


Abstract
Background/Aim: Stage-specific embryonic antigen (SSEA)-4 plays important roles in the malignant aggressiveness of various cancers. The aim of this study was to investigate the pathological characteristics of SSEA-4 in castration-resistant prostate cancer (CRPC). Materials and Methods: SSEA-4 expression and its pathological roles were evaluated in five prostate cancer (PC) cell lines and 27 CRPC tissues. The relationship between SSEA-4 and the androgen receptor (AR) was also examined. Results: SSEA-4 expression was detected in AR-negative cells (PC3, DU145, and AICaP1) but was not detected in AR-positive cells (LNCaP and AICaP2). SSEA-4 expression in human CRPC tissues was significantly higher than that in locally advanced or metastatic hormone sensitive PC (HSPC) tissues. A negative correlation was also detected between SSEA-4 and AR in CRPC tissues but not in HSPC tissues. Conclusion: SSEA-4 was over-expressed in CRPC and the changes were mediated by complex mechanisms that related to the AR and hormonal therapy.
- Stage-specific embryonic antigen (SSEA)-4
- castration-resistant prostate cancer (CRPC)
- androgen receptor (AR)
- hormonal therapy
Prostate cancer (PC) is one of the most common cancers in men. Hormonal therapy using androgen deprivation is one of the standard therapies that is often selected as a conservative therapy for patients with metastatic PC, advanced age, and/or severe comorbidity. However, in most cases, there are limitations to suppressing the malignant potential owing to the acquisition of resistance against hormonal therapy. Ultimately, due to the lack of androgen dependency during androgen deprivation, hormone-sensitive PC (HSPC) cells gradually transform into castration-resistant prostate cancer (CRPC) cells (1).
The prognosis of patients with CRPC has improved because of various newly developed treatment strategies (2, 3). However, research for novel therapeutic targets continues to further prolong the survival periods as the anti-cancer effects of current treatments are unsatisfactory. Furthermore, it is known that understanding the molecular mechanisms of androgen-independent tumor growth is essential for formulating new treatment strategies for patients with CRPC. In addition, there is a consensus that hyper-activated androgen-receptor (AR) pathways through up-regulated expression/gene mutation, presence of variants, and/or paracrine/autocrine androgen synthesis are important determinants of the pathological aggressiveness of CRPC (4, 5). Therefore, most prior studies on the acquisition of androgen independence have focused on AR. However, in addition to these direct pathological phenomena via AR pathways, several molecular signaling pathways have been reported to play important roles in the malignant behavior of CRPC via androgen-/AR-independent pathways (6, 7). Based on such information, it is concluded that understanding the interactions between AR and other cancer-related molecules is vital for improving the prognosis of patients with CRPC (8, 9).
Gangliosides, which are glycolipids containing sialic acid, play crucial roles in various cancer-related characteristics such as cell adhesion, motility, and invasion, and are used to determine prognosis and outcomes (10-12). Among them, monosialosyl globopentaosylceramide [MSGb5, also called stage-specific embryogenic antigen-4 (SSEA-4)] has been reported to be associated with carcinogenesis, malignant behaviors, therapeutic resistance, and survival in various malignant cells (13-16). In a previous study, we reported that SSEA-4 expression was positively associated with malignant aggressiveness, poor anti-cancer effects of neoadjuvant hormonal therapy (NHT), and worse outcome in patients with hormone-naïve PC (17, 18). Additionally, we found that SSEA-4 expression in patients post-NHT was significantly higher than that in patients pre-NHT (18). Thus, SSEA-4 expression may be up-regulated by hormonal therapy in patients with PC.
When this possibility is considered, the following important facts should be noted: 1) in addition to androgen-dependent signaling pathways, cancer stem cells (CSCs) play important roles in the malignant aggressiveness and biological characteristics of CRPC cells (19, 20); 2) in both HSPC and CRPC, CSCs are associated with resistance to anti-cancer therapies, including hormonal therapy (21, 22); 3) several CSC markers in PC cells are increased after androgen deprivation (23, 24); and 4) SSEA-4 has been recognized as a marker of CSCs in PC (25, 26). Consequently, detailed information on the pathological significance and regulatory mechanisms of SSEA-4 expression is crucial for devising novel treatment strategies for patients with PC, especially those with CRPC. However, the pathological significance of SSEA-4 in CRPC is still unclear as previous studies were performed for HSPC, and not for CRPC. Therefore, we performed in vivo and in vitro studies to clarify the biological and pathological characteristics of SSEA-4 in CRPC.
Materials and Methods
Antibodies and reagents. The mouse IgM monoclonal antibody (mAb) RM1 has been established as the specific mAb targeting SSEA-4 (27). We purchased the following antibodies: mouse anti-AR mAbs (Upstate Biotechnology, Lake Placid, NY, USA), horseradish peroxidase (HRP)-labeled rabbit anti-mouse IgM (Zymed Laboratories, South San Francisco, CA, USA), HRP-labeled rabbit anti-mouse IgG (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), and FITC-goat anti-mouse IgG+IgM (H+L) (Kirkegaard & Perry Laboratories; KPL, Middlesex, UK). We used N-Histofine® Simple Stain MAX PO (MULTI) (Nichirei Biosciences Inc., Tokyo, Japan) for immunohistochemical staining.
Cell culture. The prostate cancer cell lines PC3, DU145, and LNCaP, were purchased from American Type Culture Collection (ATCC) and they were cultured in RPMI-1640 medium containing 10% fetal bovine serum (FBS) (ICN Biomedicals, Aurora, OH, USA) at 37°C in a humidified atmosphere with 5% CO2. We established two androgen-independent cell lines (AICaP1 and AICaP2) by long-term culture of androgen-sensitive LNCaP cells in phenol red-free RPMI-1640 medium supplemented with 10% charcoal:dextran stripped (CDS)-FBS (Hyclone, Logan, UT, USA). This steroid-free medium was replaced every 3-4 days for 3 months according to a method described previously (28). Cell morphology was examined via phase contrast microscopy (Axiovert 25; Carl Zeiss, Jena, Germany).
Proliferation assay. LNCaP, AICaP1, and AICaP2 cells (1×103) were cultured in 96-well Corning tissue culture plates. The number of cells was determined daily with a plate reader, using the Fluoroskan Ascent Labsystem (Dainippon Pharma Co., Osaka, Japan) after the addition of calcein AM solution (C-3100MP, Invitrogen, Carlsbad, CA, USA).
Wound migration assay. LNCaP, AICaP1, and AICaP2 cells were incubated in 24-well Corning tissue culture plates for 24 h at 37°C. Subsequently, the surface of each culture was scratched with the tip of a 10 ml plastic pipette, washed to remove the debris, and treated with fresh culture medium. The distance between the borders (t0) was measured along the well using a grid and a Carl Zeiss phase-contrast inverted microscope (Axiovert 25) at 40× magnification. Experiments were performed in triplicate and the distance was measured in four fields of each well. After incubation for 24 h at 37°C, the distance between the borders (tfinal) was measured again in each field. The speed of migration was calculated as the distance by which the cells advanced from both borders (tg–tfinal) at the marked points divided by 2, and expressed as mm/h.
Matrigel invasion assay. LNCaP, AICaP1, and AICaP2 cells were grown in 75 cm2 culture dishes and trypsinized at 70-80% confluence. The cells were then counted and suspended in RPMI-1640 medium containing 0.1% CDS-FBS (2×105/ml). Thereafter, 100 ml aliquots of the cell suspensions were added to the upper compartments of a 24-well Matrigel-coated Transwell chamber (8 mm pores), while 600 ml of RPMI-1640 medium containing 0.1% CDS-FBS and 50 mg/ml of fibronectin was added to each lower compartment. After allowing the cells to migrate for 48 h, the cells remaining in the upper compartment were removed with a cotton swab. The cells that had invaded through the Matrigel and adhered to the outer (lower) surface of the membrane were fixed and stained with Giemsa stain for counting under a light microscope (×100) in five different fields (the average number per field was calculated). Each assay was performed in triplicate.
Western blot analysis. Cell lysates were obtained from cells grown to 70% confluence. Cells cultured in 75 cm2 culture dishes were washed thrice with PBS, scraped off, collected into Pyrex tubes, and centrifuged at 1,200 rpm (310×g) for 3 min. After the supernatant was discarded, 400 ml of lysis buffer [20 mM Tris (pH 7.4) 150 mM NaCl, 2 mM EDTA, 1% NP-40, 50 mM NaF, 10 mg/ml leupeptin, 1 mM Na3VO4, and 1 mM PMSF] were added to the cell pellet. The cells were then homogenized by sonication on ice, incubated for 30 min at 4°C, and centrifuged at 7,500 rpm (5,282×g) for 15 min at 4°C. Next, the supernatant was collected, and the protein concentration was assayed. Aliquots of lysates containing equal amounts of protein were boiled for 5 min in SDS-PAGE sample buffer [3× sample buffer], resolved by 10% SDS-PAGE, and transferred to polyvinylidene fluoride (PVDF) membranes (Hybond™-P; Amersham Biosciences, Piscataway, NJ, USA). After overnight blocking with 3% bovine serum albumin (BSA) in TBST at 4°C, the PVDF membranes were incubated overnight at 4°C with 1% BSA in TBST and the primary antibodies The membranes were then washed, incubated with the secondary antibodies (1:2,000), and developed via enhanced chemiluminescence using a WEST-one™ detection kit (iNtRON Biotechnology, Inc., Gyeonggi-do, Republic of Korea). The membranes were viewed using the LAS-1000 image analyzer (Fuji Film, Tokyo, Japan).
Analysis of gangliosides. Gangliosides were extracted according to a procedure described in previously published studies (29, 30). Briefly, samples of the same weight dissolved in chloroform/methanol (1:1) were spotted onto Silica gel 60 high-performance thin-layer chromatography (HPTLC) plates (Merck, Darmstadt, Germany) and developed in a chamber containing chloroform/methanol/0.2% aqueous CaCl2 (50:40:10). The expression of SSEA-4 was determined via HPTLC immunostaining with the RM1 mAb. Briefly, an HPTLC plate was immersed in high-molecular weight 0.5% poly(isobutyl methacrylate) (Aldrich Chemical Co., Milwaukee, WI, USA) in hexane/chloroform (9:1) for 40 s, dried in air for 30 min, blocked with 3% BSA in phosphate-buffered saline (PBS) for 1 h, and reacted with RM1 for 2 h at room temperature. Subsequently, the plate was washed, incubated with 0.1% HRP-labeled rabbit anti-mouse IgM in PBS, washed again, and stained with Konica Immunostaining HRP-1000 (Konica, Tokyo, Japan).
Androgen receptor cDNA transfection into AICaP1 cells. We detected the expression of SSEA-4 in AR-negative PC cells. To determine whether the AR directly influences the level of SSEA-4 expression, we transfected AICaP1 cells with a vector containing AR cDNA. The AR cDNA ligated into pIRES2-EGFR vector plasmid was kindly provided by Prof. Isaacs (Johns Hopkins University School of Medicine) and transfected into AICaP1 cells in 6-well plates using Lipofectamine 2000™ (Invitrogen) according to the manufacturer’s protocol (31). On the next day, 800 mg/ml of G-418 was added to the culture medium to select transfected cells called AICaP1-AR. AR expression by AICaP1-AR cells was confirmed using western blotting.
Patients with CRPC and NHT specimen data. The expression levels of SSEA-4 and AR were investigated in 27 patients diagnosed with CRPC. Additionally, we used the data on SSEA-4 expression in pre-NHT tissues (n=60) and post-NHT tissues (n=60) from a previously published report (18). The median (interquartile range) duration of NHT was 7 months (4-10 months), and detailed information on these tissues is available in our previous reports (18, 32). PC tissues judged as non-adenocarcinoma, such as neuroendocrine carcinoma or small cell carcinoma, were excluded from this study population. This study design complied with the principles of the Declaration of Helsinki and its revision and was approved by the Institutional Review Board of Nagasaki University Hospital (No: 16K15690). Additionally, written informed consent was obtained from all patients.
Immunohistochemistry. Immunohistochemical staining for SSEA-4 was carried out using formalin-fixed paraffin-embedded specimens of PC tissues. SSEA-4 expression level was classified into the following four groups according to the staining intensity and percentage of positively stained PC cells: none (–), mild (+), moderate (++), and strong (+++). The detailed methods used for immunohistochemistry and evaluation have been described in our previous reports (17, 18). AR expression was also immunohistochemically investigated in the same PC specimens. Anti-AR mouse monoclonal antibody (sc-7350) and EnVision Detection System (K5007) were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA) and Dako (Glostrup, Denmark), respectively. Immunohistochemical analyses were performed in accordance with a previously published report (33). Briefly, endogenous peroxidase activity was blocked by 3% hydrogen peroxide for 30 min, and antigen retrieval was performed in citrate buffer at 100°C for 30 min. The sections were then incubated overnight with the anti-AR antibody at 4°C. The immunoreactive score (IRS; values 0-12) was calculated as the product of staining intensity (none=0, weak=1, moderate=2, and strong=3) and proportion of positive cells (0-5.0%=0, 5.1-25%=1, 25.1-50.0%=2, 50.1-75%=3, and 75.1-100%=4). Slides were evaluated at different times by two investigators (YM and TM), who were blinded to the clinicopathological features. The mean of the values provided by these investigators was used as the IRS. In this study, AR expression was classified into 3 groups: low (IRS=0-5), middle (6-8), and high (9-12) based on its interquartile range.
Statistical analyses. The chi-square test was used for categorical comparison of the data. The Scheffé test was used for multiple comparisons of the data. All data are expressed as mean (SD), and significance was defined as p<0.050. All statistical analyses were performed on a personal computer with the statistical package, StatView for Windows (version 5.0; Abacus Concept Inc, Berkeley, CA, USA).
Results
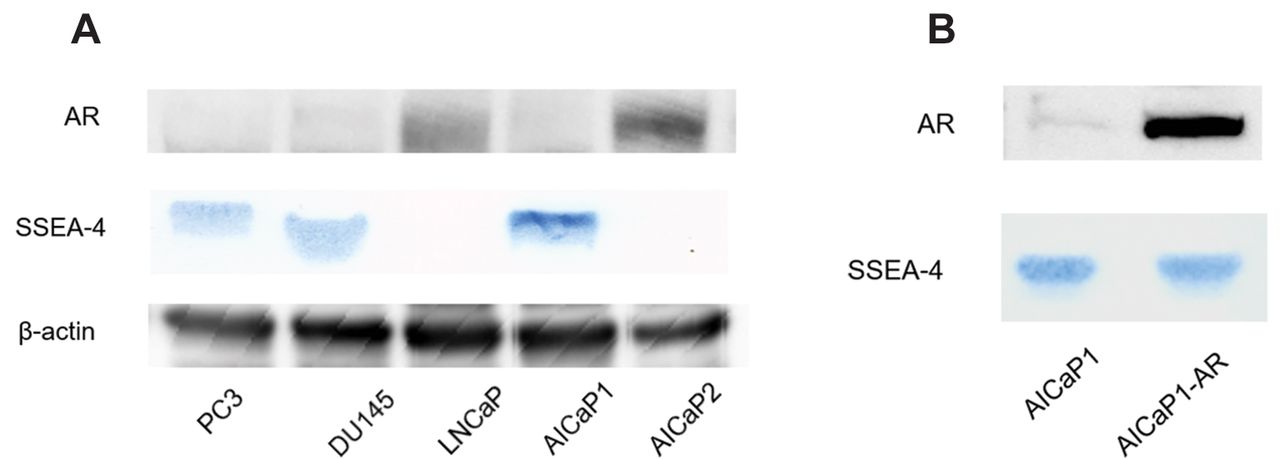
Expression levels of PSA, AR, and SSEA-4 in the prostate cancer cell lines. We examined the expression of AR in five PC cell lines (LNCaP, PC3, and DU145, AICaP1 and AICaP2). Neither AR nor PSA was detected in PC3, DU145, and AICaP1 cells, whereas both molecules were detected in AICaP2 and LNCaP cells (Figure 1A). Thus, the AR-related characteristics were found to differ between AICaP1 and AICaP2 cells. Subsequently, SSEA-4 was found to be expressed in PC3, DU145, and AICaP1 cells, but not in LNCaP and AICaP2 cells (Figure 1A). Altogether, SSEA-4 was detected in the AR-negative PC cell lines (PC3, DU145 and AICaP1), but not in the AR-positive cell lines (LNCaP and AICaP2).

SSEA-4 expression in prostate cancer cell lines. (A) Comparison of androgen receptor (AR) and stage-specific embryonic antigen-4 (SSEA-4) expression among prostate cancer cell lines. AR was analyzed via western blotting and SSEA-4 was examined via HPTLC immunostaining. (B) Change in SSEA-4 expression by transfection of AR cDNA into AICaP1.
Based on these findings, we transfected AR cDNA into AICaP1 cells to determine whether the AR had a direct influence on the expression of SSEA-4. After AR cDNA was transfected into AR-negative AICaP1 cells, AR expression by AICaP1-AR cells was confirmed. However, SSEA-4 expression remained unaltered in AICaP1-AR cells compared to that in AICaP1 cells (Figure 1B).
Pathobiological characteristics of AICaP1 and AICaP2 cells. We examined the pathobiological characteristics of two androgen-independent PC cell lines, that is, AR-negative AlCaP1 and AR-positive AlCaP2 cells used to identify the pathological characteristics of androgen-independent PC cells in previous reports (34), but their detailed biological characteristics and pathological roles remained unclear. Therefore, we evaluated them in various in vitro studies. Subsequently, the obtained growth curves revealed that the proliferation of AICaP1 cells was significantly higher than that of LNCaP and AICaP2 cells (p<0.05) (Figure 2A). Furthermore, the motility and invasive capacity of AICaP1 cells were significantly greater than those of LNCaP and AICaP2 cells (p<0.05 and p<0.01, respectively; Figure 2B and C). There were no significant differences in proliferation, motility, and invasive capacities between AICaP2 cells and LNCaP cells (p=0.40, 0.36, and 0.86, respectively).

Cell proliferation (A), motility (B), and invasion (C) assays carried out with LNCaP, AICaP1, and AICaP2 cells.
SSEA-4 expression in human CRPC tissues. Representative examples of mild (+), moderate (++), and strong (+++) expression of SSEA-4 in CRPC tissues are showed in Figure 3A-C, respectively. The staining distribution of SSEA-4 in CRPC tissues was similar to that found in HSPC tissues in previous studies (17, 18). Finally, 15 of the 27 (55.6%) CRPC tissues were assessed to have strong (+++) expression of SSEA-4, and no specimen was found to have no (-) expression (Figure 4). Upon comparison of SSEA-4 expression between CRPC and HSPC tissues, we observed that SSEA-4 immunoreactivity in CRPC was significantly higher than that in T3T4N0M0 and metastatic HSPC (p<0.001 and p=0.025, respectively; Figure 4). Likewise, regarding grade group (GG), SSEA-4 immunoreactivity in CRPC was significantly higher than that in the high-GG (Gleason score 4+4 or more) (p=0.006; Figure 4).

Representative examples of SSEA-4 expression in castration-resistant prostate cancer (CRPC) tissues. (A): mild (+), (B): moderate (++), and (C): strong (+++) expression. Magnification: ×400.

Comparisons of SSEA-4 expression levels between castration-resistant prostate cancer (CRPC) tissues and hormone sensitive prostate cancer (HSPC) tissues with various pathological features. SSEA-4 expression in CRPC tissues was significantly higher than that in metastatic HSPC tissues (A) and high-grade group tumors (B).
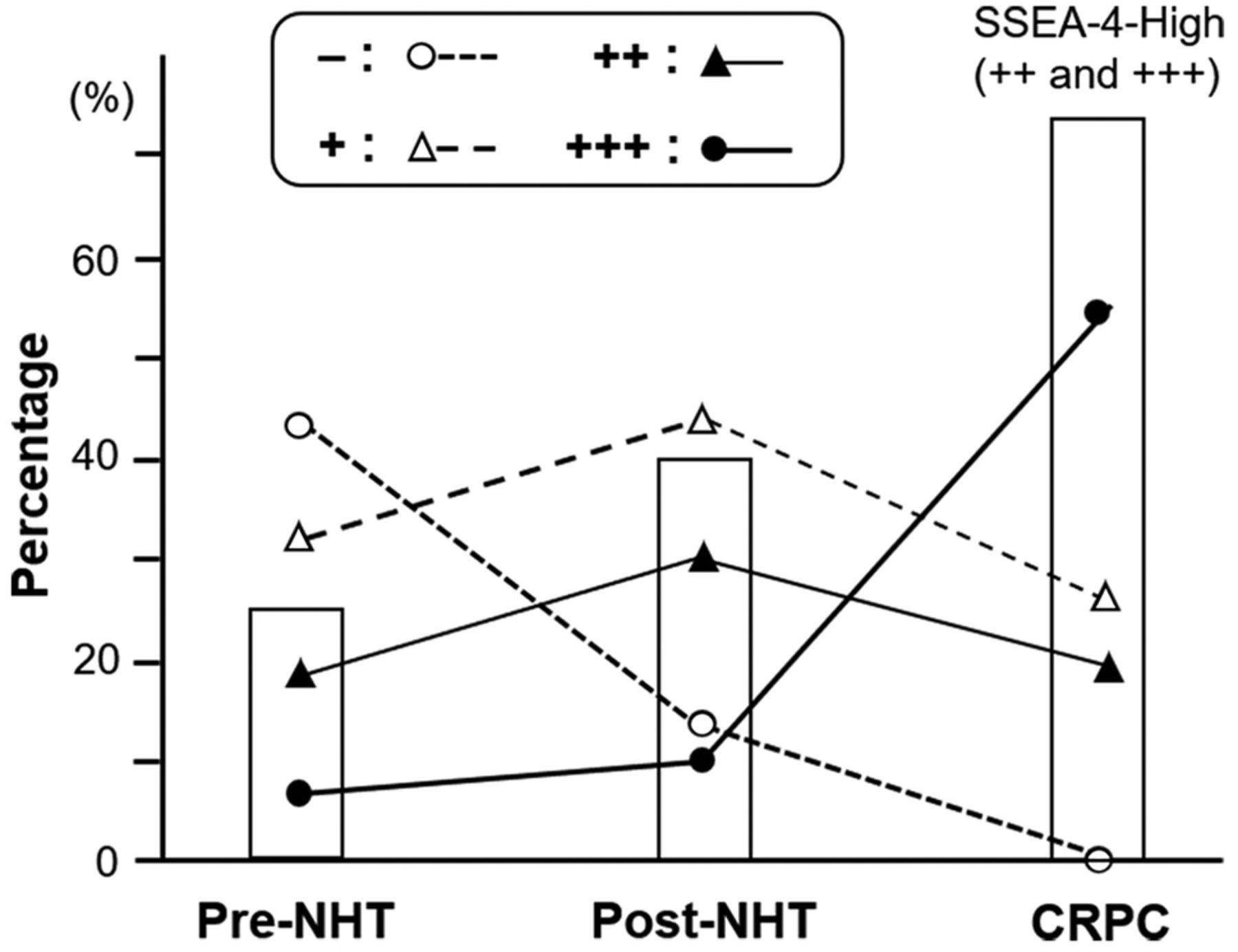
In a previous study, we reported the expression of SSEA-4 in pre-NHT and post-NHT tissues (18). In the present study, we compared SSEA-4 expression in these NHT (pre and post) and CRPC tissues (Figure 5). We found that the percentage of positively-stained PC cells were in the descending order of CRPC>post-NHT>pre-NHT tissues (bars in Figure 5). Furthermore, as shown via black dots in Figure 5, the number of strongly (+++) stained cells was remarkably higher in CRPC than in post-NHT tissues (by 44.6%), whereas the number in post-NHT tissues was only slightly higher than that in pre-NHT tissues (by 3.3%). In contrast, the number of SSEA-4-negative cells (blue dots in Figure 5) rapidly decreased after NHT, with no detection in CRPC tissues.

Comparisons of SSEA-4 expression levels among pre-neoadjuvant hormonal therapy (NHT), post-NHT, and CRPC tissues. Bars indicate the percentage of tissues with high SSEA-4 expression (++ and +++). Blue round, white triangle, black triangle, and red circle dot represent – (none), + (mild), ++ (moderate), and +++ (strong) expression of SSEA-4, respectively. Data on the ratios of pre-NHT and post-NHT tissues were acquired from our previous report (18).
A negative correlation between SSEA-4 and AR expression in PC tissues. The relationship between SSEA-4 expression and AR expression in CRPC tissues is shown in Table I. High IRS for AR was not found in CRPC tissues displaying strong SSEA-4 expression. Contrastingly, five of the seven CRPC tissues with mild SSEA-4 expression were determined to have a high IRS for AR. Statistical analysis revealed that SSEA-4 expression was negatively associated with AR expression in CRPC tissues (p=0.004). However, when a similar analysis was performed with HSPC tissues, no significant relationship was observed in organ-confined, T3-4N0M0, and metastatic PC (p=0.203, p=0.161, and p=0.349, respectively). Therefore, a negative relationship between SSEA-4 and AR was only detected in CRPC tissues.
Relationship between SSEA-4 and the androgen receptor in castration-resistant prostate cancer.
Discussion
In the present study, the in vitro studies revealed that SSEA-4 was detected in AR-negative PC cell lines (PC3, DU145, and AICaP1), but not in the AR-positive PC cell lines (LNCaP and AICaP2). Additionally, in human PC tissues, SSEA-4 immunoreactivity in CRPC cells was significantly higher than that in high GG (4/5), locally advanced, and metastatic HSPC. In a previous study, we reported that SSEA-4 expression was positively associated with GG, T stage, and metastasis in patients with HSPC (17). Therefore, SSEA-4 was hypothesized to have some crucial roles in CRPC, and its importance in patients with CRPC may be higher than those with HSPC. Nonetheless, our results showed that SSEA-4 was not expressed in AICaP2 even though this cell line was androgen-independent. Furthermore, the malignant potential, which was evaluated by cancer cell proliferation, migration, and invasion, was significantly higher in AICaP1 cells than in AICaP2 cells. Such findings suggest that SSEA-4 expression in PC cells is associated with complex mechanisms, including the AR signaling system.
Our results revealed an important aspect of the regulatory mechanism of SSEA-4 expression in PC cells. Among our five PC cell lines, SSEA-4 expression was detected in AR-negative PC cells, but not in the AR-positive PC cell lines. Moreover, our in vivo studies using human CRPC tissues demonstrated that SSEA-4 expression was negatively associated with AR expression. Such findings indicate that AR might play crucial regulatory roles in SSEA-4 expression. However, we should simultaneously note that such significant correlation between AR and SSEA-4 was not found in human HSPC tissues. Additionally, our in vitro study showed that the transfection of AR cDNA had no significant effect on SSEA-4 expression in AICaP1 cells. Based on these results, it is indicated that while there exists a negative correlation between AR and SSEA-4 in CRPC, AR status does not directly affect SSEA-4 expression. In fact, various cancer-related factors, including tumor necrosis factor-α, mammalian target of rapamycin, and Wnt signaling, have been reported to affect SSEA-4 expression under physiological and pathological conditions (5, 35, 36).
When the pathobiological significance of SSEA-4 in CRPC is being discussed, it must be noted that SSEA-4 is recognized as a marker of CSCs in a variety of malignancies (37, 38). Presently, acquisition of the CSC phenotype is regarded as an important characteristic of CRPC (21). Furthermore, androgen deprivation has been reported to stimulate the reprogramming of PC cells to acquire stem-cell like characteristics (39, 40). With regard to the change in SSEA-4 expression caused by hormonal therapy, our results demonstrated that its expression levels were the highest in CRPC, followed by post-NHT tissues, and pre-NHT tissues. In short, SSEA-4 expression was up-regulated by the duration of hormonal therapy. Based on these findings, we hypothesize that the up-regulation of SSEA-4 expression in CRPC reflects the increase in PC cells with stem cell-like characteristics. In fact, the AR-negative phenotype has been reported to be essential for maintaining the CSCs of PC and promoting CRPC cells (41), thereby supporting our abovementioned hypothesis that SSEA-4 expression is only detected in the AR-negative PC cell lines.
In recent years, SSEA-4 has been suggested to be a novel therapeutic target in a variety of malignancies (42-44). For example, a mAb directed against SSEA-4 (MC-813-70) has been suggested as a potential therapeutic agent for triple-negative breast cancer cells expressing SSEA-4 (43). Interestingly, our results showed that SSEA-4 was expressed in AR-negative CRPC. Therefore, it is possible that inhibition of SSEA-4 may lead to suppressed cell survival and progression, even though PC cells manage to survive via AR-independent signaling pathways. Based on these findings, we suggest that SSEA-4 is a potential therapeutic target in patients with PC, especially those with CRPC.
Our study has several limitations. First, the number of CRPC tissues was relatively low. Second, the detailed biological and molecular characteristics of our established androgen-independent PC cell lines (AICaP1 and AICaP2) have not been fully elucidated. Lastly, we could not elucidate the detailed interaction between SSEA-4 and AR in CRPC in the current study. Therefore, further studies elucidating the pathological and molecular characteristics of these cell lines are important for their future use.
Conclusion
Our in vitro studies demonstrated that SSEA-4 was expressed in AR-negative PC cell lines, whereas not expressed in AR-positive ones. In human CRPC tissues, a negative correlation between SSEA-4 and AR was observed. We hypothesize that SSEA-4 is indirectly associated with AR, although the detailed mechanisms of these interactions remain to be fully understood. Additionally, SSEA-4 was expressed in all cases of CRPC tissues and its immunoreactivity level was significantly higher in human CRPC tissues than in HSPC with high GG and metastasis in the descending order of CRPC>post-NHT>pre-NHT tissues. These results indicate that SSEA-4 is a novel biomarker for CRPC, especially AR-negative PC cells. According to previous studies on various malignancies, SSEA-4 might reflect the increased CSC count in CRPC tissues. We suggest that SSEA-4 is a potential therapeutic target in patients with CRPC.
Acknowledgements
This work was supported by a grant from JSPS KAKENHI (grant number: 18K09197 and 19K09713).
Footnotes
This article is freely accessible online.
Authors’ Contributions
Study concept: YMi and SS. Study design: YM. In vitro analyses: TT, TS, AI, SS. Clinical data collection: KM, TM, KO. Immunohistochemical analyses: JH, TM, YM, YMu. Statistical analyses: YM, TM. Manuscript preparation: JH, TM, YMi. Manuscript editing: AI, SS. Manuscript review: HS. All Authors read and approved the final manuscript.
Conflicts of Interest
The Authors declare no conflicts of interest in relation to this study.
- Received May 26, 2021.
- Revision received June 16, 2021.
- Accepted June 17, 2021.
- Copyright © 2021 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.