


Abstract
Background: Pancreatic cancer is the most lethal digestive cancer and the fourth overall cause of cancer death in the US. Asparagus, a widely consumed savory vegetable, is a rich source of antioxidants, saponins, vitamins, and minerals. In recent years, it has been shown that components of asparagus have anticancer effects on endometrial adenocarcinoma, and in prostate, breast, and colon cancer. In pancreatic cancer, it has been shown to have an anticancer effect on the KLM1-R cell line. This study was designed to investigate if asparagus extract (AE) had any effect on the growth of a widely used pancreatic cancer cell line MDAPanc-28 and to elucidate possible molecular mechanisms behind it. Materials and Methods: Clonogenic survival assay, proliferation, and caspase-3 activity kits were used to evaluate the effects of AE on cell survival, proliferation, and apoptosis pathway of MDAPanc-28 cells. We further investigated the possible molecular mechanisms by using reverse transcription-polymerase chain reaction. Results: The colony numbers and proliferation of MDAPanc-28 cells were surprisingly increased when treated with AE. The relative caspase-3 activity in cancer cells decreased when they were treated with AE. The pro-proliferative effect of AE on MDAPanc-28 cells correlated with down-regulation of anti-proliferative molecules P21 and P53. The potential anti-apoptotic effect of AE correlated with down-regulation of the pro-apoptotic molecule Fas cell surface death receptor (FAS) and down-regulation of caspase-3 activity. Conclusion: AE exhibits a pro-tumor effect on MDAPanc-28 pancreatic cancer cells by down-regulation of P21, P53, and FAS.
Pancreatic cancer (PC) is particularly dangerous since it is typically not diagnosed until the advanced stages of the disease. This is compounded by the fact that the overall 5-year relative survival rate is about 11%, and even lower at 3% when found at a later stage, with its incidence rising every year (1). However, this illustrates an increase compared to the 5-year relative survival rate of 10% in 2021 despite still being 3% when found at later stages, potentially indicating that treatments for early-stage PC are improving (2). Despite the possible improvements, an estimated 61,210 individuals are predicted to be diagnosed and 49,830 are expected to die from PC in 2022 (1). Due to the deadly nature of this cancer, it is a necessity that patients are provided with new and innovative therapeutic treatments. Surgical resection is currently the best treatment option, raising the 5-year relative survival rate to 15-25% (3). Despite having the best outcomes for patients with pancreatic cancer, the morbidity rates of these procedures remain high (3, 4). Neoadjuvant chemotherapy may be used prior to exercising surgical options and adjuvant chemotherapy can be continued afterward (5). Unfortunately, 75-85% of individuals diagnosed with PC do not have the option of a surgical route due to the progression of their disease, and chemotherapy and radiation therapy alternatives are explored instead (3, 5). Gemcitabine is a common chemotherapeutic option for PC. However, more modern treatments have come along in conjunction with gemcitabine that exhibit more effective results. Current cancer treatments have saved an estimated 3,495,700 lives since the late 20th century alone (1). It is important to pursue investigations that will continue this trend.
The cell line used in this study, MDAPanc-28, is a human pancreatic adenocarcinoma that was established in 1996. This cell line expresses trypsin and carbonic anhydrase II (6). The α2,3-sialyltransferases, ST3Gal III and ST3Gal IV, are overexpressed in MDAPanc-28, which allows for the synthesis of sialyl-Lewis(x), resulting in the adhesive and migratory characteristics of this cell line (7).
Asparagus contains many different biologically active compounds that have been shown to assist in the reduction of proliferation and induction of apoptosis of various cancer cells. Some of these compounds include rutin, different types of saponins, and sulforaphane (8-10). In other recent studies showing the effect of extracts from various species of asparagus on cancer, there appears to be a broad anticancer effect, usually inhibiting proliferation, inducing apoptosis, and having a cytotoxic effect depending on the tumor cell line (9, 11-14). Some studies have even investigated the use of AE as a potential therapy option to accompany current cancer treatments (15-17). Aside from cancer, saponins found in asparagus have also been shown to inhibit enzymes which might prove to be helpful in treatment for those with Alzheimer’s disease (18).
Our laboratory has conducted many studies on the effects of fruit and vegetable extracts, in addition to cytokines, on cancer cell lines to date (19-23). The investigation of these extracts and cytokines as potential radiosensitizers has also been explored in parallel (24-28). Of our more recent findings, kiwifruit extract was shown to have an anti-proliferative and pro-apoptotic effect as a radiosensitizer against CRL-11147 melanoma cells (29). Additionally, the cytokine, interleukin-39, was found to halt growth and increase apoptosis of the T24 bladder cancer cell line (30). It would be interesting to expand our studies about the effect of agents from vegetables and fruits on the growth of cancer. In pancreatic cancer, AE has been shown to have an anticancer effect on the KLM1-R cell line (13). However, little is known about its effects on the MDAPanc-28 pancreatic cancer cell line. Contrary to other currently used human pancreatic cancer cell lines, MDAPanc-28 displays both acinar and ductal traits (6). This unique characteristic makes the cell line especially important to study. This study was designed to investigate if AE had any effect on the growth of MDAPanc-28 pancreatic cancer cells and elucidate possible molecular mechanisms behind it.
Materials and Methods
Tumor cell line. The human MDAPanc-28 cell line used in this study was provided by Dr. Citrin from the Radiation Oncology Branch of the National Institutes of Health (Center for Cancer Research, National Cancer Institute, MD, USA). MDAPanc-28 cells were cultured in Dulbecco’s modified Eagle’s medium (Invitrogen, Carlsbad, CA, USA). A solution of 10% heat-inactivated fetal bovine serum and 1% penicillin-streptomycin (Invitrogen) were used to supplement the medium. Treatment options were performed once MDAPanc-28 cells reached 70% confluence.
Treatment of MDAPanc-28 with AE. Once at 70% confluence, MDAPanc-28 cells were treated with AE at a concentration of 50 μg/ml for 72 hours. The optimal concentration for the experimental treatment was determined by previous studies (28, 31-33). Organic Asparagus 50% Saponins Extract (Badmonkey Botanicals, Tacoma, WA, USA) was used as the AE. The control consisted of MDAPanc-28 cells in medium only for the same amount of time.
Clonogenic survival assay. The clonogenic survival assay was completed as described elsewhere (34). MDAPanc-28 cells were harvested from their culture flasks using TrypLE Express (Invitrogen) then suspended in phosphate-buffered saline and were counted by a hemocytometer. The clonogenic survival assay was performed by plating 1,000 cells onto 60-mm Petri dishes in triplicate and incubating them at 37°C in a humidified incubator with 5% CO2. On day five after seeding, fresh medium was added. After 9 days of incubation, the MDAPanc-28 cells were fixed with 10% formaldehyde and stained with 0.05% crystal violet. The number of AE-treated colonies were counted, averaged, and expressed as a percentage of the total colonies in the control (untreated).
Measurement of caspase-3 activity. The test for the biological activity of the apoptotic marker, caspase-3, in MDAPanc-28 cells was completed with a caspase-3/CPP32 colorimetric assay kit (BioVision, Waltham, MA, USA) which has been described previously (35). The results for AE-treated cells were normalized to these of the untreated controls.
Determination of proliferation with Quick Cell Proliferation Assay Kit. In order to further determine cell proliferation, a quick cell proliferation assay kit (BioVision) was used according to the manufacturer’s instructions. Cell content of the samples was determined by measuring metabolic activity reflected by the production of formazan dye by spectroscopy (36). The results for AE-treated cells were normalized to these of the untreated controls.
Reverse transcription-polymerase chain reaction. AE-treated and control MDAPanc-28 cells were initially washed with phosphate-buffered saline and then homogenized by TRIzol (Invitrogen). Once the mRNA was extracted, the concentrations of each sample were determined by NanoDrop (Thermo Fischer Scientific, Waltham, MA, USA). For the treatment and control conditions, 1 μg of mRNA was reverse transcribed as previously reported (34). To ensure that the amount of amplified mRNA was normalized, concentrations of the housekeeping gene, glyceraldehyde 3-phosphate dehydrogenase, were also recorded. The relative mRNA concentrations were determined by gel electrophoresis in 2% agarose gel and staining with ethidium bromide. UV light was then used to visualize the staining using an IS-1000 digital imaging system (Life Sciences, St. Louis, MO, USA). The amount of mRNA present in the control and AE treated cells were normalized relative to the mRNA concentration of glyceraldehyde 3-phosphate dehydrogenase. The RNA primer sequences used in this study are listed in Table I.
RNA primers used for reverse transcription-polymerase chain reaction.
Statistical analysis. Each experiment was performed at least three times. Statistical analysis of the data was performed using an unpaired two-tailed t-test. A p-value of less than 0.05 was used to denote statistical significance.
Results
AE induces proliferation of MDAPanc-28 cells. The purpose of this research was to study the effect of AE on the proliferation and growth of MDAPanc-28 cells. As observed in the data, AE treatment increased proliferation and survivability of the MDAPanc-28 cell line when compared to the untreated controls. The results of the clonogenic survival assay and quick cell proliferation assay showed that when treated with AE, there was both a significant increase (p<0.05) in the percentage of colonies and the optical density relative to the control (Figure 1).

Effects of 50 μg/ml asparagus extract (AE) on colony-forming ability and proliferation of MDAPanc-28 cells. The experiment was performed in triplicate and as described in the Materials and Methods section. The graphs shown represent the mean values for the percentage of colonies formed and the optical density (OD) of AE-treated cells relative to the control (Cx) cells. Bars indicate the standard deviation of each mean. *Significantly different from the control at p<0.05.
AE down-regulates the expression of cell-cycle-related proteins. The effect of AE on the mRNA expression of cyclin-dependent kinase inhibitor 2C (P18), cyclin-dependent kinase inhibitor 1A (P21), cyclin-dependent kinase inhibitor 1B (P27), tumor protein 53 (P53), cyclins B1, D1, and E1, as well as cyclin-dependent kinase 2 (CDK2) and cyclin-dependent kinase 4 (CDK4), was analyzed. Expression of both P21 and P53 significantly decreased (p<0.05) compared to the control when MDAPanc-28 cells were treated with AE (Figure 2). Additionally, there was a significant decrease (p<0.05) in CDK4 expression when compared to the control (Figure 2). This created an interesting contradiction in the results considering the role of CDK4 in progression of the cell cycle. This contradiction is further explored in the Discussion. These results suggest that the down-regulation of P21 and P53 is correlated with the increased proliferation seen in the AE-treated MDAPanc-28 cell line.

Relative mRNA expression of the anti-proliferative proteins, cyclin-dependent kinase inhibitor 2C (P18), cyclin-dependent kinase inhibitor 1A (P21), cyclin-dependent kinase inhibitor 1B (P27), tumor protein 53 (P53), cyclins B1, D1, and E1, as well as cyclin-dependent kinase 2 (CDK2) and cyclin-dependent kinase 4 (CDK4), were determined in MDAPanc-28 cells treated with 50 μg/ml asparagus extract (AE) and in untreated control (Cx) cells. The experiment was performed in triplicate and as described in the Materials and Methods section. The graphs represent the mean mRNA expression of each gene relative to the mRNA expression of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) analyzed by reverse transcription-polymerase chain reaction. Bars indicate the standard deviation of each mean. *Significantly different from the control at p<0.05.
AE down-regulates the expression of apoptosis-related genes Fas cell surface death receptor (FAS). Finally, we looked at the mRNA expression of anti-apoptotic proteins FAS, Fas ligand (FASL), TNF receptor superfamily member 10a (TRAILR1), TNF superfamily member 10 (TRAIL), CASP8 and FADD like apoptosis regulator (FLIP), BCL2 apoptosis regulator (BCL2), and survivin (baculoviral IAP repeat-containing 5, BIRC) in the AE-treated and control cells. The relative caspase-3 activity was shown to significantly decrease (p<0.05) with AE treatment when compared to the control using a caspase-3 activity kit (Figure 3). Surprisingly, considering the high survivability of this cell line, the only apoptotic gene that was significantly down-regulated (p<0.05) was FAS (Figure 4). Both these results helped propose a correlation between the down-regulation of FAS and the potential reduction of apoptosis seen in the AE-treated MDAPanc-28 cell line. These effects may have had a role in the increased survival of the cell line that was observed.

Relative caspase-3 activity in MDAPanc-28 cells treated with 50 μg/ml asparagus extract (AE) and in untreated control (Cx) cells. The experiment was performed in triplicate and as described in the Materials and Methods section. Data are the mean plus the standard deviation. *Significantly different from the control at p<0.05.
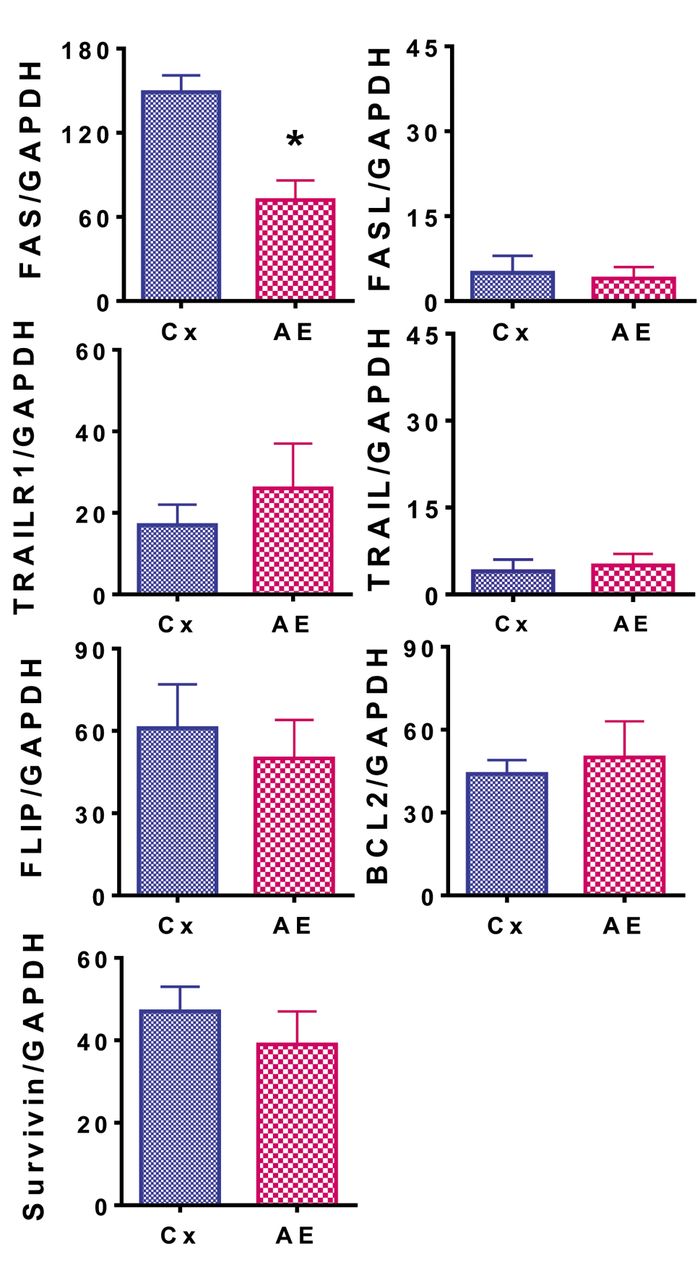
Relative expression of apoptosis-related genes, Fas cell surface death receptor (FAS), Fas ligand (FASL), TNF receptor superfamily member 10a (TRAILR1), TNF superfamily member 10 (TRAIL), CASP8 and FADD-like apoptosis regulator (FLIP), BCL2 apoptosis regulator (BCL2), and survivin (baculoviral IAP repeat-containing 5, BIRC), were observed in MDAPanc-28 cells treated with 50 μg/ml asparagus extract (AE) and in untreated control (Cx) cells. The experiment was performed in triplicate and as described in the Materials and Methods section. Data are the mean plus the standard deviation of expression for each gene relative to that of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) analyzed by reverse transcription-polymerase chain reaction. *Significantly different from the control at p<0.05.
Discussion
Pancreatic cancer is the deadliest digestive cancer, being the fourth largest contributor to cancer deaths in the United States. AEs from different species containing various antioxidants, saponins, vitamins and minerals, have been shown to have a widely anticancer effect on various tissue types and cell lines. Despite this, with only 13 publications to date, the relatively understudied pancreatic cancer cell line, MDAPanc-28, has been shown to defy this general trend by displaying increased proliferation and survival in the presence of AE. Known for their anti-proliferative effects, the mRNA expression of P21 and P53 were found to be down-regulated in the presence of AE. More in line with the traditional effects of AE on cancer cells, the cell-cycling protein CDK4 was shown to be down-regulated. When compared to the medium-only controls, AE increased the percentage of colonies as well as optical density, reflecting an increase in proliferation and survival of this cell line. As far as we are aware, this is the first study looking into the effects of AE on the MDAPanc-28 cell line. The stark deviance from the previously seen effects of AE on other cancer cell lines denotes the clinical significance of this study.
The vast majority of studies focusing on the effect of AE on cancer found it to have anticancer effects. A study on breast, colonic, and pancreatic cancer cell lines, revealed that AE inhibited cell proliferation by the up-regulation of ras homolog family member A (RHOA) and the down-regulation of Rac family small GTPase 1 (RAC1) in the Rho GTPase pathway (37). In a study investigating the interactions between AE and the hepatocellular cancer cell line, HepG2, in addition to the down-regulation of CDK4, there was a significant increase in P21 mRNA expression, while P53 did not statistically differ (38). A study on the colorectal cancer cell line, HCT-116 presented an increase in P53 mRNA expression in response to AE (39). When AE was used on the MDA-MB231 and MCF7 breast cancer cell lines, it resulted in cell-cycle arrest between the G1 and S stages (40). Despite not looking into specific mRNA concentrations in that study, the cell-cycle arrest between the G1 and S stages seems to suggest the down-regulation of CDK4, a seemingly common effect of AE cancer treatments. Our study also showed this down-regulation of CDK4, but additionally presented the down-regulation of P21 and P53, which is contradictory to the effects of AE on other cancer cell types. The stark difference in P21 and P53 expression in MDAPanc-28 cells compared to other cell lines when treated with AE should be studied further.
The use of AE as a potential chemosensitizer is an important avenue to explore. Future studies on the effects of AE as a chemosensitizer in the MDAPanc-28 cell line may yield interesting and beneficial results considering the pro-cancer effect of AE on the cell line alone. In the KLM1-R pancreatic cancer cell line it was found that AE treatments down-regulated heat-shock protein 27 (13). Heat-shock protein 27 was also shown to be involved in gemcitabine resistance of pancreatic cancer cell lines (41). If the chemosensitizing effects of AE can overcome its proliferative and potential anti-apoptotic effects in the MDAPanc-28 cell line, it could prove to become a very important tool in future pancreatic cancer treatments. As mentioned earlier, one major difference between MDAPanc-28 and many other human pancreatic cancer cell lines is that it displays both acinar and ductal traits. However, most of the common pancreatic cell lines used in research are exclusively ductal. Acknowledging this distinction may help yield better treatments for pancreatic cancer displaying this characteristic.
The proteins P21 and P53 are both known for their antiproliferative effects in cellular biology. As shown in the data of this study, mRNA expression of both P21 and P53 was significantly reduced when MDAPanc-28 cells were treated with AE. This was accompanied by a significant increase in colony formation and proliferation when treated with AE. It has been shown that both P21 and P53 work cooperatively to free BCL2-associated X, apoptosis regulator (BAX) from BCL2 complexes (42). Once liberated, BAX can exert pro-apoptotic effects on the cell. With both P21 and P53 down-regulated in response to AE treatments, this decreases the likelihood for the liberation of BAX, therefore reducing apoptosis and allowing for increased proliferation. Our data revealed that BCL2 mRNA concentrations in the MDAPanc-28 cell line did not statistically significantly change with AE treatment. Due to this, our results for BCL2 likely did not affect this mechanism.
One of the most well-known pro-apoptotic proteins in cellular biology is FAS. Found in the external plasma membrane, FAS is activated by FASL. This activation leads to signal transduction and programmed cell death. As shown in this study, there was a significant decrease in FAS mRNA expression when AE treatment was used and no statistically significant difference in FASL expression. In a study on the hepatocytes of mice, it was found that interferon-γ (IFNγ)-null mice (Ifng−/−) showed lower levels of FAS mRNA expression than Ifng+/− mice (43). It is possible that AE affects IFNγ expression or proteins further down its transduction pathway, such as signal transducer and activator of transcription 1 (STAT1), causing a reduction in FAS mRNA expression. To our knowledge, no studies have been performed on the effect of asparagus on IFNγ in reference to cancer biology, or STAT1. Additionally, in a recent study on the colorectal cancer cell line, HCT-116, AE was shown to up-regulate FAS expression, displaying a completely different effect from those shown by our data (38). The effect of AE on these proteins warrants further study.
Our data collected on CDK4 represents a contradiction in the proliferative characteristics of this cell line. However, as discussed earlier, the down-regulation of CDK4 is a common outcome of AE treatment. CDK4 is involved in the progression from the G1 phase to the S phase of the cell cycle (44). The inhibition of CDK4 can be used as a method of therapy for cancer due to this effect in halting cell growth and proliferation (45). Therefore, its down-regulation in the presence of AE would hint at anti-proliferative effects, however, the data declare otherwise. It is likely that the proliferative effects of down-regulating P21 and P53 were able to outweigh the antiproliferative effects of down-regulating CDK4. It is important to question what potentially aggressive levels of proliferation this cell line might have achieved when coupled with AE treatment if the CDK4 level had remained unchanged. There is clinical significance in the possibility of using AE to enhance the effectiveness of CDK4 inhibitors as a cancer treatment. Further studies on the interplay between CDK4 mRNA expression and AE treatment in the MDAPanc-28 cell line might help to elucidate mechanisms, aiding studies like those by Goel et al. focused on using CDK4 inhibitors as a method of cancer treatment (45).
There are limitations present in this research. Only one cell line was studied, although our laboratory is actively studying other pancreatic cancer cell lines and will address the effect of AE on them in separate articles. Studies comparing the effect of AE on multiple pancreatic cancer cell lines may assist in finding the mechanisms of action. Additionally, this study only reflects the in-vitro effects of AE on the MDAPanc-28 cell line. However, the study is being expanded to other cell lines, in addition to animal studies, in order to further address the effect of AE on pancreatic cancer.
In summary, in this study, AE exhibited a pro-tumorous effect on MDAPanc-28 pancreatic cancer cells, with the down-regulation of P21, P53, and FAS in vitro. Despite the increase in proliferation and potential decrease in apoptosis, the action of AE in down-regulating CDK4 might be potentially useful in conjunction with CDK4 inhibitors as a cancer treatment. There is currently little known about the interactions between AE and pancreatic cancer. This study expands the breadth of knowledge on the topic and may assist in developing new and innovative treatment options.
Acknowledgements
This study was partially supported by a grant from Des Moines University for Yujiang Fang, M.D., Ph.D. (IOER 112-3749).
Footnotes
Authors’ Contributions
Yujiang Fang designed the study. Yujiang Fang, Huaping Xiao, Jacob T. Hough, Xuhui Chen, Jacob Lee, Aldo Dominguez, Tianru Shi, Joseph Schmidt, and Qian Bai performed the experiments. Yujiang Fang, Zuliang Deng, and Mark R. Wakefield analyzed the data. Yujiang Fang, Zuliang Deng, Huaping Xiao, Mark R. Wakefield, and Jacob T. Hough interpreted the data. Jacob T. Hough wrote the draft and Yujiang Fang revised the article critically.
Conflicts of Interest
The Authors declare that they have no conflicts of interest.
- Received March 1, 2022.
- Revision received April 3, 2022.
- Accepted April 6, 2022.
- Copyright © 2022 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.