


Abstract
Background/Aim: p62 (also known as sequestosome 1) is involved in cancer progression, and high expression of p62 indicates poor clinical outcome in several cancer types. However, the association between p62 gene expression and cancer stem cells (CSCs) in breast cancer subtypes remains unclear. Materials and Methods: In the present study, genomic datasets of primary breast cancer (The Cancer Genome Atlas, n=593; and Molecular Taxonomy of Breast Cancer International Consortium, n=2,509) were downloaded. p62 Expression was then examined in normal and breast cancer tissues derived from the same patients. Kaplan-Meier and multivariate Cox regression analyses were employed to evaluate disease-specific survival. Next, the effect on cell viability and in vitro tumor-sphere formation of p62 knockdown using targeted small interfering RNA was assessed by using cells with high activity of aldehyde dehydrogenase 1 (ALDH1high). Results: Patients with normal-like, luminal A or luminal B breast cancer with p62high had poor prognosis. Furthermore, patients with p62high ALDH1A3high luminal B type also exhibited poor prognoses. Knockdown of p62 suppressed viability and tumor-sphere formation by ALDH1high cells of the luminal B-type cell lines BT-474 and MDA-MB-361. These results suggest that p62 is essential for cancerous progression of ALDH1-positive luminal B breast CSCs, and contributes to poor prognosis of luminal B breast cancer. Conclusion: p62 is potentially a prognostic marker and therapeutic target for ALDH1-positive luminal B breast CSCs.
Breast cancer has the highest prevalence among cancers of women worldwide, with 2.26 million new cases (24.5% of all cancer cases in women) and 685,000 cancer-associated mortalities (15.5% of all cancer-associated mortalities among women) annually (1). Breast cancer is classified using two parameters: Immunohistochemistry and gene-expression patterns [prediction analysis of microarray 50 (PAM50)] (2-8). Based on its PAM50, breast cancer is classified into at least six subtypes: Normal-like, luminal A, luminal B, human epidermal growth factor receptor type 2 (HER2)-enriched, claudin-low and basal-like (5, 7, 8). Among these, the luminal B type expresses estrogen receptor, and certain luminal B tumors express HER2 and highly express proliferation-related genes such as marker of proliferation Ki-67 (MKI67). In addition, the luminal B type has poorer prognosis (7, 9-15). Breast cancer treatment mainly entails surgery, radiotherapy and drug therapy, including chemotherapy, endocrine therapy and molecular targeted therapy. However, there are still numerous refractory cases, and further stratification and development of prognostic markers and therapeutic targets are required for improving quality of life of patients. Luminal B type can be treated with endocrine therapy and a HER2-targeted antibody, such as trastuzumab (12, 14). Given its poor prognosis, it is necessary to identify effective prognostic markers and molecular targets for the luminal B breast cancer subtype.
Tumor consists of both cancer stem cells (CSCs) and differentiated cancer cells. CSCs exhibit stem cell-like functions such as self-renewal, multipotency and tumorigenicity (16, 17). Since the majority of CSCs are resistant to conventional antitumor treatments, such as chemotherapy and radiotherapy, the development of targeted therapies against CSCs is needed to improve the clinical outcomes of patients (17, 18).
Aldehyde dehydrogenase 1 (ALDH1) is an enzyme that converts aldehydes into carboxylic acids. ALDH1A1 and ALDH1A3 are known as CSC markers in several cancer types (19-24). Expression of ALDH1A1 and ALDH1A3 is correlated with tumor grade, metastasis and prognosis of patients with breast cancer (25-27), and ALDH1A3 significantly contributes to ALDH1 activity in breast cancer (25, 28).
p62 (also known as sequestosome 1) is a multifunctional adapter protein that is involved in various physiological functions (29), including nuclear factor-κB signaling (30-33), antioxidant response (34) and autophagy (35-37). p62 is overexpressed (38-48) in several cancer types, and high p62 expression is associated with poor prognosis (41, 44, 46, 49-52) in several types of cancer. In breast cancer, p62 gene and protein are overexpressed (53-55) and high p62 protein expression is associated with poor prognosis of overall and disease-free survival (54, 56). In addition, patients with high p62 gene expression have shorter recurrence-free (55, 57) and metastasis-free (55) survival. However, the association between p62 gene expression and CSCs among breast cancer subtypes remains unclear.
The present study examined the association between p62 and ALDH1A3 among breast cancer subtypes. Furthermore, we also examined the role of p62 in ALDH1A3-positive CSCs of luminal B breast cancer.
Materials and Methods
Analysis of The Cancer Genome Atlas dataset (TCGA). TCGA breast cancer dataset (58) was downloaded from Oncomine (https://www.oncomine.org; Thermo Fisher Scientific, Inc., Waltham, MA, USA) (59) on January 5, 2021. This dataset contains mRNA expression data from 61 normal breast tissue and 532 primary breast tumor samples, and the clinicopathological data of these patients had been previously reported (60). The expression of p62 mRNA (Probe ID: A_23_P81399, A_23_P81401) is presented using the log2 median-centered ratio for both normal and cancer tissues. p62 mRNA were plotted using a paired comparison of normal versus cancer tissue from the same patients (n=60) using the Wilcoxon signed-rank test. Two-sided values of p<0.05 were considered to indicate a statistically significant difference. All statistical analyses were carried out using BellCurve for Excel ver. 3.10 (Social Survey Research Information Co., Ltd., SSRI, Tokyo, Japan).
Analysis of the Molecular Taxonomy of Breast Cancer International Consortium (METABRIC) dataset. The METABRIC dataset (n=2,509) (61, 62) was downloaded from cBioportal (https://www.cbioportal.org/) (63, 64) on October 30, 2020. The clinicopathological data from these patients had been previously summarized (60). The METABRIC dataset contains data on both gene alterations (n=2,173) and mRNA expression levels of primary breast cancer samples (n=1,904), and disease-specific survival (DSS) data with mRNA expression levels (n=1,423). The mRNA expression levels were compared among breast cancer subtypes using the Kruskal-Wallis test followed by Steel-Dwass’s multiple comparison test for post-hoc analysis. The optimal cutoff thresholds to divide patients into groups with high and low expression were defined using receiver operating characteristic curves associating p62, ALDH1A1 or ALDH1A3 gene expression with DSS. The optimal cutoff threshold was determined using the Youden’s index. Survival curves based on DSS were plotted using the Kaplan-Meier method, and curves were compared using the log-rank (Cochran-Mantel-Haenszel) test. A multivariate Cox regression analysis was used to evaluate the influence of gene expression and to estimate adjusted hazard ratios (HRs) using DSS statuses, with age at diagnosis as a confounding factor. Two-sided values of p<0.05 were considered to indicate a statistically significant difference. All statistical analyses were carried out using BellCurve for Excel ver. 3.10 (SSRI).
Cell culture and siRNA transfection. The human luminal B type breast cancer cell lines BT-474 and MDA-MB-361, and human normal (nontransformed) mammary epithelial cell line MCF10A were obtained from the American Type Culture Collection (Manassas, VA, USA) and were cultured at 37°C in the presence of 5% CO2. BT-474 cells were cultured in Dulbecco’s modified Eagle’s medium containing 10% fetal bovine serum (Capricorn Scientific GmbH, Ebsdorfergrund, Germany) and 0.01 mg/ml insulin (Nacalai Tesque, Inc., Kyoto, Japan). MDA-MB-361 cells were cultured in Dulbecco’s modified Eagle’s medium containing 10% fetal bovine serum. MCF10A cells were maintained in mammary epithelial cell growth medium (Lonza, Basel, Switzerland).
p62 knockdown (KD) in BT-474 and MDA-MB-361 cells was achieved using transfection of two siRNAs and two Dicer-Substrate siRNAs (DsiRNAs), which led to long-term suppression of gene expression (65). The sequences were as follows. p62 siRNA-1: 5’-GUGAACUCCAGUCCCUACATT-3’ and p62 siRNA-2: 5’-GUGACGAGGAAUUGACAAUTT-3’) (Sigma-Aldrich; Merck KGaA, St. Louis, MO, USA). p62 DsiRNA-1: sense strand: 5’-GUGAACUCCAGUCCCUACAGAUGCC-3’ and antisense strand: 5’-GGCAUCUGUAGGGACUGGAGUUCACCU-3’; and p62 DsiRNA-2: sense strand: 5’-GUGACGAGGAAUUGACAAUGGCCAT-3’ and antisense strand: 5’-AUGGCCAUUGUCAAUUCCUCGUCACUG-3’ (Integrated DNA Technologies, Inc., IDT, Coralville, IA, USA). MISSION siRNA Universal Negative Control (Sigma-Aldrich; Merck KGaA) and Negative Control DsiRNA (sense strand: 5’-CGUUAAUCGCGUAUAAUACGCGUAT-3’ and antisense strand: 5’-AUACGCGUAUUAUACGCGAUUAACGAC-3’) were used as controls. Transfection was performed using Lipofectamine RNAiMAX (Thermo Fisher Scientific, Inc.). Cells were transfected with 10 nM siRNA and incubated for 24 h, followed by transfection with 10 nM DsiRNA and subsequent incubation for an additional 24 h before assays were performed.
Western blotting. Western blotting of cells was performed as previously described (60, 66-70). The following antibodies were used: Rabbit polyclonal anti-p62 (PM045, 1:10000; Medical & Biological Laboratories, Tokyo, Japan); mouse monoclonal anti-p62 (610833, 1:10,000; BD Biosciences, San Jose, CA, USA); rabbit monoclonal anti-ALDH1A1 (ab52492, 1:3,000; Abcam, Cambridge, UK); rabbit polyclonal anti-ALDH1A3 (PA5-29188, 1:5,000; Thermo Fisher Scientific, Inc.); mouse monoclonal anti-β-actin (60008-I-Ig, 1:10,000; ProteinTech Group, Inc., Rosemont, IL, USA) as the primary antibodies. Goat anti-mouse and anti-rabbit IgG (7076S and 7074S, 1:5,000; Cell Signaling Technology, Inc., Danvers, MA, USA) were used as secondary antibodies.
Isolation of ALDH1high cells. ALDH1high cells were isolated as previously described (60, 66-68, 70, 71) from BT-474 and MDA-MB-361 cells with p62 KD using ALDEFLUOR™ assay kit (STEMCELL Technologies, Inc., Vancouver, Canada).
WST-8 assay. WST-8 assay was performed as previously described (60, 66, 67, 69, 71). Briefly, after p62 KD, unsorted cells or sorted ALDH1high cells were seeded into 96-well plates (1×105 cells/well) (Thermo Fisher Scientific, Inc.) and incubated for 5 days (BT-474 and unsorted MDA-MB-361 cells) or 9 days (ALDH1high cells derived from MDA-MB-361 cells). Cell viability was then assessed using Cell Counting Reagent SF (Nacalai Tesque, Inc.). The results of unsorted cells were analyzed by Dunnett’s multiple comparison test (unpaired). The results of sorted ALDH1high cells were analyzed by Student’s t-test. Two-sided values of p<0.05 were considered to indicate a statistically significant difference. Data are presented as the mean±standard error of the mean of three independent experiments. All statistical analyses were carried out using BellCurve for Excel ver. 3.10 (SSRI).
Tumor-sphere culture. Tumor-sphere formation assay was conducted as previously described (60, 66-71). ALDH1high cells were isolated via ALDEFLUOR assay upon p62 KD. The isolated ALDH1high cells were plated in ultralow-attachment 96-well plates (5×104 cells/well) (Greiner Bio-One GmbH, Frickenhausen, Germany) and cultured for 7 days (BT-474) or 15 days (MDA-MB-361) and were captured using a DMIL LED microscope (Leica, Wetzlar, Germany). The number and size of tumor spheres over 1,256 μm2 (BT-474) and 314 μm2 (MDA-MB-361) were measured using ImageJ 1.51j8; Java 1.8.0_112 (64-bit) (National Institutes of Health, Bethesda, MA, USA). Statistical significance was determined with Student’s t-test. Two-sided values of p<0.05 were considered to indicate a statistically significant difference. Data are presented as the mean±standard error of the mean of three independent experiments. All statistical analyses were carried out using BellCurve for Excel ver. 3.10 (SSRI).
Results
Kaplan–Meier analyses indicate that clinical outcomes are poor for patients with p62high normal-like, luminal A or luminal B breast cancer. p62 is highly expressed in breast cancer compared with normal mammary epithelium (53, 55). Consistent with this, the present study confirmed that p62 expression was significantly higher in breast cancer in paired comparison of p62 mRNA expression between normal and tumor tissues derived from the same patients in TCGA breast cancer dataset (Figure 1A). Next, p62 expression was examined in different breast cancer subtypes using the METABRIC dataset. Among the subtypes classified based on PAM50, p62 was highly expressed in the normal-like, luminal A, luminal B and HER2-enriched subtypes than in the claudin-low and basal-like types (Figure 1B).

p62 is highly expressed in breast cancer, and patients with p62high normal-like, luminal A and luminal B breast cancer subtypes exhibit poor clinical outcomes. A: Paired comparison of p62 mRNA expression in normal and tumor tissues derived from the same patients (n=60) in the The Cancer Genome Atlas with Probe IDs A_23_P81399 (left) and A_23_P81401 (right). Median values are shown at the bottom of graphs. Significantly different at **p<0.01 by Wilcoxon signed-rank test. B: p62 mRNA expression by breast cancer subtype. Significantly different at: *p<0.05, **p<0.01 and ***p<0.001 by Kruskal–Wallis test followed by Steel–Dwass’s multiple comparison test for post-hoc analysis. C: Kaplan– Meier analyses comparing disease-specific survival (DSS) according to p62 mRNA expression in breast cancer overall and by breast cancer subtype. p-Values were calculated with the log-rank (Cochran–Mantel–Haenszel) test. HER2: Human epidermal growth factor receptor type 2.
To assess the role of p62 expression in PAM50 subtypes, the present study analyzed a METABRIC dataset that included the gene-expression data from patients with breast cancer. The association between p62 expression and prognosis among the breast cancer subtypes was examined by using the Kaplan–Meier method to compare DSS between patients with p62high and p62low disease. Firstly, patients with p62high status had a poor prognosis in breast cancer overall (p<0.001) (Figure 1C). Among the breast cancer subtypes, patients with p62high disease had significantly poorer clinical outcomes than those with p62low normal-like, luminal A or luminal B disease (Figure 1C). Importantly, there was no significant difference between the survival rates of patients with p62high and p62low HER2-enriched, claudin-low or basal-like type (Figure 1C). These results suggest that p62 is involved in cancer progression and contributes to poor clinical outcomes in patients with normal-like, luminal A and luminal B breast cancer.
Kaplan–Meier analyses indicate that patients with p62high ALDH1A3high luminal B breast cancer have a poorer clinical outcome. ALDH1A1 and ALDH1A3 are known to be CSC markers in several cancer types (19-24). Therefore, the present study next assessed the prognosis of patients with p62high ALDH1A1high and of those with p62high ALDH1A3high status. Kaplan–Meier analyses did not show significant differences between the DSS rates of patients with p62high ALDH1A1high and patients with other statuses in breast cancer overall nor in any subtype (Figure 2A). Kaplan–Meier analyses showed that patients with p62high ALDH1A3high had poorer clinical outcomes than those with other statuses in breast cancer overall and in those with luminal B breast subtype (Figure 2B). Importantly, there was no significant difference between the survival rates of patients with p62high ALDH1A3high disease and other patients with normal-like, luminal A, HER2-enriched, claudin-low or basal-like type (Figure 2B). In comparison of DSS among patients according to combined expression of p62 with ALDH1A1, no significant differences were noted in breast cancer overall, nor in any subtype (Figure 3A), whereas DSS of patients with p62high ALDH1A3high status was significantly poorer only in those with the luminal B breast cancer subtype (Figure 3B). These results suggest that p62 is involved in cancer progression and contributes to poor clinical outcomes of patients with ALDH1A3high luminal B breast cancer.
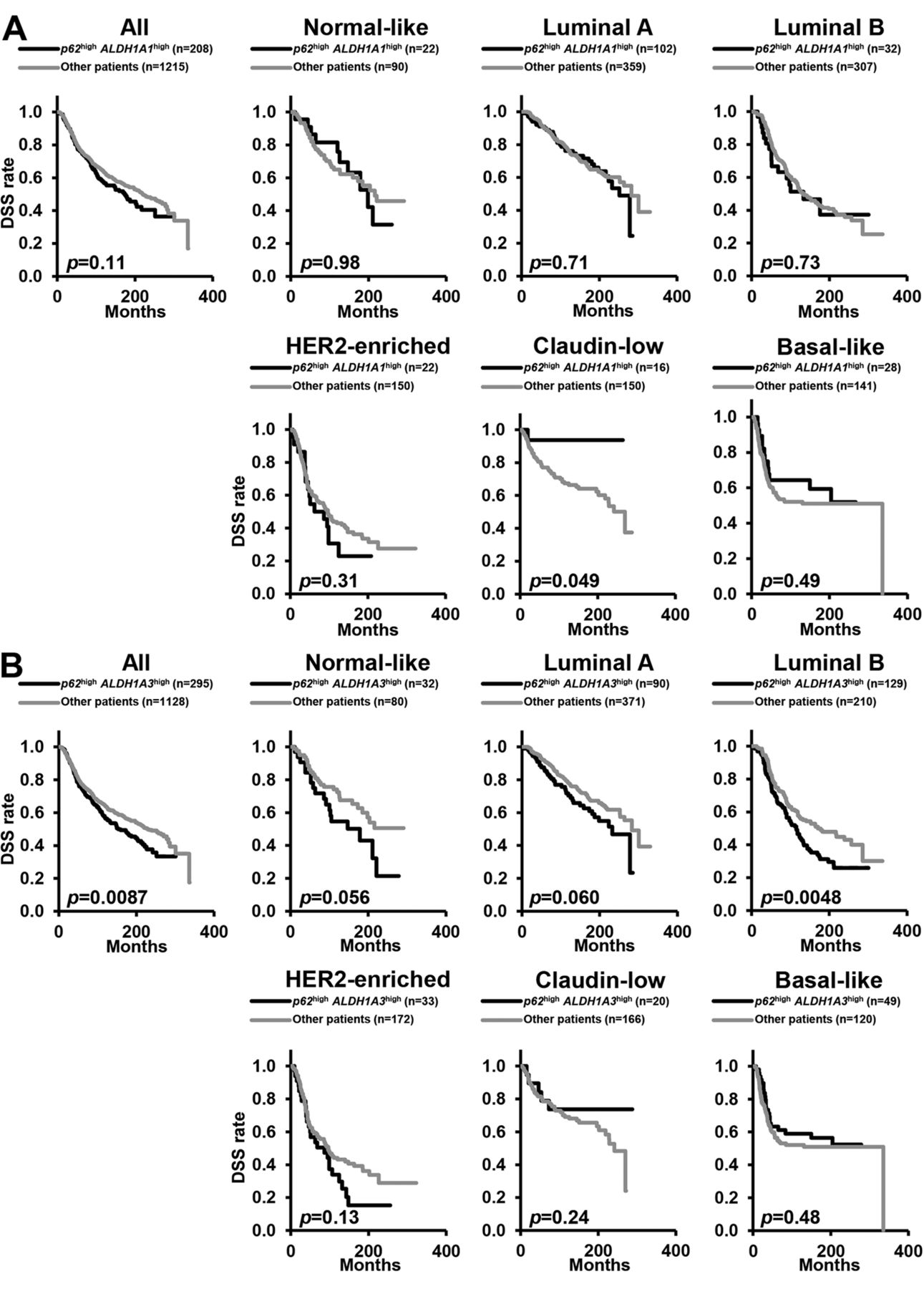
Patients with luminal B breast cancer with high expression of p62 (p62high) and aldehyde dehydrogenase 1A3 (ALDH1A3high) exhibit poor clinical outcomes. A: Kaplan–Meier analyses comparing disease-specific survival (DSS) between patients with p62high ALDH1A1high disease and others (p62high ALDH1A1low, p62low ALDH1A1high and p62low ALDH1A1low) in breast cancer overall and by breast cancer subtype. B: Kaplan– Meier analyses comparing DSS between p62high ALDH1A3high patients and others (p62high ALDH1A3low, p62low ALDH1A3high and p62low ALDH1A3low) in breast cancer overall and by breast cancer subtype. p-Values were calculated with the log-rank. (Cochran-Mantel-Haenszel) test. HER2: Human epidermal growth factor receptor type 2.

Patients with luminal B breast cancer with high p62 (p62high) and high aldehyde dehydrogenase 1A3 (ALDH1A3high) expression exhibit poor clinical outcomes. Kaplan-Meier analyses comparing DSS according to combined expression of p62 with ALDH1A1 (A) and with ALDH1A3 (B) in breast cancer overall and by breast cancer subtype. p-Values were calculated with the log-rank (Cochran-Mantel-Haenszel) test. HER2: Human epidermal growth factor receptor type 2.
Multivariate Cox regression analysis indicates that patients with p62high ALDH1A3high luminal B breast cancer have a poorer clinical outcome. To confirm the results obtained by Kaplan-Meier analyses, a multivariate Cox regression analysis of DSS was performed with age at diagnosis as a confounding factor using the same dataset (Table I). Patients with p62high status had significantly poorer clinical outcomes considering breast cancer overall, and normal-like or luminal A subtype than those with p62low disease (Table I). Patients with p62high luminal B, HER2-enriched, claudin-low or basal-like subtypes did not have poorer clinical outcomes than those with p62low status (Table I). Multivariable Cox regression analysis also showed that p62high ALDH1A1high status was not predictive of poorer clinical outcomes than other p62 ALDH1A1 statuses in breast cancer overall nor in any subtype (Table I). However, p62high ALDH1A3high disease was predictive of significantly poorer clinical outcomes compared with other p62 ALDH1A3 statuses in breast cancer overall and in patients with luminal B breast subtype (Table I) but not in those with other breast cancer subtypes (Table I). These results suggest that p62 is involved in cancer progression and contributes to poor prognosis of ALDH1A3high, i.e.-CSC-enriched, luminal B breast cancer.
Multivariable Cox regression analysis of disease-specific survival according to expression of p62, aldehyde dehydrogenase 1 (ALDH1A1) and ALDH1A3 by breast cancer subtype.
p62 siRNA KD suppresses cell viability and in vitro tumorsphere formation by ALDH1high cells. Based on the above results, the present study next examined the possibility that p62 may serve as a therapeutic target of ALDH1A3-positive CSCs in luminal B breast cancer. The luminal B type breast cancer cell lines BT-474 and MDA-MB-361 were used, which overexpress p62 protein compared with the normal human (non-transformed) mammary epithelial cell line MCF10A (Figure 4A). p62 siRNA significantly suppressed p62 protein expression in BT-474 and MDA-MB-361 cells (Figure 4B). WST-8 cell viability assays with BT-474 and MDA-MB-361 cells showed that p62 KD suppressed cell viability (Figure 4C). These findings suggest that p62 contributes to cell survival/proliferation in luminal B breast cancer.
ALDH1A1 protein was slightly expressed in BT-474 and MDA-MB-361 cells. ALDH1A3 protein was overexpressed in BT-474 compared with MCF10A and MDA-MB-361 cells (Figure 4A). Next, to assess the role of p62 in ALDH1-positive luminal B CSCs, ALDEFLUOR assays were used to examine the effects of p62 depletion on cell viability and in vitro tumor-sphere formation in ALDH1high BT-474 and MDA-MB-361 cells. p62 KD led to suppression of cell viability by ALDH1high BT-474 and MDA-MB-361 cells (Figure 4D). This result suggests that p62 is essential for survival/proliferation. In addition, p62 siRNA suppressed the in vitro area of tumor spheres formed by ALDH1high BT-474 and MDA-MB-361 cells but it did not suppress the number of in vitro tumor spheres (Figure 4E, respectively). These results suggest that p62 is also essential for tumor formation by ALDH1-positive breast CSCs in luminal B breast cancer. Furthermore, p62 may be a therapeutic target as well as a prognostic marker in ALDH1-positive luminal B breast cancer.
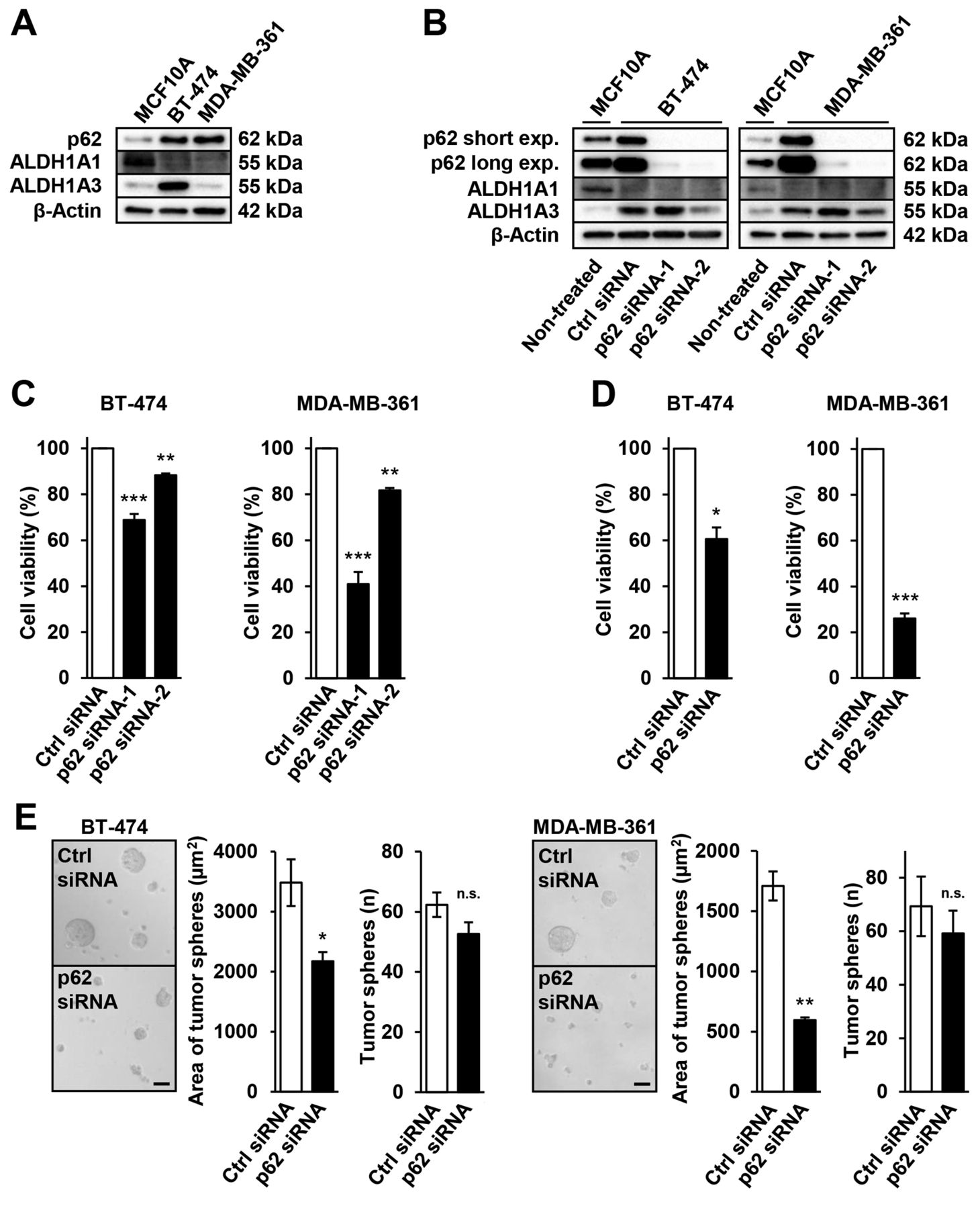
Knockdown of p62 with siRNA leads to the suppression of cell viability and in vitro tumor-sphere formation by luminal B breast cancer cells with high aldehyde dehydrogenase 1 (ALDH1) expression. A: Western blotting of p62, ALDH1A1 and ALDH1A3 expression in native MCF10A, BT-474 and MDA-MB-361 cells. B: Western blotting of p62, ALDH1A1 and ALDH1A3 in BT-474 and MDA-MB-361 cells with and without knockdown of p62 using targeted siRNA-1/2 and DsiRNA-1/2. Non-treated MCF10A cells acted as a positive control. C: WST-8 assays assessing cell viability of BT-474 and MDA-MB-361 cells after p62 knockdown using targeted siRNA-1/2 and DsiRNA-1/2. D: WST-8 assays assessing cell viability of ALDH1high BT-474 and MDA-MB-361 cells after p62 knockdown using targeted siRNA-1 and DsiRNA-1. E: In vitro tumor sphere-formation culture was performed to assess tumor-sphere formation by ALDH1high BT-474 and MDA-MB-361 cells following p62 knockdown via targeted siRNA-1 and DsiRNA-1. Representative images are shown; scale bar, 50 μm. Data represent the mean±standard error of the mean of three independent experiments. Significantly different at: *p<0.05, **p<0.01 and ***p<0.001 by Student’s t-test or Dunnetts multiple comparison (unpaired). n.s.: Not significantly different.
Discussion
The present study demonstrated that p62 was essential for survival/proliferation and tumor formation by ALDH1high breast CSCs in luminal B breast cancer, and was involved in cancer progression and contributed to poor prognosis of ALDH1A3high luminal B breast cancer.
p62 gene amplification and enhancement of gene expression were detected in clear-cell renal cell carcinoma (72). Therefore, p62 gene alterations were assessed using the METABRIC and TCGA datasets in this study. p62 gene amplification was detected in 1.7% and 1.0% of breast cancer cases, respectively (METABRIC: 37/2,173; and TCGA: 10/996), p62 mutation was detected in 0.0 and 0.50% (METABRIC: 0/2,173; TCGA: 5/996), and deletion was detected in 0.092 and 0.20% (METABRIC: 2/2,173; TCGA: 2/996), respectively. Thus, the high level of p62 mRNA expression observed in breast cancer compared with that of normal tissues may reflect the transcriptional activation of p62. It has been reported that p62 is a target gene in cancer, which is located downstream of nuclear factor-E2-related factor 2, nuclear factor-κB and activator protein-1 (34, 73, 74). A detailed mechanism of p62 gene expression in ALDH1-positive luminal B CSCs will need to be investigated.
High p62 expression is associated with poor prognosis in several cancer types, including breast cancer (38-48, 53-55). However, few reports have mentioned an association between p62 gene expression and prognosis, and none have mentioned breast cancer subtypes. The present results revealed that patients with p62high normal-like, luminal A and luminal B breast cancer exhibited poor prognosis (Figure 1C), and patients with p62high ALDH1A3high luminal B breast subtype had poor clinical outcomes (Figure 2B and Figure 3B; Table I). These results suggest that p62 is involved in cancer progression and contributes to poor prognosis of ALDH1A3high patients, that is, patients who have luminal B ALDH1A3-positive breast CSCs. Among the breast cancer subtypes, p62 mostly appears to contribute to the poor prognosis of luminal B type. In normal-like and luminal A breast cancer, p62 might be associated with CSC markers other than ALDH1A1 and ALDH1A3. ALDH1A1 contributes to adhesion, migration, extravasation, initial colonization steps (28) and poor prognosis (26); however, the present results suggest that p62 contributes to survival/proliferation of and tumor formation by ALDH1A3-positive (not ALDH1A1-positive) CSCs in luminal B breast cancer (Figure 2A and Figure 3A; Table I). Thus, p62-targeted therapies may be suitable for ALDH1A3-positive CSC-enriched luminal B breast cancer.
The characteristics of luminal B type compared with the luminal A type are as follows: Luminal B is often HER2-positive and exhibits high expression of proliferation-related genes such as MKI67, as well as higher histological grade and worse prognosis (7, 9-15). As far as we are aware, there have been no reports to date on the association between p62 and luminal B breast cancer by PAM50 classification; however, it has been reported that the expression of p62 protein is correlated with that of HER2 in breast cancer (75, 76), and that p62 facilitates HER2-induced mammary tumorigenesis through multiple signaling pathways (77). On the other hand, one of the characteristics of luminal B type compared with HER2-enriched type is high estrogen receptor expression (4, 5, 8). Thus, the contribution of p62 to luminal B type ALDH1A3-positive CSCs may be associated with HER2 and estrogen receptor.
As shown in Figure 4C, p62 contributes to cell survival/proliferation in luminal B breast cancer cells. Xu et al. reported that colony numbers were suppressed by p62 depletion via short hairpin RNA, and that tumor xenografts derived from stably transfected, p62-depleted MDA-MB-231 cells exhibited significantly reduced growth rates and numbers of Ki67-positive cells (54). Nozaki et al. reported that MDA-MB-453 and MFM-223 cells transfected with p62 siRNA exhibited significantly reduced cell proliferative activity according to the results of 5-bromo-2-deoxyuridine incorporation assay (78). Notably, the results of our study revealed that p62 KD led to stronger suppression of the viability of sorted ALDH1high cells than of unsorted cells (Figure 4C and D). This suggests that the contribution of p62 to cell survival/proliferation is greater in ALDH1-positive CSCs than in other differentiated cells.
In vitro,p62 KD suppressed the size of tumor spheres, but did not significantly reduce the number of tumor spheres formed by ALDH1high cells (Figure 4E). These results indicate that p62 is involved in cell proliferation rather than in cell survival in ALDH1-positive luminal B breast CSCs.
It has been reported that p62 contributes to the characteristics exhibited by CSCs, including survival/proliferation, tumor formation (54, 78), infiltration, metastasis (55), and chemotherapy and radiotherapy resistance (47, 52, 79-81) in various cancer types, including breast cancer. p62 participates in maintaining breast cancer stem-like properties by stabilizing MYC proto-oncogene bHLH transcription factor mRNA (54), and nuclear fctor-E2-related factor 2 activation (82). It is necessary to investigate the mechanism by which p62 is involved in these properties exhibited by ALDH1-positive luminal B breast CSCs.
p62 protein interacts with atypical protein kinase C (PKCλ), and this interaction is involved in several biological functions (29, 83, 84). PKCλ is also essential for the survival and in vitro tumor-sphere formation of ALDH1-positive breast CSCs, and high expression of PKCλ and ALDH1A3 is associated with poor prognosis at late tumor-stage of breast cancer (68). Therefore, the association between p62 and PKCλ in ALDH1-positive luminal B breast CSCs should be further investigated.
The present study revealed that p62 contributes to survival/proliferation and tumor formation in ALDH1-positive luminal B breast cells, which leads to cancer progression and contributes to poor prognosis of ALDH1A3-positive, i.e. CSC-enriched, luminal B breast cancer. Overall, it can be concluded that p62 is potentially a prognostic marker and therapeutic target for ALDH1-positive luminal B breast CSCs.
Acknowledgements
This work was supported by Grant-in-Aid for Scientific Research (C) of JSPS (20K07207) (K.A.), JST Moonshot R&D (JPMJPS2022) (S.O.), Tokyo University of Science Grant for President’s Research Promotion (K.A.), Grant-in-Aid for JSPS Research Fellows (21J13718) (H.M.), Grant-in-Aid for Research Activity Start-up (21K20732) (S.T.), JST SPRING (JPMJSP2151) (A.O.), and Nagai Memorial Research Scholarship from the Pharmaceutical Society of Japan (A.O. and H.M.).
Footnotes
Authors’ Contributions
A.O., C.O., A.M., T.S. and Ke.S. performed bioinformatics; A.O., S.T., Ya.H. and A.M. performed the experiments; A.O., M.K., C.M., A.M., N.M., Yu.N. and Y.X. performed analyzed the data; Y.M., Yo.N., T.H., Yo.H., Ka.S. and S.O. supplied experimental materials and resources; A.O., H.M. and K.A. conceived the study; A.O. and K.A. drafted the article; all Authors contributed to discussion and review of the final article; and approved it.
Conflicts of Interest
The Authors declare that they have no competing interests in relation to this study.
- Received May 9, 2022.
- Revision received June 6, 2022.
- Accepted June 8, 2022.
- Copyright © 2022 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).





{kind=link}
{kind=link}
{kind=link}
{kind=link}