


Abstract
Background/Aim: Effect of capsicodendrin on the NF-κB pathway was studied in MCF-7 cancer cells. Materials and Methods: The transcription factor assay was used to screen for NF-κB activity. The effect on IKKβ, ICAM-1, and caspase-7 were studied using western blot. Caspase-1 was studied using Promega Caspase-Glo® assay. Reactive oxygen species (ROS) were detected using the fluorescent probe DCFH-DA. The potentiometric dye JC-1 was used to assess mitochondrial membrane potential (ΔΨm) and the cell cycle was examined using a fluorescence-activated cell sorter. Results: NF-κB p65 inhibitory effect was IC50=8.6 μM and cytotoxic activity was IC50=7.5 μM. The upstream IKK and the downstream ICAM-1 were down-regulated. Sub G1-phase population increased to 81% after 12 h of treatment with capsicodendrin (10 μM) and there was no loss of ΔΨM. Conclusion: Increased levels of intracellular ROS promoted activity of caspase-1 and induced cell death in MCF-7 cells. Capsicodendrin may be a future anticancer agent that prevents the progression of metastatic breast cancer.
Despite new effective anticancer treatments, chemoresistant breast tumor cells spread to other organs and the survival rate (26%) remains low for metastatic breast cancer (1-3). Breast cancer patients have high serum levels of pro-inflammatory cytokine tumor necrosis factor-α (TNF-α). It has also been shown that transmembrane TNF-α plays a role in the development of chemoresistance and metastasis of breast cancer cells (4-5). TNF-α induces the activity on the nuclear factor kappa B (NF-κB) (Figure 1), which is a heterodimer composed of p65 bound to IκB. IκB kinase (IKK) phosphorylates IκB. The release from IκB activates NF-κB before it is translocated to the nucleus, and the blockage of both IKKβ and nuclear import reduces tumor expansion (6-8). Activation of the NF-κB p65 signal plays an important role in vivo in tumor progression and promotes the resistance to certain tumorigenic agents and chemotherapy (7, 8). Previous studies have shown that NF-κB p65 prevents cancer cells from entering apoptosis and contributes to chemoresistance as well as to the progression of the disease (9). In addition, it has been shown that MCF-7 estrogen-sensitive breast cancer cells overexpress the anti-apoptotic protein Bcl-2, which prevents apoptosis. Thus, inhibition of NF-κB p65 prevents tumor growth and metastasis of estrogen-sensitive breast cancer (10, 11).

Capsicodendrin role in the NF-κB p65 pathway. (A) Structure of capsicodendrin isolated from Cinnamosma fragrans and (B) proposed pathway for the transcription factor NF-κB p65 in MCF-7 estrogen-dependent cells.
Moreover, intracellular adhesion molecule ICAM-1 has been shown to be induced by TNF-α in MCF-7 cells (Figure 1). Herein, the effects on ICAM-1 and the anti-proliferative effect of capsicodendrin in MCF-7 estrogen-dependent breast cancer cells were investigated. Also, the effect of capsicodendrin on the NF-κB pathway and the viability of treated cells was further analyzed. The apoptotic effect and the effect on cellular oxidative stress was also investigated in vitro. The aim of this study was to assess the possible mechanism of action through which capsicodendrin (Figure 1A) induces cell-cycle arrest in breast cancer cells.
Materials and Methods
Cell culture. The MCF-7 cancer cell line was obtained from American Type Culture Collection (ATCC# HTB-22, Manassas, VA, USA). Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) and Roswell Park Memorial Institute medium (RPMI-1640), containing 10% fetal bovine calf serum and 10% Antibiotic-Antimycotic (Gibco, Rockville, MD, USA). Cells were kept at 37°C and in an atmosphere containing 5% CO2.
SRB assay. MCF-7 cells were plated in a 96-well plate and treated with capsicodendrin for 72 h at 37°C and incubated in a 5% CO2 atmosphere. Cells were fixed using 20% trichloroacetic acid for 30 min, followed by staining with sulforhodamine (SRB) (0.4%) for 30 min at room temperature. SRB was removed by washing with acetic acid three times. After adding 200 μM Tris base solution to each well, the 96-well plates were placed on a shaker for 5 min. Absorbance reading was performed at a wavelength of 515 nm using a Fluostar Optima plate reader (BMG Labtech Inc, Durham, NC, USA). Paclitaxel (Tocris Bioscience, Bristol, UK) was used as a positive control.
NF-κB assay. The NF-κB p65 assay was performed according to a previously established protocol (12). Nuclear extracts were prepared from HeLa cells and the Transcription Assay System (Pierce Biotechnology, Rockford, IL, USA) was used to evaluate the binding affinity of the NF-κB subunit p65 to the biotinylated consensus sequence. Luminescence was detected using a Fluostar Optima plate reader (BMG Labtech Inc). Rocaglamide (Enzo Life Sciences, Inc., Farmingdale, NY, USA) was used as a positive control.
Immunoblotting. Cells were treated with capsicodendrin at different concentrations (0.008, 0.016, 0.4, 2.0 and 10 μM) for 3 h. Briefly, cells were lysed using PhosphoSafe Lysis Buffer (Novagen, Madison, WI, USA). Protein concentration was determined by using a Bradford protein assay kit and albumin standard (Thermo Scientific, Waltham, MA, USA). Absorbance was measured using a Fluostar Optima plate reader (BMG Labtech Inc.). Lysates were analyzed by western blot analysis with primary (1:1,000) and secondary antibodies (1:2,000). Equal amounts of protein (20 μg) were loaded together with a LDS sample loading buffer (Invitrogen, Carlsbad, CA, USA) and resolved using Nu-PAGE 10% SDS-PAGE Bis-Tris gels together with SeeBlue® Plus2 Pre-Stained Standard (Invitrogen). Proteins were transferred to a polyvinylidene fluoride (PVDF) membrane using transfer buffer, tris-buffered saline solution with Tween 20® (TBS-T). The blots were then blocked at room temperature using non-fat milk and probed using primary antibodies against each target protein including NF-κB p65, IKKβ, ICAM-1, and caspase-7, using BSA in TBS-T overnight. Conjugated antibodies were detected using chemiluminescent substrates with a Supersignal Femto kit from Thermo Scientific.
Reactive oxygen species. A reactive oxygen species (ROS) assay was performed following a previously described procedure (13). Generated intracellular levels of ROS were measured using the fluorescent probe 2’,7’-dichlorodihydrofluorescein diacetate (DCFH-DA). MCF-7 cells were seeded in a 96-well plate, treated with capsicodendrin, daunomycin, vitamin C or DMSO and followed by 5 h incubation at 37°C with 5% CO2. Subsequently, cells were incubated with H2O2 (1.25 mM) and FeSO4 for 30 min at 37°C. Later, the fluorescent probe DCFH-DA was added to determine intracellular ROS. Fluorescence was measured using FLUOstar Optima fluorescence plate reader (BMG Lab technologies GmbH Inc.) at an excitation wavelength of 485 nm and emission wavelength of 530 nm. All treatments performed in triplicate are representative of at least two different experiments.
Cell cycle. Cells were plated and treated using five different concentrations of capsicodendrin. After 12 h of incubation, cells were harvested and pelleted by centrifugation, washed with PBS and fixed in ice-cold 70% ethanol. The DNA was stained with 10 μg/ml propidium iodine (PI) in a reaction solution containing 1 mM EDTA and 100 μg/ml RNase A. Fluorescence emitted from the propidium iodine-DNA complex was quantified using BD FACS Canto II (Biosciences, San Jose, CA, USA) at 488 nm.
Mitochondrial transmembrane potential (ΔΨm) assay. The mitochondrial transmembrane potential (MTP) fluorescence-activated cell sorting (FACS) assay kit (Cayman Chemical Company, Ann Arbor, MI, USA) was used to assess MTP (ΔΨm) in MCF-7 cells after treatment, using FACS analysis (12). Cells were seeded on 10 cm plates and treated with capsicodendrin for 24 h. Next, cells were harvested using Trypsin-EDTA (Gibco), washed in phosphate-buffered saline (PBS) and re-suspended in assay buffer. The potentiometric dye 5,5’,6,6’-tetrachloro-1,1’,3,3’-tetraethylbensimidazoylcarbocyanine iodide (JC-1) was used to stain MCF-7 cells. A volume of 50 μl of JC-1 stain was added to the cells and incubated for 15 min at 37°C in 5% CO2. At high ΨΔm, red fluorescence J-aggregates are formed in healthy cells; however, in apoptotic cells with low ΔΨm, JC-1 remains in the monomeric form, which only exhibits green fluorescence. Analysis was performed using FACS Canto II (BD Bioscience, San Jose, CA, USA). The mitochondrial function was assessed, and the J-aggregates were detected using an excitation wavelength of 520-570 nm, and emission of 570-610 nm, respectively.
Caspase-Glo 1 inflammasome assay. Caspase-1 activity was determined using a modified protocol from Promega Caspase-Glo® 1 Inflammasome Assay (G9951) (Promega, Madison, WI, USA). MCF-7 cells were plated overnight in white 96-well plates in 100 μl medium at a density of 20×104 cells/ml, and at 37°C in a humidified 5% CO2 incubator. Next, cells were treated with capsicodendrin or doxorubicin (control), for 24 h. Four concentrations of both capsicodendrin and the control were tested, using the IC50 value as the second concentration and one concentration 5× higher than IC50 as well as two concentrations 5× and 10× lower than the IC50 value. Reagents used in the assay were prepared according to the specifications detailed by the manufacturer. After treatment, 70 μl of medium was then discarded and the appropriate volume of Caspase-Glo® 1 Reagent was added, while Caspase-Glo® 1 YVAD-CHO Reagent was added at a ratio of 1:1 sample volume to reagent volume. The plate was then incubated at room temperature for 1 h and the luminescence was measured up to 120 min using a Fluostar Optima plate reader (BMG Labtech Inc.).
Results and Discussion
Capsicodendrin was isolated from the bark of Cinnamosma fragrans Baill. (Canellaceae) (Figure 1A) (14). C. fragrans is a medicinal plant endemic to Madagascar and is used in the treatment of malaria symptoms, fatigue, intestinal parasite infections, intoxication and headaches (14, 15).
The results obtained from the SRB assay showed that capsicodendrin displayed cytotoxic activity in the MCF-7 hormone-dependent breast cancer cell line (IC50=7.5 μM), suggesting the potential for further analysis as breast cancer is a leading cause of death in women. The cytotoxic activity was compared to the inhibitory activity of paclitaxel (IC50=0.0021 μM). Capsicodendrin has previously displayed cytotoxic activity against other cancer cell lines: HeLa cervical cells (IC50=2.97 μM), HT-29 colon cancer cells (IC50=1.04 μM) and murine leukemia cell line L1210/0 (IC50=0.58 μM), human T-lymphocyte cell lines Molt4/C8 (IC50=1.51 μM) and CEM/0 (IC50=1.61 μM) (14, 16, 17).
Capsicodendrin also displayed NF-κB inhibitory activity (IC50=8.6 μM) when tested by using the NF-κB p65 ELISA assay (Figure 1B). The NF-κB p65 inhibitory effect in HeLa cells was compared to that of rocaglamide (IC50=0.075 μM). Subsequently, western blot analysis was performed at five different concentrations to assess the expression levels of NF-κB p65 in hormone-dependent breast cancer MCF-7 cells. At the concentration of 0.04 μM, the levels of NF-κB p65 were down-regulated in capsicodendrin-treated MCF-7 cells when compared to untreated cells (Figure 2A).

Immunoblotting studies on NF-κB. (A) Western blot analysis shows the expression of the transcription factor NF-κB after 3 h of incubation with capsicodendrin. The NF-κB subunit p65 (Rel A) was down-regulated in capsicodendrin-treated MCF-7 cells and the effect on protein expression was concentration-dependent. The positive control was treated with rocaglamide and the negative control was not treated. β-Actin was used as an internal control. (B) Western blot analysis showed the expression of proteins in the NF-κB pathway. The expression of IκB kinase β (IKKβ) was down-regulated in capsicodendrin-treated MCF-7 cells in a concentration-dependent manner. β-Actin was used as an internal control.
Furthermore, IKK is a pivotal upstream regulator of the NF-κB pathway (18). Thus, the effect on the upstream IKKβ was assessed (Figure 2B). Capsicodendrin decreased the expression of IKKβ as shown by western blot analysis at the treated concentration of 0.04 μM. It has previously been shown that NF-κB acts as a pro-metastatic factor, facilitating adhesion of tumor cells to endothelial cells and has also been reported that capsicodendrin prevented the formation of new blood vessels (19). In the present study, it was shown that the expression of intracellular adhesion molecule (ICAM-1) was down-regulated in MCF-7 cells that were treated with capsicodendrin at 0.016 μM (Figure 3A). The glucoprotein ICAM-1 is a member of the immunoglobulin superfamily. It is an essential molecule in cell-to-cell adhesion and mediates interaction between different cells. Silencing the expression of ICAM -1 decreases colony-formation in MCF-7 tumor cells and reduces cell adhesion. Further, it is involved in the transmigration of tumor cells and during metastasis. The down-regulation of human ICAM-1 inhibits the invasion and metastatic ability of human breast cancer cell lines (20). A previous study has also shown that NF-κB p65 inhibition suppressed expression of ICAM-1 and sensitized cells to anticancer compound treatment (21, 22). Additionally, the present findings are in complete agreement with our previous studies performed in order to understand the mechanism of action of capsicodendrin, and demonstrated that it displayed angiostatic activity through selective inhibition of VEGFR2-mediated AKT signaling and dysregulated autophagy in endothelial cells during angiogenesis (21). In addition, it has previously been shown that ICAM-1 is under the control of NF-κB in MCF-7 cells during the metastatic phase when tumor cells migrate through the vessels in the lymphatic route (22). Thus, ICAM-1 is a key player in cancer and the effect of capsicodendrin on this target is worthy of further study.

Expression of ICAM-1 and caspase-7. (A) ICAM-1 expression in capsicodendrin-treated MCF-7 cells after 3 h of treatment. Decreased levels of adhesion molecule ICAM-1 were detected in TNF-α treated cells. The effect was concentration-dependent. (B) The effect of capsicodendrin on caspase-7 expression after 3 h of treatment in MCF-7 cells breast cancer cells was analyzed. Immunoblot analysis showed that increasing concentrations of caspsicodendrin led to decreased levels of caspase-7. The effect was concentration-dependent. The effect was compared with rocaglamide, the positive control.
Moreover, treatment with capsicodendrin induced oxidative stress in treated estrogen–sensitive MCF-7 cells. The levels of ROS increased in capsicodendrin-treated cells compared to cells treated with the positive control daunomycin (Figure 4). Similarly, it has been found that capsicodendrin depleted glutathione in human myeloid leukemia cells and reduced the intracellular antioxidant capacity of cancer cells (23). This suggests that oxidative stress promoted cell death in treated MCF-7 cells. Studies have shown that ROS play an important role in multidrug resistance in MCF-7 cells and thus together with the inhibition of NF-κB p65, the mechanism of action of capsicodendrin may prevent chemoresistance in cancer patients (24).
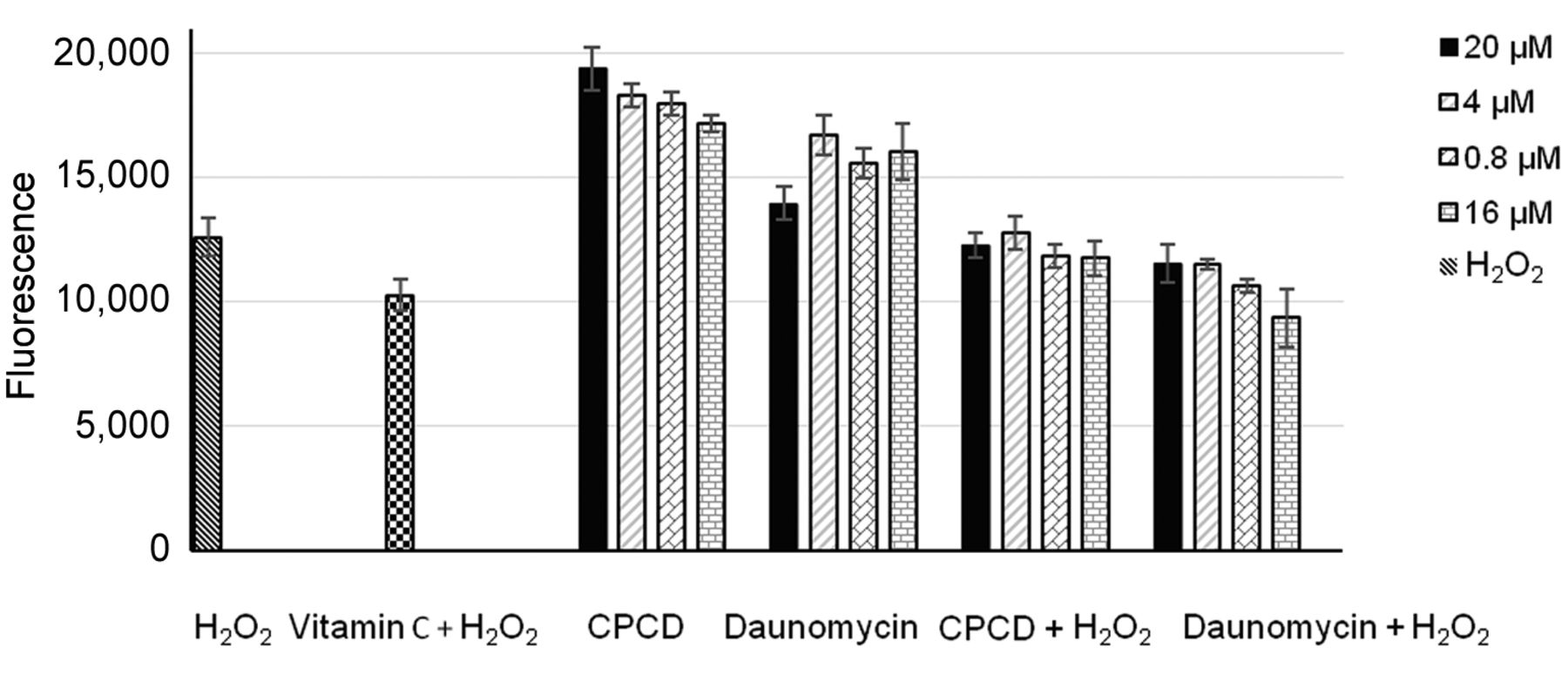
Intracellular levels of reactive oxygen species (ROS) formed in capsicodendrin treated estrogen-dependent breast cancer MCF-7 cells after 5 h of treatment. The effect was compared to the positive control daunomycin (a topoisomerase inhibitor). The cells were treated at four different concentrations. The formation of ROS was detected with the fluorescent probe DCFH-DA. The highest concentration of capsicodendrin (20 μM) led to a higher ROS formation than the highest concentration of daunomycin (20 μM). The effect of capsicodendrin was concentration-dependent. Vitamin C was used as a negative control.
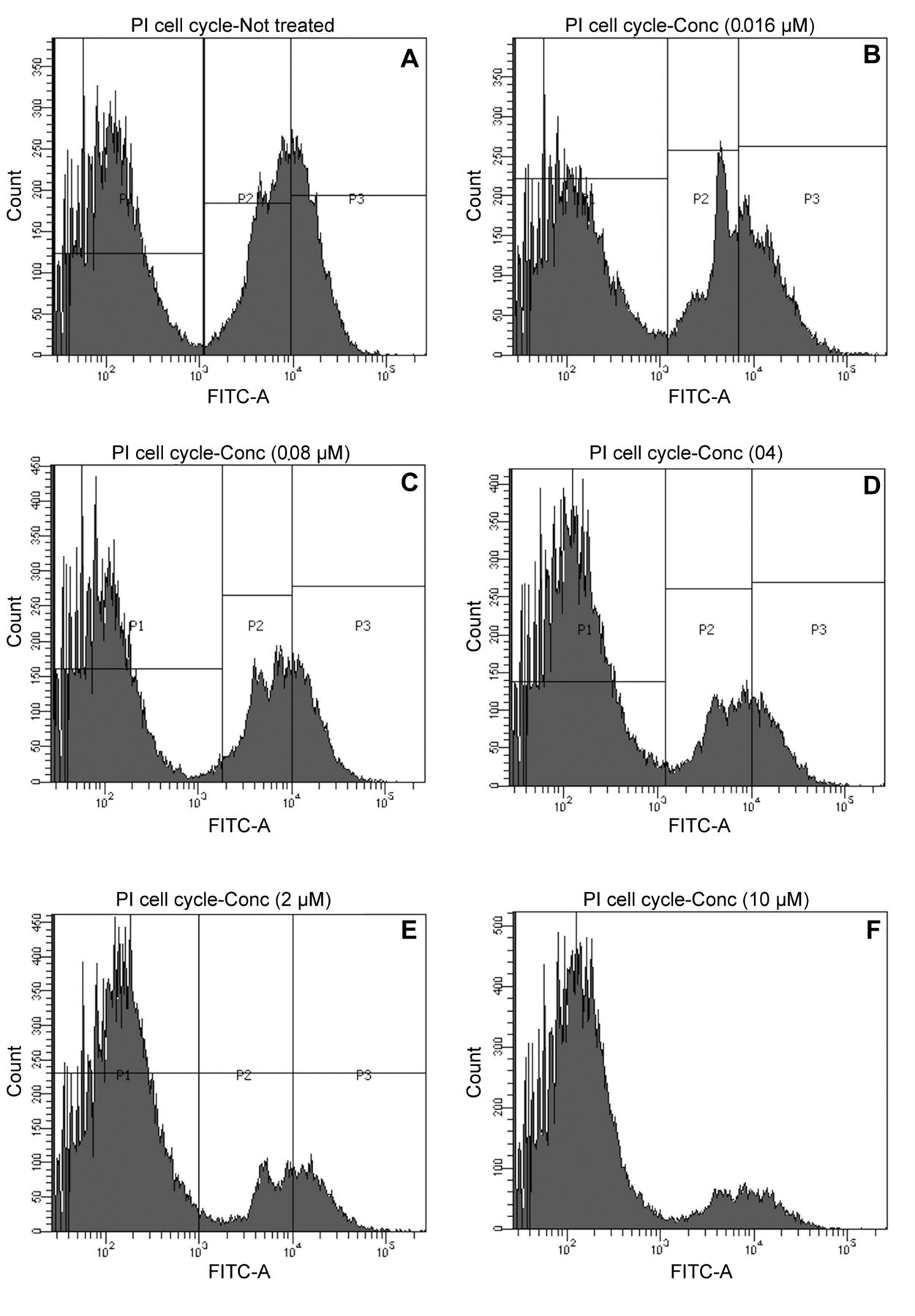
The effect of capsicodendrin on cell-cycle arrest was examined using FACS. The sub G1-phase population increased to 81% after 12 h of treatment with capsicodendrin (10 μM). This was compared to 46% of cells detected in sub G1-phase in non-treated MCF-7 cells (Figure 5A-F). There was a significant loss of cells and the results showed that cell cycle arrest had been induced. The effect observed was concentration-dependent; however, mitochondrial induced cell-death was not observed in capsicodendrin-treated cells and JC-1 monomers were not detected (Figure 6). The results showed no loss of the outer ∆Ψm nor mitochondrial dysfunction or cell-death in capsicodendrin-treated MCF-7 cells. Hence, the effect was found to be independent of mitochondrial activity.
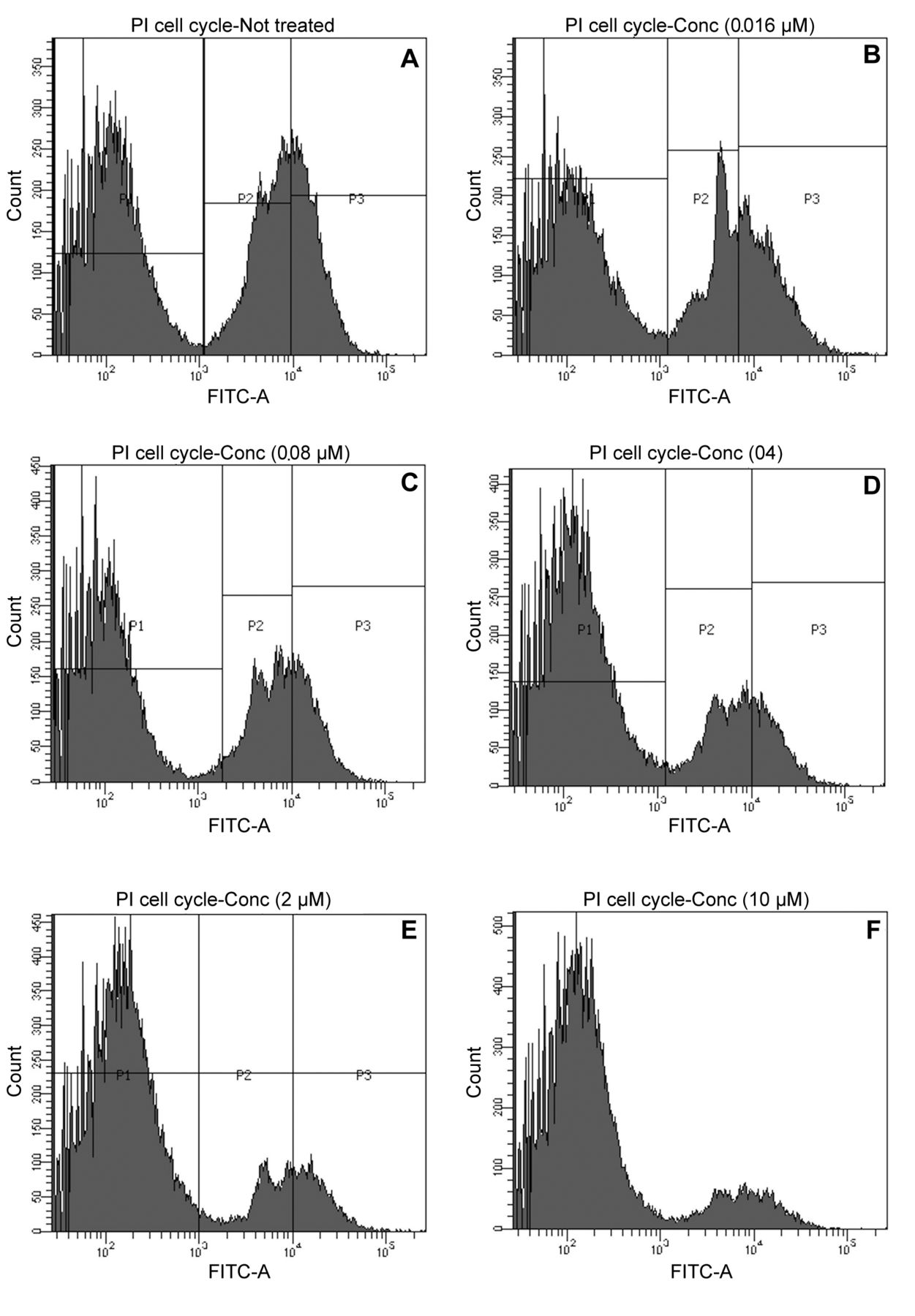
The effect of capsicodendrin on DNA fragmentation was evaluated in MCF-7 cells using fluorescence-activating cell flow cytometry (FACS). The cells were treated with capsicodendrin at five different concentration levels for 12 h: (A) control, (B) 0.0016 μM, (C) 0.008 μM, (D) 0.4 μM, (E) 2.0 μM, (F) 10 μM. The increasing concentration of capsicodendrin led to an increase in the number of cells in sub-G1 phase.
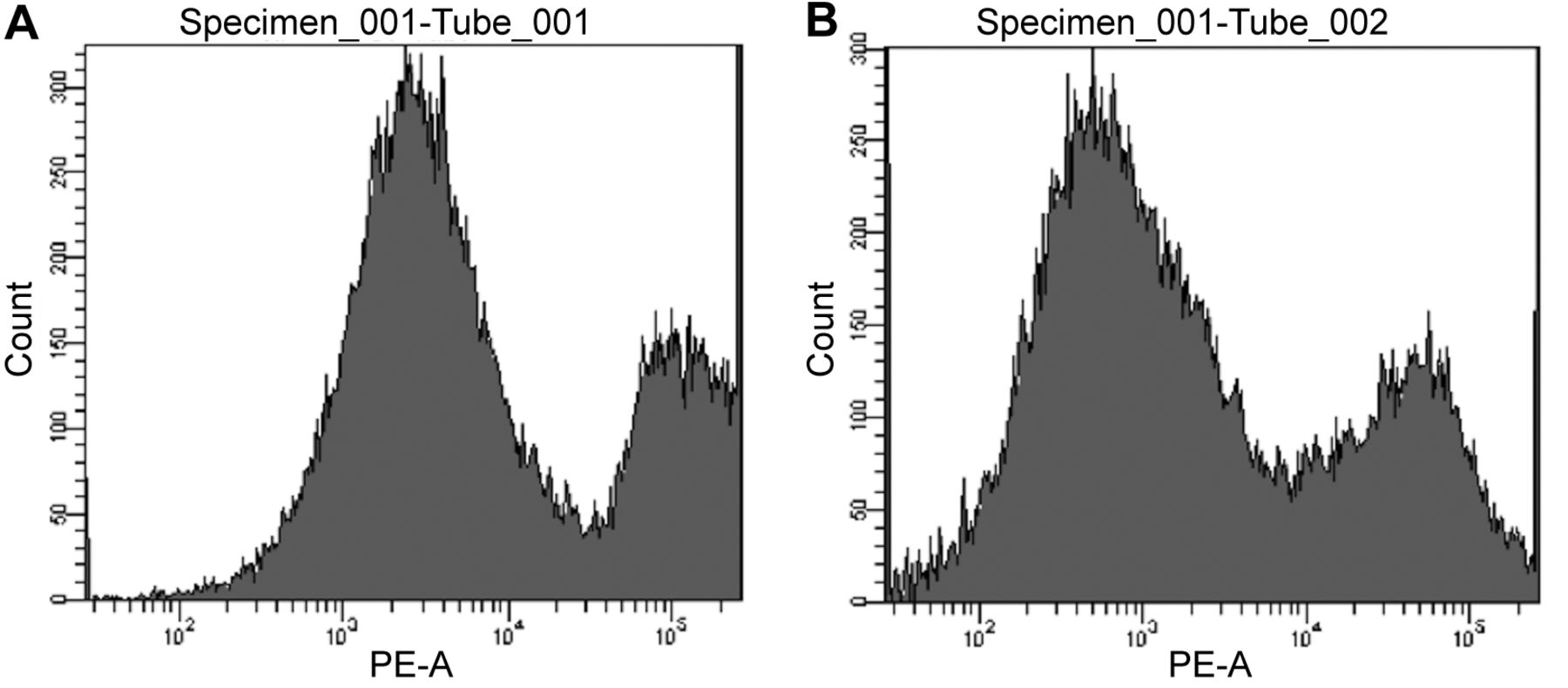
The loss of mitochondrial membrane potential (∆Ψm) is a major event during apoptosis. ∆Ψm was assessed using the cationic dye JC-1 in MCF-7 cells treated without (A) or with capsicodendrin (B). In apoptotic unhealthy cells, JC-1 monomers were detected, as indicated by the green fluorescent cells when analyzed by cell flow cytometry. There was no increase in the formation of JC-1 monomers (564-606 nm) in capsicodendrin-treated cells when compared with untreated cells.
According to previous studies, MCF-7 cells have a high expression of the anti-apoptotic protein Bcl-2 and lack the cellular response to apoptotic stimuli such as e.g., the activity of caspase-3 (25, 26). To further understand the mechanism and the biochemical interactions through which capsicodendrin promoted cell-cycle arrest in MCF-7 cells, the effect on both caspase-1 and caspase -7 were investigated.
Caspase-1 is part of the inflammasome and is involved in inflammatory response. It has been shown to activate pro-inflammatory cytokine IL-1β (27). Increasing evidence suggests that IL-1β might be involved in autoinflammatory diseases of unknown origin (28). Recent studies have shown that there are increased levels of IL-1β in the tumor microenvironment of breast cancer tissue. An increase in IL-1β was estrogen-dependent and IL-antagonists inhibited tumor development and angiogenesis (29-31). Herein, we report the effect of capsicodendrin on caspase-1 enzymatic activity in MCF-7 cells (Figure 7A). Capsicodendrin (0.12-3.50 μM) increased caspase-1 activity in a caspase-1 GLO assay compared with doxorubicin (0.14-3.5 μM) (Figure 7B). In addition, treatment with increasing concentrations of caspsicodendrin (0.12-3.0 μM) induced caspase-1 activity even in cells treated with the inhibitor YVAD-CHO. The effect correlates with increased levels of ROS and suggests that ROS promotes the activity of caspase-1. This effect was both concentration-dependent and time-dependent. It has previously been reported that caspase-1 induces p53-dependent cell-cycle arrest and inflammasome-mediated apoptosis in estrogen dependent MCF-7 cells (32).

Caspase-1 activity was determined using a modified protocol from Promega Caspase-Glo® 1 Inflammasome Assay (G9951). The YVAD-CHO is a specific caspase-1 inhibitor. The MCF-7 cells were treated with doxorubicin (topoisomerase inhibitor) and capsicodendrin. Capsicodendrin (0.12-3.50 μM) (A) increased caspase-1 activity compared with doxorubicin (0.14-3.5 μM) that showed inhibitory activity (B). Increased concentration levels of caspsicodendrin (0.12-3.0 μM) induced caspase-1 activity. Capsicodendrin potentiated caspase-1 activity in a concentration-dependent manner. The effect was compared to the negative control.
Earlier studies have shown that the levels of caspase-7 are over-expressed in estrogen-dependent breast cancer cells and thus the levels were evaluated in capsicodendrin wild-type p53 protein expressing MCF-7 cells (33, 34). The results of this study showed that the levels of caspase-7 were down-regulated in capsicodendrin-treated (0.008-10 μM) MCF-7 cells (Figure 3B). The CASP7 promoter has five estrogen responsive elements and thus could represent potential specific targets in estrogen-dependent cancer cells (33). Furthermore, caspase-7 targets various cell-cycle inhibitors such as p21 and DNA repair proteins (35, 36). It has been shown that reduction in p21 contributes to proliferation and cell growth. Cyclin-dependent kinases (CDKs) play an important role and promote cell cycle progression (37). Thus, CDK inhibitory proteins, such as p21, inhibit CDK activity, which, in turn, causes cell-cycle arrest in G1/S phase and negatively affects cancer cell proliferation (38, 39). The down-regulated levels of caspase-7 correlated with the cell cycle arrest in MCF-7 treated cells. This inhibitory effect may have depended on the inhibitory effect of p21 on cell proliferation (33).
Furthermore, in our previous studies, capsicodendrin was shown to be able to be converted to its more reactive monomer cinnamodial, in the presence of nucleophilic solvents such as pyridine and DMSO in vitro. We proposed that capsicodendrin is present in plants as a chemical reservoir of the bioactive cinnamodial against predators (20). The conversion of capsicodendrin to cinnamodial may qualify the compound as a prodrug; however, this remains to be evaluated in vivo. In this study, we focused on understanding the antiproliferative mechanisms of capsicodendrin against MCF-7 cells. Diligent efforts were made to use as little DMSO as possible during the assays in order to specifically examine the effect of capsicodendrin. In summary, the results from this study showed that capsicodendrin inhibited the NF-kB pathway, increased oxidative stress, induced cell-death through caspase-1, and down-regulated the expression of caspase-7. Targeting the NF-κB pathway could potentiate the effect of existing anticancer treatments in the management of metastatic cancer (33, 40, 41).
Conclusion
The findings from the present drug discovery study showed that the NF-κB inhibitory effect of capsicodendrin induced cell death through a mitochondrial-independent pathway in malignant MCF-7 cells. The NF-κB inhibitory effect of capsicodendrin increased oxidative stress, the activity of caspase-1, and decreased the expression of estrogen-dependent caspase-7. In conclusion, inhibitors that target the NF-κB p65 signal pathway may induce additive anti-cancer therapeutic effects. Capsicodendrin and its potential analogues may be novel anticancer agents targeting metastatic breast cancer cells that may improve the prognosis of recurrent breast cancers.
Acknowledgements
The Authors greatly acknowledge the financial support through program project grant P01 CA125066 and its supplement P01 CA125066-10S1 from the National Cancer Institute, NIH, Bethesda, MD to carry out the presented work. The Authors are also thankful for the partial support provided by the Jack L. Beal Endowment Fund of Dr. A. Douglas Kinghorn for Mr. Nathan Ezzone.
Footnotes
↵This article is freely accessible online.
# Current address: Department of Chemistry and Biomedical Sciences, Faculty of Health and Life Sciences, Linnaeus University, Kalmar, Sweden.
Authors’ Contributions
Dr. Esperanza J. Carcache de Blanco coordinated and supervised overall efforts and conceived experiments in vitro as well as participated in the preparation and revision of the manuscript. Dr. L. Harinantenaina Rakotondraibe provided capsicodendrin as he isolated and identified it in his laboratory. Mr. Nathan Ezzone performed experiments in the ROS assay and the Caspase-Glo assay. Dr. Ulyana Munoz Acuna performed the NF-κB p65 assay, FACS study, the JC-1 assay western blot analysis, analyzed the data and participated in the preparation of the manuscript.
Conflicts of Interest
The Authors declare no conflicts of interest in relation to this study.
- Received October 8, 2021.
- Revision received November 12, 2021.
- Accepted November 18, 2021.
- Copyright © 2021 International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.