


Abstract
Background/Aim: Carboxyl terminus of Hsc70-interacting protein (CHIP) is a ubiquitin ligase that induces ubiquitination and degradation of its target proteins including oncoproteins. We reported that its down-regulation is associated with tumor progression and metastasis of breast cancer. However, the mechanism through which CHIP gene affects cancer cells is unclear. Materials and Methods: We extracted RNA from 45 primary breast cancer samples and compared CHIP mRNA expression profiles, promoter DNA methylation status, and clinicopathological information. Results: CHIP mRNA expression was significantly correlated with the tumor progression status. In several samples, a pinpoint CpG methylation in the CHIP gene promoter region was significantly correlated with CHIP mRNA expression. When this specific CpG was methylated in estrogen receptor (ER)-positive cases, a significant difference in 5-year recurrence was not found compared with ER-negative cases. Conclusion: CpG methylation contributes to the long-term prognosis of ER-positive breast cancer.
Recent studies have shown that the ubiquitin-proteasome pathway plays an important role in the regulation of tumor properties (1-3). This pathway controls selective degradation of short-lived proteins including many oncoproteins. Proteins targeted for proteasomal degradation are sequentially tagged with ubiquitin by ubiquitinating enzymes E1, E2, and E3. In particular, E3 ubiquitin ligases play a pivotal role in ubiquitination. They catalyze the linkage of ubiquitin to the substrate protein, and polyubiquitinated substrates are degraded by the proteasome. E3 ubiquitin ligases have been studied extensively in the field of cancer, and some of them have emerged as therapeutic targets.
Carboxyl terminus of heat shock cognate (Hsc) 70-interacting protein (CHIP) is a U-box-type E3 ubiquitin ligase (4, 5). CHIP interacts with Hsc70/90 and is involved in ubiquitination and degradation of its target proteins including several oncoproteins (6-8). In breast cancer, CHIP plays a key role in degradation of misfolded estrogen receptor alpha (ERα) and human epidermal growth factor receptor 2 (HER2) (9-11). Recently, we reported that down-regulation of CHIP is significantly associated with tumor progression and metastasis of human breast cancer (12). We found that CHIP expression levels are markedly down-regulated in advanced tumor samples, such as node metastasis-positive and late-stage cancers, compared with normal samples. Thus, CHIP is closely associated with the malignancy of breast cancer, but the mechanism that regulates its mRNA expression is unclear. To elucidate this mechanism, we focused on a representative gene regulatory mechanism, DNA methylation of the gene promoter region. It is well known that epigenetic regulation such as DNA methylation plays crucial roles in mRNA expression (13-15). Aberrant DNA methylation of CpG islands in gene promoter regions leads to silencing of several important genes including tumor suppressor genes, and the CHIP gene has a CpG island in its promoter region. Therefore, in this study, we analyzed the DNA methylation status of the CHIP gene promoter region and compared it with its mRNA expression levels among 45 breast tumor samples. Moreover, to confirm whether the DNA methylation status is associated with clinicopathological characteristics and prognosis, we compared the methylation status with the clinicopathological characteristics and recurrence rate of patients.
Materials and Methods
Tumor samples. Primary human breast cancer tissues were obtained during surgery from 45 patients after patient consent from 1999 to 2001 at Saitama Cancer Center Hospital (Saitama, Japan), embedded in optimal cutting temperature (OCT), and transferred to Tohoku University. The study was approved by Tohoku University Ethics Committee (Approval No. 2009-81). The estrogen receptor (ER) and progesterone receptor (PgR) protein expression in the samples were determined by enzyme immunoassays. The cut off value for ER and PgR was set at 10 units.
RNA preparation and real-time RT-PCR. Several 5 μm-thick sections were prepared from tumor tissues embedded in OCT and mounted on film-coated glass slides. Then, each section was fixed, dehydrated with methanol, and stained with toluidine blue staining prior to laser capture microdissection (LCM). The sections were also stained with hematoxylin-eosin to determine the outlines of tumor areas. Only outlined tumor areas were microdissected using a Leica LMD7000 (Leica Microsystems GmbH, Wetzlar, Germany). RNA was extracted from the collected tumor cells using an RNeasy Micro Kit (Qiagen, Mississauga, Canada), according to the manufacturer’s instructions. Total RNA was converted to first-strand cDNA using a QuantiTect Reverse Transcription Kit (Qiagen). Real-time PCR was performed using a StepOne™ Real-Time PCR System (Applied Biosystems Inc., Foster City, CA, USA). The oligonucleotides used in real-time PCR were as follows: forward primer 5’-GGA GAG CTA TGA TGA GGC CAT C-3’, reverse primer 5’-AAG TTC AGC CGC TGC TCC T-3’, and probe 5’-CTG CAG CGA GCT TAC AGC CTG GC-3’ for CHIP; forward primer 5’-GAG TAC TTG CGC TCA GGA GGA-3’, reverse primer 5’-ACG TGG ACA TCC GCA AAG A-3’, and probe 5’-CAA TGA TCT TGA TCT TCA TTG TGC TGG GTG-3’ for β-actin as the internal control.
DNA preparation and bisulfite sequencing. Several 40 μm-thick sections containing a large tumor fragment were prepared from tissues embedded in OCT. DNA from these tumor samples was extracted using a QIAamp DNA Mini Kit (Qiagen), according to the manufacturer’s instructions. Extracted DNA was treated with bisulfite using an EpiTect Bisulfite Kit (Qiagen), according to the manufacturer’s instructions. Then, nested PCR was carried out at 95°C for 2 min, followed by 42 cycles (40 cycles for 2nd PCR) at 95°C for 30 s, 48°C for 30 s (1 min for 2nd PCR), 72°C for 1 min, and final extension at 72°C for 7 min. The oligonucleotides used in nested PCR were as follows: 1st PCR forward primer 5’-TTG GAT TTA TTA GGG AGG TT-3’, 2nd PCR forward primer 5’-TTA AGT TGT TAG GTT AGT AG-3’, and common reverse primer 5’-CTA AAC TAC CAT TCT AAA AC-3’. The PCR amplicon was electrophoresed, and DNA from the correctly sized band was extracted using a QIAquick Gel Extraction Kit (Qiagen). The purified DNA was sequenced using an ABI PRISM 3100 Genetic Analyzer (Applied Biosystems Inc.).
Statistical analysis. Statistical analysis was performed with the StatFlex 6.0 software program (Artech Co., Ltd., Osaka, Japan). Comparison of two independent groups was performed with the Mann-Whitney U-test. For comparison among three groups or more, the Kruskal-Wallis test was used. p<0.05 was considered to be statistically significant.
Results
CHIP mRNA expression significantly correlates with tumor progression of breast cancer tissue. To examine the role of CHIP gene in breast cancer, we assessed the relationship between the CHIP mRNA expression and clinicopathological characteristics of the cases (Table I). CHIP mRNA was significantly decreased with lymph node metastasis and stage progression. Other factors related with tumor progression such as the tumor size and TNM status tended to be associated with lower CHIP mRNA expression although without statistical significance. The CHIP gene is an ER target with an ERE half site in its promoter region, but CHIP mRNA expression was not significantly correlated with expression of ER protein or its target gene PgR. These results suggest that another transcription activation mechanism regulates the expression of CHIP gene in addition to ER-dependent transcription.
Relationship between CHIP mRNA expression and clinicopathological factors.
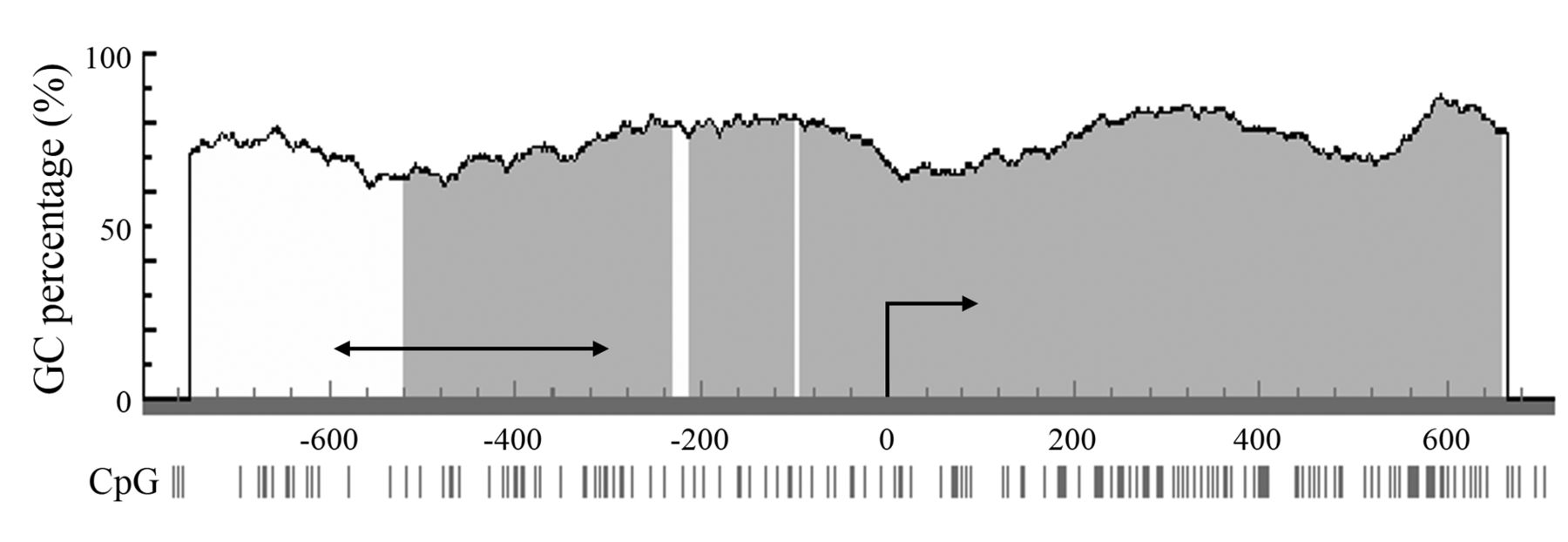
Promoter methylation of the CHIP gene regulates its expression. We hypothesized that down-regulation of CHIP expression in breast cancer occurred through promoter methylation as a representative epigenetic regulation system. Therefore, bisulfite-specific PCR was performed using DNA extracted from the samples, and the DNA methylation status was determined by analysis of the sequence of the PCR product. The analyzed region was 249 bp from -600 bp to - 352 bp of the transcription initiation site in the CHIP gene (Figure 1). This region was identified as a domain containing the region closely related to transcriptional activation using a luciferase reporter assay in preliminary experiments (data not shown). Results from the methylation analysis of the 38 breast cancer tissues indicated that methylation occurred in the 4th to 9th CpG sites of the region in 50% (19/38) of the samples (Figure 2). Samples with no less than 50% (three point) methylation of these six CpG sites were designated as positive for methylation. Using this threshold, samples were divided into methylation (M) and unmethylation (U) groups. We then compared the methylation status of M and U groups with the CHIP mRNA expression status (Figure 3A). Although there was no statistical significance, the methylation status seemed to correlate with CHIP mRNA expression. Interestingly, there were six samples with only the 9th CpG (CpG9) unmethylated in the M group, whereas one sample only had CpG9 methylated in the U group (Figure 2). Therefore, we divided the samples into CpG9 M and CpG9 U groups and compared the CpG9 methylation status with the CHIP mRNA expression status (Figure 3B). Statistical analysis revealed significant intergroup differences, namely CHIP mRNA expression was significantly decreased in the CpG9 M group compared with the CpG9 U group. However, no significant difference in CHIP mRNA expression was observed between groups with other CpG pinpoint methylations (data not shown).

Analyzed region in the CHIP gene promoter. The region analyzed by bisulfite DNA sequencing was 249 bp from -600 bp to -352 bp of the transcription initiation site in the CHIP gene. The horizontal arrow indicates the sequence of the analyzed region, and the curved arrow indicates the transcription initiation site. The gray area indicates a CpG island.

DNA methylation status of the CHIP gene in breast tumor tissues. (Left) The region analyzed by bisulfite DNA sequencing included 17 CpG sites. White circles indicate unmethylated CpG sites, and black circles indicate methylated CpG sites. Samples with no less than 50% (three point) methylation in the 4th to 9th CpG sites were designated as the region-methylated group. (Right) CHIP mRNA expression levels of four groups divided by region methylation and the CpG9 methylation status. Black bars are the region-methylated/CpG9-methylated group. Gray bars are the region-methylated/CpG9-unmethylated group. Light gray bars are the region-unmethylated/CpG9-methylated group. White bars are the region-unmethylated/CpG9-unmethylated group.

CHIP mRNA expression level classified by the methylation status in patients. The intergroup difference of CHIP mRNA expression in patients divided into methylated or unmethylated groups is shown. Box plots show the intergroup difference of the (A) CpG4-9 and (B) CpG9 methylation status in each group.
DNA methylation coordinately regulates CHIP mRNA expression with ER. As shown in Table I, the ER status was not correlated with CHIP mRNA expression, even though CHIP is an ER target gene. It was assumed that this pinpoint CpG9 methylation interfered with the mechanism through which ER regulates CHIP mRNA expression. Therefore, ER-positive and -negative groups were classified into M and U groups, and CHIP mRNA expression of the four groups was compared (Figure 4). Interestingly, remarkable down-regulation of CHIP mRNA expression was observed only in the ER-negative M group. In contrast, the other three groups maintained high mRNA expression, especially the ER-negative U group (p<0.05). These results suggest that CHIP gene is complimentarily regulated by ER and DNA methylation. Specifically, the reason that there was no difference in ER-positive and -negative groups was CpG methylation.

CHIP mRNA expression level classified by the methylation status and ERα in patients. The intergroup difference of CHIP mRNA expression in patients divided into methylated or unmethylated and ER-positive or -negative groups is shown. Box plots show the intergroup difference of the CpG9 methylation status in each group.
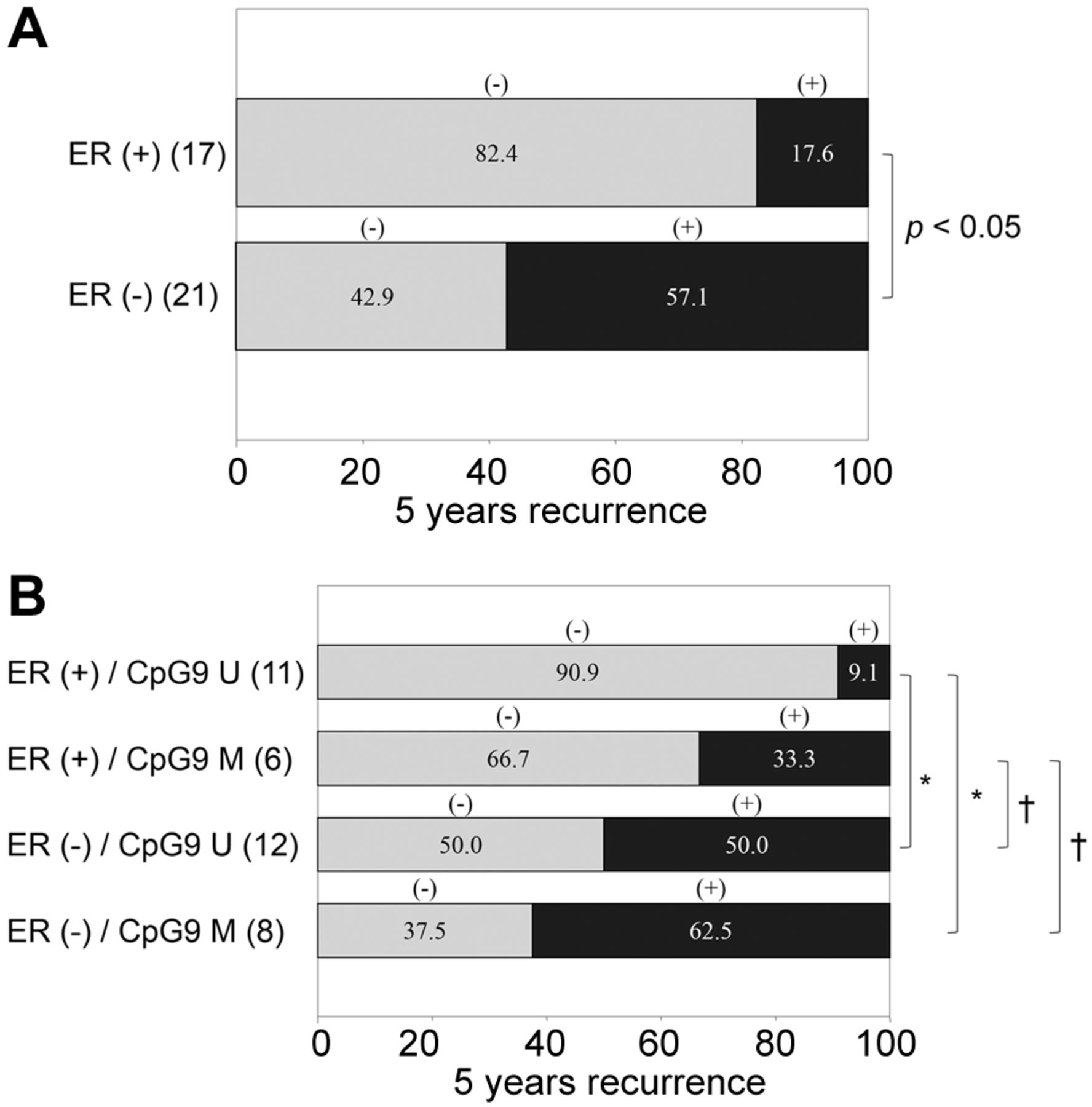
CpG9 methylation of the CHIP gene predicts long term recurrence. CHIP expression suppresses distal metastasis (12). Therefore, we investigated whether CpG9 methylation-regulated expression was associated with tumor recurrence after 5 years. Although the number of patients with recurrence in the ER-positive group was significantly lower than that in the ER-negative group, some metastatic recurrence was observed in patients despite ER positivity (Figure 5A). In the analysis of the four groups divided by ER positivity and CpG9 methylation (Figure 5B), the recurrence rate in the ER-positive/GpG9 U group was significantly lower than that in the two ER-negative groups. However, in the ER-positive/CpG9 M group, there was no significant difference in the recurrence rate compared with the two ER-negative groups. These results suggest that ER-positive breast cancer can be divided in two prognostic groups by the CpG9 methylation status. Thus, ER and methylation status may be valuable for prognosis prediction.

Recurrence rate after 5 years in patients divided by ERα expression and CpG9 methylation. The intergroup difference of the recurrence rate after 5 years in patients divided into ER positive/negative and CPG9-methylated/unmethylated groups is shown. Bars indicate the intergroup difference of (A) only ERα, and (B) ERα and CpG9 methylation in each group. Gray bars indicate the non-recurrence group, and black bars indicate the recurrence group. *p<0.05, †N.D.
Discussion
When determining a therapeutic strategy for breast cancer, the resistance mechanism is one of the most important considerations. ER is the factor that relates most strongly to breast cancer proliferation and progression, and is used to predict the efficacy of hormonal therapy (16-20). Generally, although ER-positive breast cancer is considered to have a good prognosis compared with ER-negative breast cancer, it is not necessarily an indicator of long-term recurrence. According to the U.S. patient registration database in 1992-2007 (21), ER-negative cases have a peak recurrence rate at an early stage compared to 1-3 years postoperatively, and the rate decreases over time. However, ER-positive cases maintain a constant rate of recurrence in the long term, which is higher than the rate of ER-negative cases at 5 years postoperatively. According to reports of the recurrence risk in many countries, the recurrence risk of ER-positive breast cancer exceeds that of ER-negative breast cancer at 3-4 years postoperatively (21). It is necessary to follow-up patients ER-positive breast cancer for several years postoperatively for recurrence. Therefore, new additional indices are required for ER-positive breast cancer.
CHIP is an ER target gene with an important role in breast cancer (4-12). CHIP mRNA expression in clinical samples was negatively correlated with tumor progression, especially lymph node metastasis and advanced stage. We have reported that CHIP promotes degradation of SRC-3 (12), an important coactivator of ER (22-25), as a target of ubiquitination in breast cancer. SRC-3 elevates the transcriptional activity of Smad2 and expression of Twist1, and causes tumor metastasis (12, 26). Additionally, SRC-3 causes resistance to endocrine therapy in primary breast cancer by targeting phosphorylation pathways such as HER2 (27, 28). Similar to previous reports, in our samples, SRC-3 degradation may have been associated with repression of tumor progression and metastasis. It has been reported that CHIP expression leads to regression of cancer. Although CHIP is a target gene of ER, its expression was not correlated with ER expression in previous reports or this study. Thus, there may be mechanisms regulating its expression other than those mediated by ER.
To delineate the expression mechanism, we focused on the epigenetic regulation of the CHIP gene and analyzed the DNA methylation status of its promoter region in clinical samples. Methylation was identified in the 4th to 9th CpG sites in 50% of the samples. Furthermore, methylation of the 9th CpG in this region was significantly correlated with down-regulation of CHIP mRNA expression. Analysis of transcription factor binding sites in this domain revealed binding of c-Ets, c-Myb, and c-Rel, which are methylation-sensitive transcription factors. If CpG islands are methylated, binding of these transcriptional factors is inhibited by the methyl group itself or a methyl CpG-binding protein, and transcriptional activity is down-regulated (29, 30). Therefore, methylation transcription factor recognition sequence in this region may be a mechanism through which CHIP expression is regulated.
CHIP mRNA expression did not correlate with expression of ER protein, a known CHIP genetic control factor (12). To investigate the mechanism that regulates transcription of the CHIP gene independently of ER, the samples were divided into four groups by positivity for the ER and CpG9 methylation. High expression of CHIP mRNA was observed in the CpG9 U group regardless of ER expression. Moreover, in the CpG9 M group, down-regulation of CHIP mRNA was observed only in the ER-negative group. Because CHIP mRNA was highly expressed except in the ER-negative/CpG9 M group, CHIP gene expression may be regulated by two mechanisms employing ER and/or CpG9 methylation. Furthermore, methylation of CpG9 and its nearby region was significantly enhanced in a sample from a <50 years old (data not shown). This result suggests that the down-regulation by DNA methylation occurs in a young population, and that the regulation mechanism involves malignancies such as juvenile breast cancer.
We also analyzed the tumor recurrence rate. ER is the one of the most valuable prognostic factors of breast cancer (16-20). Our analysis also reflected this tendency. The ER-positive group showed a significantly lower recurrence rate than the ER-negative group. However, in the ER-positive group, there were some patients with cancer recurrence. Even in ER-positive cases, the CpG9-methylated group showed no significant difference in recurrence rate compared to each of the ER-negative methylation groups. Previously, we reported the relationship between ER and CHIP protein expression (31), and recurrence-free survival was significantly better in the ER-positive cases with high CHIP expression than in ER-negative cases. However, although positive for ER, such cases with low CHIP expression tended to have worse recurrence-free survival than those with high CHIP expression. Additionally, there are several reports of expression regulated by pinpoint CpG methylation in breast cancer (32, 33). Our results and these reports suggest that this methylation of CpG9 can also predict the long-term prognosis of ER-positive breast cancer.
It is difficult to specify the prognosis of many patients by only one or a few factors such as protein expression. Additional factors that increase the accuracy of prognosis prediction in combination with existing factors should be identified. Epigenetic modifications such as DNA methylation or histone modification are powerful predictive factors and should be further investigated.
Acknowledgements
The Authors thank Toshifumi Niwa, Yuko Seino and Chika Tazawa for valuable assistance and comments on this study. The Authors also thank M. Arico from Edanz Group (www.edanzediting.com/ac) for editing a draft of this manuscript. This study was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science, Sports and Culture of Japan, a Grant-in-Aid for Cancer Research from the Ministry of Health, Labor, and Welfare, Japan, the Program for Promotion of Fundamental Studies in Health Sciences of the National Institute of Biomedical Innovation (NIBIO), and a Grant from the Smoking Research Foundation.
Footnotes
Authors’ Contributions
Tatsuyuki Gohno: Conceptualization, Methodology, Formal analysis, Investigation, Writing - original draft; Toru Hanamura: Methodology, Investigation, Writing - review and editing; Masafumi Kurosumi: Methodology, Investigation, Writing - review and editing; Hiroyuki Takei: Methodology, Investigation, Writing - review and editing; Yuri Yamaguchi: Methodology, Investigation, Writing - review and editing; Shin-Ichi Hayashi: Conceptualization, Methodology, Investigation, Writing - review and editing, Supervision, project administration, funding acquisition.
Conflicts of Interest
The Authors have no conflicts of interest to declare in relation to this study.
- Received November 25, 2021.
- Revision received December 13, 2021.
- Accepted December 14, 2021.
- Copyright © 2022 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.