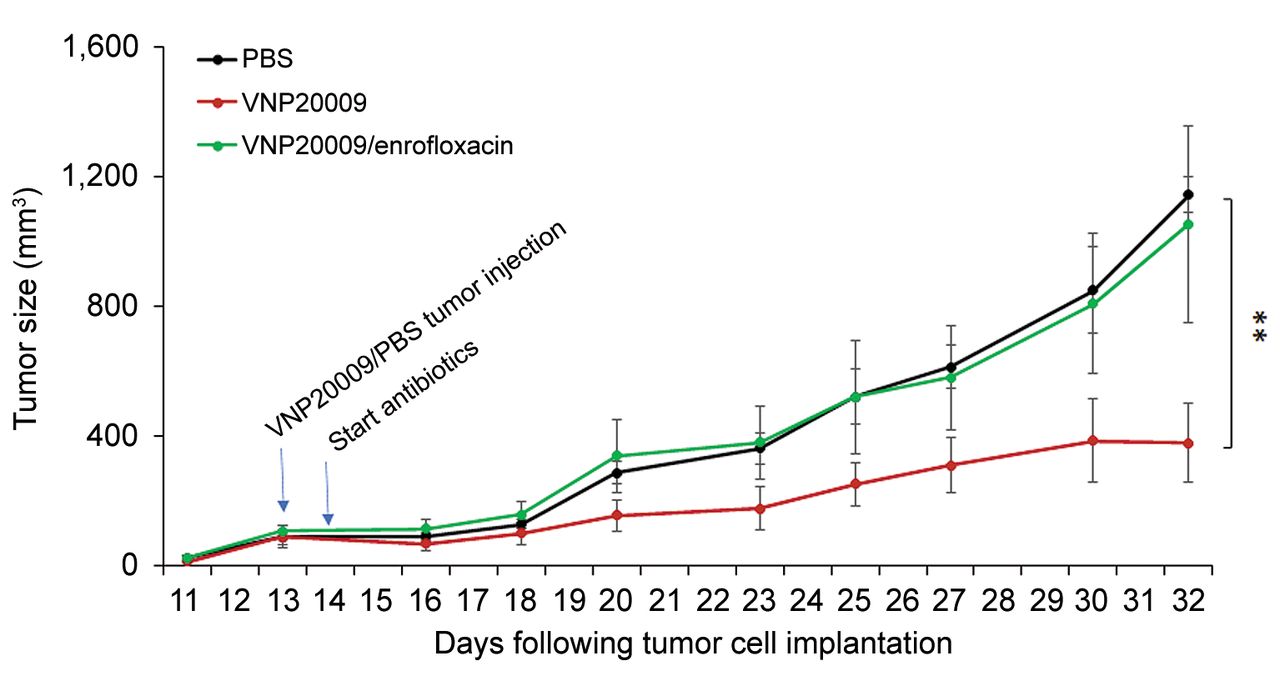
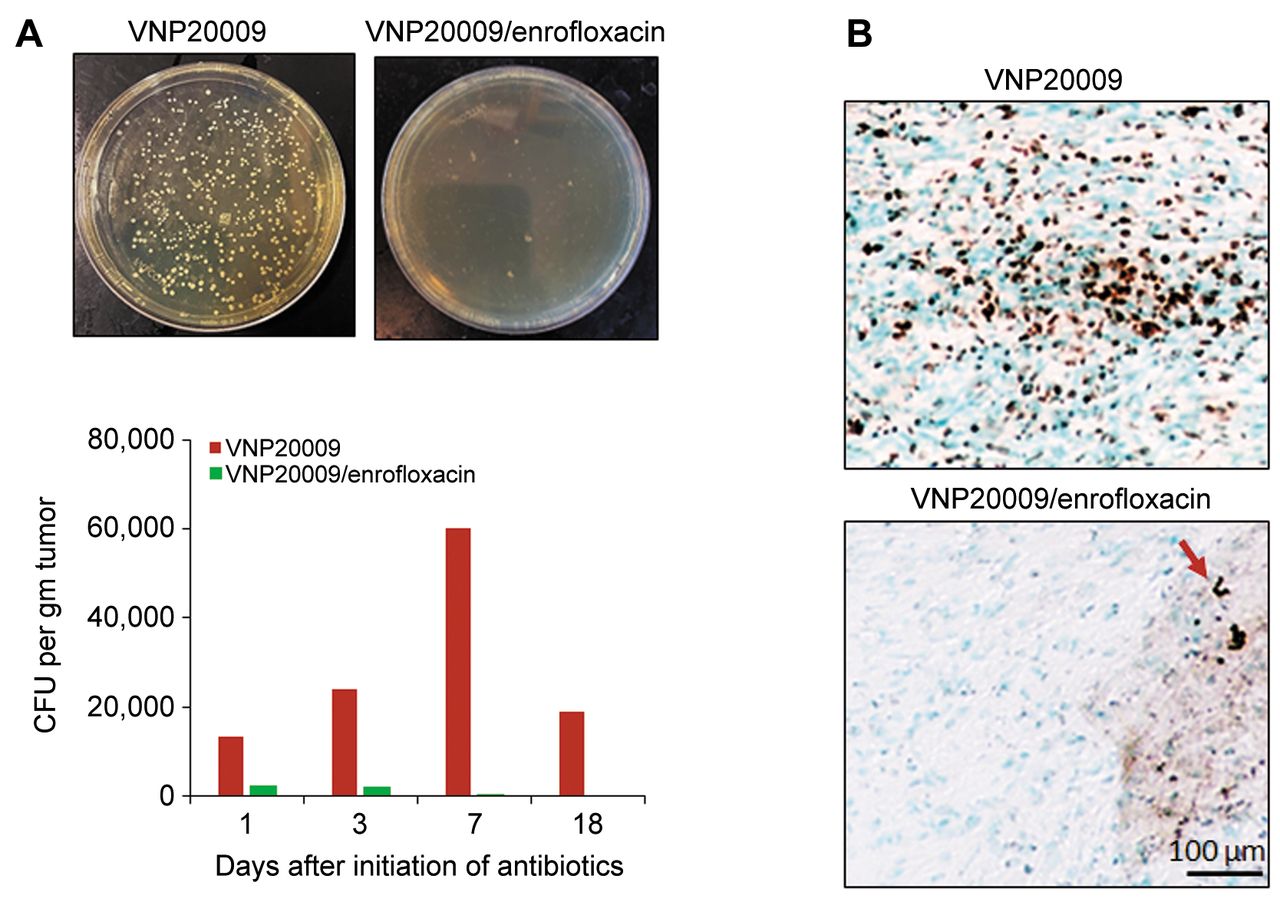
I.t. S. typhimurium suppressed schwannoma growth in mice treated with enrofloxacin starting 8 days following bacterial injection but less than that observed in VNP20009-injected non-antibiotic treated mice. In a second experiment, antibiotic infusion was initiated eight days post i.t. VNP20009 injection. Approximately five days after initiation of enrofloxacin infusion, tumor growth in the antibiotic-treated group started to increase relative to non-antibiotic-treated VNP20009-injected mice, with this difference becoming statistically significant at three weeks after initiation of enrofloxacin infusion (Figure 3). This relative increase in tumor growth correlated with a decrease in viable and stained S. typhimurium in tumors excised from i.t. VNP20009-injected, enrofloxacin-treated animals (Figure 4A and B).

Delay of infusion of enrofloxacin until 8 days after i.t. bacterial injection abrogated S. typhimurium-induced tumor growth retardation of schwannoma tumor development in syngeneic mice. Volume of injected tumors over time by treatment (PBS: n=9 mice; i.t. VNP20009 injection with or without enrofloxacin infusion starting eight days later: n=8 mice/group). Repeated measure ANOVA was utilized to compare the tumor volumes between the different groups. Data are shown as mean±SEM. **p<0.01.
Infusion of enrofloxacin eight days after S. typhimurium VNP20009 injection reduced viable bacteria in the implanted tumors. A) Colony forming units (CFUs) of bacteria present in the implanted schwannoma tumors following surgical removal on days 1, 3, 8 and 19 (N=2/group/data point) after initiation of enrofloxacin infusion, homogenization in PBS, and serial plating on Luria-Bertani (LB) plates. Images of LB agar plates represent the bacterial CFUs collected from tumors on day 19. B) Immunohistochemical staining of bacterial ompA in the tumor sections collected on day 19 shows plentiful bacterial brown positive staining in the VNP20009 injected tumors, while only scattered bacterial colonies were detected in the mice received both i.t. VNP20009 and enrofloxacin infusion. Red arrow indicates positive staining.
Discussion
We recently demonstrated that intratumoral injection of attenuated Salmonella typhimurium strain VNP20009 is a promising schwannoma therapeutic strategy (12). The current study indicated that the presence of viable VNP20009 is required for bacteria-mediated schwannoma growth control, as demonstrated by the abrogation of VNP20009 mediated tumor growth control by enroflaxacin infusion initiated at 1 day post i.t. VNP20009 injection. Initiating enrofloxacin 8 days following i.t. VNP20009 injection diminished bacteria-mediated schwannoma growth control but did not abolish the effect. This finding supports the hypothesis that ongoing colonization by VNP20009 is essential for efficacy and is a critical consideration for translation of attenuated S. typhimurium to clinical trials. Another important consideration for clinical trials of any BCT is the ability to stop the clinical trial early and control the effects of bacterial treatment in the event study termination is warranted because of safety concerns. For example, in previous clinical trials of VNP20009 in advanced or metastatic tumors, the administration of ciprofloxacin, ceftriaxone, cefixime, and co-trimoxazole has been prescribed for the control of treatment in the event of any dose-limiting toxicity (6, 7) (see also clintrials.gov, Study NCT00004216). VNP20009 has been shown to be susceptible to each group of antibacterial agents deemed as acceptable for testing of Salmonella by the National Committee for Clinical Laboratory Standards (7). The results of the current study indicate that in clinical trials of VNP20009 schwannoma treatment the S. typhimurium would be similarly responsive to antibiotic intervention.
Acknowledgements
The Authors are grateful for the funding provided by the Department of Anesthesia, Critical Care & Pain Medicine at Massachusetts General Hospital, MA, USA, and charitable donations by Neurofibromatosis Northeast and NF2 BioSolutions to the Rebecca Grasso Fund. The Authors thank Tim Ahern, a technical writer supported by Mulberry Biotherapeutics, for his editorial comments and help with the preparation of this manuscript.
Footnotes
Authors’ Contributions
Initial conceptualization, experimental design, conducting experiments, data analysis, manuscript preparation: Sherif G. Ahmed. Initial conceptualization, experimental design, data analysis, manuscript preparation: Gary J. Brenner.
Conflicts of Interest
G.J.B. has a financial interest in Mulberry Biotherapeutics, Inc. (Wellesley, MA, USA), a company developing novel biologic therapies for schwannoma and related neoplasms. G.J.B.’s interests were reviewed and are managed by Massachusetts General Hospital and Mass General Brigham in accordance with their conflict-of-interest policies.
- Received November 15, 2022.
- Revision received November 28, 2022.
- Accepted November 29, 2022.
- Copyright © 2023 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.