


Abstract
Background/Aim: miRNA functional analysis involves transfection with miRNA-based oligos to identify gain-of or loss-of function cellular phenotypes. Apoptosis is a common phenotypic endpoint for miRNA functional analysis. We report that four common cell dissociation enzymes, TrypLE, Accutase, Trypsin, and Accumax, can differentially impact cell viability and apoptosis in Annexin V flow cytometric analysis after miRNA-based transient transfection. Materials and Methods: We transiently transfected a nonsense oligo into an epithelial cancer cell line (UM-SCC-12) for 24 h. Cells were harvested with either TrypLE, Accutase, Accumax, or Trypsin after 5 min. The Annexin V/7-AAD assay via flow cytometry was employed. Studies were performed in triplicate. Significant effects were detected by ANOVA, followed by Tukey’s Multiple Comparison tests. Results: Trypsin produced the lowest cell viability and lowest percentage of apoptotic cells, specifically when compared to TrypLE and Accutase, respectively (p<0.01). Importantly, transfected trypsinized cells had a significant difference in cell viability and necrosis (p<0.05) when compared with non-transfected trypsinized cells, highlighting the influence of miRNA-based transfection on Annexin V flow cytometric outcomes. Interassay variability was lowest with TrypLE (1.13 %). As such, TrypLE provided the greatest reproducibility and reliability in our cell line. Conclusion: Our study highlights the variable effects of cell dissociation enzymes on transfected cells. Overall, the variability may lead to errors in detection of apoptotic cells using the Annexin V assay after miRNA-based transfection. Before assay use, we recommend pretesting cell dissociation enzymes on transfected cells to ensure reliable and reproducible results.
MicroRNAs (miRNAs) are small non-protein-coding RNAs that regulate expression of genes implicated in homeostatic processes such as apoptosis (1). Aberrant miRNA function may suppress apoptosis, which may, in turn, promote the growth of cancerous cells (1). In vitro transient transfection using either mimic or inhibitor miRNA oligos is commonly employed in combination with apoptosis assays to determine the manner in which deregulated miRNAs can influence tumor cell behavior. In apoptosis detection, flow cytometry has become one of the most widely used and superior methods to distinguish between viable, early apoptotic, late apoptotic or necrotic cells when combined with fluorescent staining of the Annexin V protein and an intercalating dye agent such as 7-Ammino-actinomycin D (7-AAD) (2-4). Annexin V protein binds to extracellular phosphatidylserine (PS) during early apoptosis, whereas 7-AAD stains DNA fragments exposed because the plasma membrane is compromised during later stages of apoptosis (2, 3, 5-8).
Successful execution and reliable interpretation of the Annexin V/7-AAD flow cytometry assay necessitates both coordinated timing of reagent input and the generation of live, healthy cells into a single cell suspension. However, cell membrane structure can be damaged depending on the method of cell dissociation used during preparation into single cell suspension, thereby reducing cell viability and generating false positive apoptotic or necrotic cells (9). Moreover, several studies have reported that transfection reagents alone can have cytotoxic and apoptotic effects on cells (10-12). These findings necessitate careful preparation of transfected cells into single cell suspension for subsequent flow cytometric apoptosis analysis.
Cell dissociation during the preparation of single cell suspensions may be achieved via enzymatic or non-enzymatic methods. Enzymatic cell dissociation can be achieved with different proteolytic and collagenolytic enzymes. Non-enzymatic dissociation can be achieved via non-enzymatic reagents or through mechanical means such as cell scraping. Generally, enzymatic dissociation is recommended in flow cytometric apoptosis assays because it more efficiently detaches cells and yields higher cell viability compared to non-enzymatic dissociation methods (13-17). However, some dissociation enzymes may also compromise cell viability and generate false positives in apoptosis/necrosis detection. Therefore, it is imperative to optimize gentler cell harvesting methods of transfected cells for subsequent apoptotic/cell viability flow cytometric analysis since transient transfection reagents may leave cells more susceptible to enzymatic damage.
To date, no studies have evaluated the impact of different enzymatic dissociation methods on cell viability in Annexin V/7-AAD flow cytometric analysis specifically after transient transfection. In this study, we used a nonsense miRNA oligo to transfect a laryngeal cancer cell line, and subsequently generated a single cell suspension using four different dissociation enzymes – Trypsin, TrypLE, Accutase, and Accumax. Here, we describe our results and provide further recommendation to improve the assay.
Materials and Methods
Transfection. Human laryngeal squamous cell carcinoma cell line, UM-SCC-12, was purchased from the University of Michigan Head and Neck cell line repository. Cells were cultured in a T 75 cm2 flask containing Dulbecco Modification of Eagle’s Medium 1X (DMEM, 10-013-CV, Corning, Manassas, VA, USA) supplemented with 10% heat inactivated fetal bovine serum (FBS, 35-011-CV, Corning, Woodland, CA, USA) and 2% Penicillin/Streptomycin (PENSTREP, 15-140-122, Gibco, Grand Island, NY, USA), within a humidified atmosphere containing 5% CO2 at 37°C. Approximately 1.25×105 cells per well were transiently transfected with miRCURY LNA miRNA Inhibitor Control (5) oligonucleotide containing TAACACGTCTATACGCCCA (339126 YI00199006-ADA, Qiagen, Germantown, MD, USA) in a 12-well plate using Lipofectamine RNAiMAX (13778075, Thermo Fisher, Carlsbad, CA, USA) and Opti-MEM I Reduced Serum Medium (31985062, Thermo Fisher, Grand Island, NY, USA) per manufacturer’s instructions. Control wells contained DMEM only.
Enzyme treatment. At 24 h post-transfection, cells were washed with 1ml of Dulbecco’s Phosphate Buffered Saline (1× dPBS, 21-031-CV, Corning, Manassas, VA, USA) per well. A volume of 500 ul of each cell dissociation enzyme was added to each well in triplicate. Cells were incubated at 37°C for 5 min (Figure 1) with either 1× TrypLE (12604-013, Gibco), Accutase (AT-104, Innovative Cell Technologies, San Diego, CA, USA), 0.25% Trypsin (25-053-Cl, Corning), and Accumax (AM105, Innovative Cell Technologies). Control cells were incubated with Trypsin at the same conditions. Complete cell detachment was confirmed via microscopy.

Schematic representation of conducted experiment in cells transfected with nonsense oligo for 24 h.
Preparation for Flow cytometry. Cells were harvested in the reverse order that the enzymes were administered such that the cells treated with Accumax, Trypsin, Accutase and TrypLE were respectively collected. Harvested cells were transferred to separate FACS tubes containing an equal volume of cold 1× dPBS. To wash the cells, each FACS tube was then centrifuged at 300 rcf for 5 min at 4°C and the supernatant was subsequently aspirated. Cells were washed two more times under the same conditions. After aspiration, a total of 150 μl volume was left in each tube. Figure 1 shows the plate layout and the schematic of the protocol.
Flow cytometric apoptosis analysis. The Annexin V/7-AAD kit was employed as per manufacturer’s instructions with a few modifications. Each FACs tube contained approximately 3.75×105 harvested cells. A volume of 100 μl of 1× Annexin binding buffer was added to each tube, resulting in a final total volume of approximately 250 μl. Next, 5 μl of Pacific Blue dye (A351316, Invitrogen, Waltham, MA, USA) and 5 μl of the 7-AAD viability staining dye (00-6993-50, 650 nm, Invitrogen) were added to each tube. Each tube was then incubated in the dark at room temperature for 30 min. Tubes were subsequently processed in the Cytometry Core at the University of Florida using the BD Fortressa (Gainesville, FL, USA) with BD FACSDiva Software (Version 8.0.2). Compensation was first performed using unstained Annexin V apoptosis and 7-AAD necrosis controls, yielding a spectral overlap of 0 between the Pacific Blue and 7-AAD dyes. Finally, fluorescence corresponding to cell viability, apoptosis and necrosis of the harvested cells was subsequently analyzed. The study was performed in triplicate.
Statistical analyses. All data generated from the three experimental runs were analyzed using GraphPad Prism (Version 9.2.0, San Diego, CA, USA) software with the alpha level set at 0.05. Two separate one-way ANOVAs were conducted to assess the effect of enzyme condition (TrypLE, Accutase, Accumax, Trypsin, and the DMEM control) on 1) percentage of viable cells and 2) percentage of apoptotic cells (early + late apoptotic cells labeled with Annexin V) detected in Annexin V/7-AAD flow cytometric apoptosis analysis. Significant effects detected in ANOVA were followed up with Tukey’s Multiple Comparison Tests. Results are presented as Mean±SD. The coefficient of variation (CV) of the percentage of viable cells was also calculated to assess the magnitude of variability of each enzyme condition across the triplicate flow cytometry runs.
Results
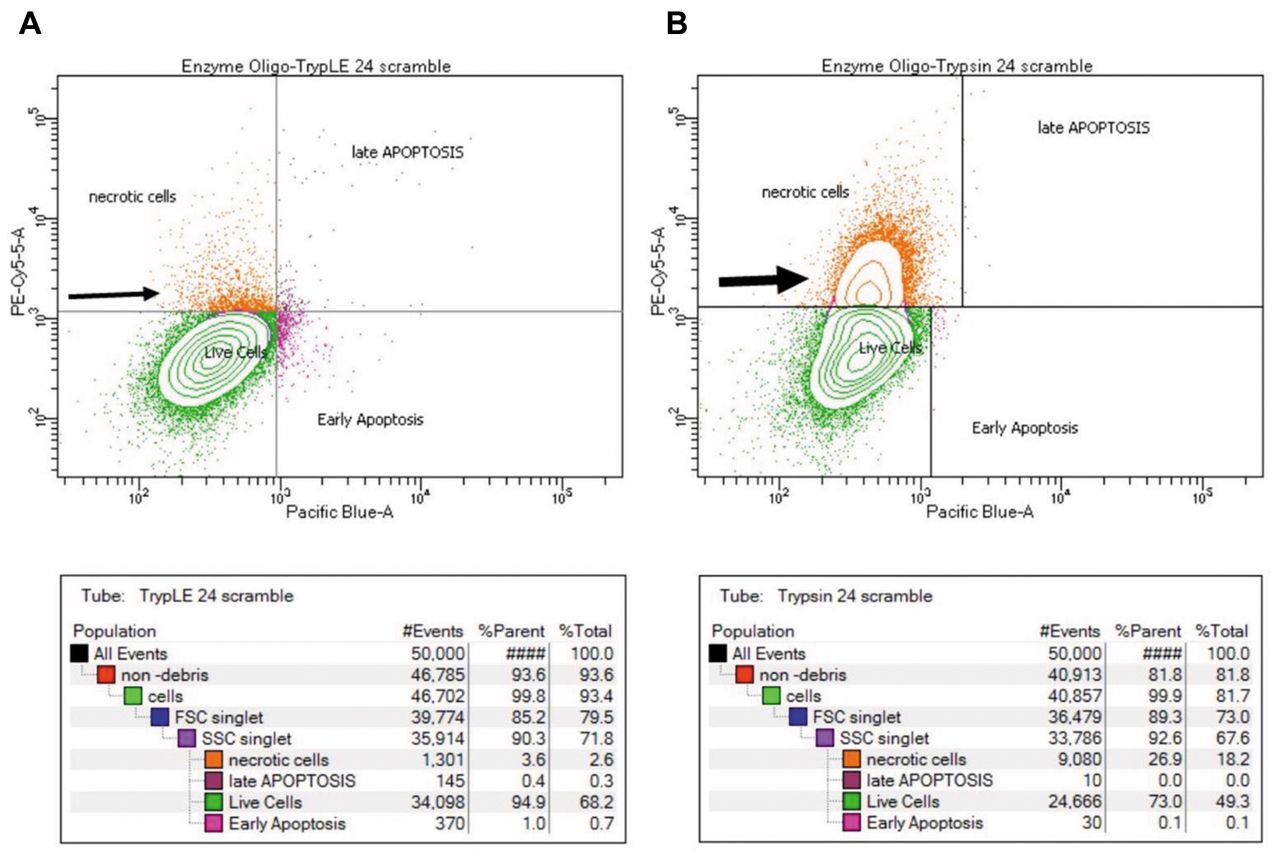
Representative flow cytometry plots after gating are displayed in Figure 2A and B and demonstrate the extent of cell membrane damage after harvesting with either TrypLE or Trypsin post transfection. The plots show a greater number of necrotic cells after harvesting with Trypsin (large arrowhead) compared with TrypLE (smaller arrow head). Review of the associated tables (seen below each plot) shows the percentage of necrotic, live, early apoptotic, and late apoptotic cells for each group. The panel indicates that harvesting with Trypsin yielded a higher percentage of necrotic cells at 23.1% and lower percentage of live cells (76.8%). Conversely, harvesting with TrypLE yielded a lower percentage of necrotic cells (4.1%) and higher percentage of live cells (95.2%).

Representative flow cytometry plots: Flow cytometry plots of transfected cells treated with (A) TrypLE and (B) Trypsin. (From left to right) Quadrant I corresponds to both positive Annexin V and 7-AAD staining of late apoptotic cells. Quadrant II is marked by positive 7-AAD staining of necrotic cells. Quadrant III denotes live cells negative for both Annexin V and 7-AAD staining. Quadrant IV shows only positive staining of Annexin V in early apoptotic cells. Tables below each plot show the population breakdown and % of parent cells.
To evaluate the significance of our observations, we first expressed cell viability as the percentage of parent live cells. By ANOVA, we identified a significant effect of the dissociation enzyme on cell viability [F(4, 10)=5.23, p<0.05] (Figure 3A). A Tukey’s Multiple Comparison test revealed that Trypsin treatment resulted in a significantly lower cell viability (78.77%±6.96) compared to TrypLE (95.67%±1.079) (p<0.01) and the Trypsin-DMEM control (91.83%±5.348) (p<0.05). Although we observed lower cell viability after dissociation via Accutase and Accumax, no significant difference was detected (Table I). Harvesting with Trypsin (CV=8.84%) exhibited the greatest variability while TrypLE (CV=1.13%) exhibited the least variability in percentage of viable cells between the three runs compared to the Trypsin-DMEM control (CV=5.82%), Accumax (CV=5.14%) and Accutase (CV=4.41%).

Cell viability and apoptosis detection following enzymatic cell dissociation. (A) TrypLE yielded the highest cell viability with the least variability between runs. Lower cell viability was observed with Trypsin compared to TrypLE and the Trypsin-DMEM control and also exhibited the greatest variability between runs. (B) Accutase yielded the most apoptotic cells compared to Trypsin and Trypsin-DMEM control. Data are presented as mean and SD. **p<0.01, *p<0.05.
Percentage of live, apoptotic and necrotic cells after harvesting.
To evaluate Annexin V staining, we expressed apoptotic cells as the percentage of parent late and early apoptotic cells. Using ANOVA, we found a significant effect of dissociation enzyme on the percentage of apoptotic cells [F(4, 10)=6.44, p<0.01] (Figure 3B). A Tukey’s Multiple Comparison test revealed that Accutase had a significantly greater percentage of apoptotic cells (1.73%±0.68) compared to Trypsin (0.07%±0.06) (p<0.01) and the Trypsin-DMEM control (0.53%±0.23) (p<0.05).
Discussion
Apoptosis is a common phenotypic endpoint following transfection of miRNA-based oligonucleotides. Annexin V/7-AAD staining combined with flow cytometric analysis is widely employed to detect apoptosis. Accurate apoptosis detection in this assay is contingent upon reducing confounding factors that may independently induce cell membrane damage. The impact of enzymatic harvesting on cell membrane integrity after transient transfection is unknown. Our goal was to evaluate the impact of four common dissociation enzymes -TrypLE, Accutase, Trypsin and Accumax-on cell viability, necrosis and apoptosis assessed via Annexin V/7-AAD flow cytometric analysis after miRNA-based transient transfection of laryngeal cancer cells.
Annexin V/7-AAD bind to cell membrane structures that have undergone morphological and biochemical changes associated with different stages of apoptosis; therefore, it is imperative that the cell membrane remain intact for accurate detection. Trypsin is a proteolytic dissociation enzyme that can lead to significant changes in cell membrane protein structure and composition. Accordingly, studies on non-transfected cancer and non-cancerous cell lines have shown that Trypsin dissociation diminishes cell surface antigen expression and induces metabolic leakage (16, 18). Authors have also shown variable cell viability results with Trypsin dissociation of non-transfected cancer and non-cancerous cell lines noting decreased cell viability in some cell lines and negligible effects in others (13-15). We showed, in transfected cells, that Trypsin significantly reduces cell viability likely related to cell membrane damage. We propose that increased cell membrane damage leads to an increase in 7AAD staining and reduced Annexin V staining in the assay, resulting in few to no apoptotic cells available for detection. Overall, although differences in cell lines may play a role, considering our findings, we cannot exclude the potential influence of miRNA-based transfection reagents on the cells.
Using nonsense oligos, several groups have noted the impact of transfection agents on cell viability and gene expression. Most reported that the agents decrease cell viability and alter gene expression, but in almost all instances, the enzymes used for dissociation were not mentioned in method sections. An example of this can be seen in a study by Böttger et al. (19), who compared the transfection reagents, INTERFERin, HiPERFect, and our reagent, Lipofectamine RNAiMAX in hepatocytes. The group reported that none of the transfection reagents caused cytotoxicity and lowered cell viability; however, they failed to mention the manner in which the cells were dissociated. Conversely, Sharma et al. compared the transfection reagents Lipofectamine, FuGene, and the process of nucleofection on cell viability, cell proliferation, and gene expression in buffalo fetal fibroblasts (11). The authors used Trypsin to detach transfected cells and found a significant decrease in cell viability and proliferation of transfected buffalo fetal fibroblast cells when compared to non-transfected trypsinized cells. In order to address these studies, we compared cell viability and necrosis in transfected and non-transfected trypsinized cells. We observed a significant difference in cell viability and necrosis where transfected trypsinized cells had a significantly lower number of viable cells and greater number of necrotic cells compared to non-transfected trypsinized cells. The significantly greater percentage of necrotic transfected cells versus necrotic non-transfected cells after Trypsin dissociation may indicate that the enzyme in conjunction with transfection agents can exacerbate membrane damage. Our study suggests that cell dissociation enzymes in conjunction with siRNA transfection agents alter cell membrane integrity in a human cancer cell line, which subsequently affects downstream Annexin V assay results in a manner that may be dissociation enzyme dependent.
It is important that we consider the interassay variability seen across dissociation enzymes. Evaluating the coefficient of variation for each enzyme, particularly for cell viability, we noted that TrypLE was the enzyme with lowest variability across studies (CV=1.13%), followed by Accutase (CV=4.41%), and Accumax (CV=5.14%) and then Trypsin (CV=8.84%). All of these CVs are considered acceptable, falling below 15%, but TrypLE’s is the least variable. TrypLE is considered a gentler alternative to Trypsin; it is less damaging to cells (mean cell viability of >95%), while having the lowest CV. Therefore, it provides the greatest reproducibility and reliability in our cell line. Collectively, these findings suggest that TrypLE is the most appropriate cell dissociation enzyme to use during cell harvesting for reliable Annexin V flow cytometric studies.
A limitation to the current study is the use of only one cell line to assess Annexin V results after enzymatic dissociation. It is possible that certain cell lines may be more or less susceptible to cell damage by different dissociation enzymes after transfection; however, the use of one cell line allowed us to focus more on the variation employed by each cell dissociation enzyme rather than the variation induced by different cell lines. It may be relevant that tumor origin, morphology, or grade of differentiation has not been found to contribute to susceptibility to cell damage by different harvesting methods (13). A strength of our analysis is the use of a nonsense oligo, which eliminated alterations in cell behavior secondary to the use of a specific oligo. Therefore, we are assured that our results are a consequence of the enzyme’s effect on our cells and not necessarily the influence of additional variables.
In conclusion, we report our analysis of the cell dissociation enzymes TrypLE, Accutase, Trypsin, or Accumax on Annexin V-7-ADD flow cytometric analysis after transfection. Our results show that Trypsin decreases the number of viable cells and increases the number of necrotic cells and as a result, cells are available for apoptotic analysis. We postulate that Trypsin in conjunction with transfection reagents may increase this phenomenon. TrypLE provides the greatest viability and least variability during harvesting. Overall, caution should be taken when selecting a method to dissociate adherent cells, as different methods can greatly compromise cell membrane structure and generate errors when detecting apoptosis after miRNA-based transient transfection. We also recommend, before assay use, pretesting cell dissociation enzymes on transfected cells to ensure reliable and reproducible results.
Footnotes
Authors’ Contributions
CG: designed and performed the experiments, analyzed the data, and wrote the manuscript. JM: performed experiments and contributed to writing of manuscript. AD: designed the experiments and analyzed the flow cytometric data. CL: contributed to experimental execution and revision of manuscript. KF: formulated the idea, designed the experiments, revised the manuscript, financial support of study.
Conflicts of Interest
The Authors declare no conflicts of interest in relation to this study.
- Received April 6, 2022.
- Revision received April 26, 2022.
- Accepted April 27, 2022.
- Copyright © 2022 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.