


Abstract
Background/Aim: Liver cancer is the fourth leading cause of cancer-related mortality globally, of which hepatocellular carcinoma (HCC) accounts for 85-90% of total primary liver cancer. A drug shortage for HCC therapy triggered us to screen the small-molecule database with a high-throughput cellular screening system. Herein, we examined whether cetyltrimethylammonium bromide (CTAB) inhibits cellular mobility and invasiveness of Mahlavu HCC cells. Materials and Methods: The effects of CTAB on cell viability were assessed using WST-1 assay, cell-cycle distribution using flow cytometric analysis, migration/invasion using woundhealing and transwell assays, and associated protein levels using western blotting. Results: Treatment of Mahlavu cells with CTAB transformed its mesenchymal spindle-like morphology. In addition, CTAB exerted inhibitory effects on the migration and invasion of Mahlavu cells dose-dependently. CTAB also reduced the protein levels of matrix metalloproteinase-2 (MMP2), MMP9, RAC family small GTPase 1, SNAIL family transcriptional repressor 1 (SNAI1), SNAI2, TWIST family basic helix-loop-helix transcription factor 1 (TWIST1), vimentin, N-cadherin, phospho-fibroblast growth factor (FGF) receptor, phospho-phosphoinositide 3-kinase, phospho-v-Akt murine thymoma viral oncogene and phospho-signal transducer and activator of transcription 3 but increased the protein levels of tissue inhibitor of metalloproteinases-1/2 and E-cadherin. Rescue experiments proved that CTAB induced mesenchymal–epithelial transition in Mahlavu cells and this was significantly dose-dependently mitigated by basic FGF. Conclusion: CTAB suppressed the migration and invasion of Mahlavu cells through inhibition of the FGF signaling pathway. CTAB seems to be a potential agent for preventing metastasis of hepatic cancer.
Liver cancer is the fourth leading cause of cancer-related mortality globally (1). Approximately 75% of cases of primary tumor of the liver are classified as hepatocellular carcinoma (HCC), for which the 5-year survival rate is estimated to be only 12% (2). In spite of therapies including surgical excision and emerging treatment strategies, such as molecular targeted therapy, the prognosis for patients with HCC is poor. The reasons for this are attributed to its fast progression, the rapid development of intrahepatic and extrahepatic metastasis, and the absence of efficient therapy, therefore leading to recurrence (3, 4). Recurrence is an important challenge in the treatment of HCC and is the leading cause of death (5). Repression of cancer cell metastasis is recognized as a critical factor in therapeutics of cancer (6). Therefore, new potential approaches or medicines must be explored to prevent metastasis of HCC.
Exploring the initiation of metastasis for cancer progression, studies have shown that there must be changes in the physiological state of the cells, including epithelial to mesenchymal transition (EMT), subsequently mesenchymal to epithelial transition (MET), and finally leading to colonization at other sites (7). Thus, disruption of the mesenchymal characteristics of cancer cells is supposed to be the prime prerequisite for diminishing cancer cell motility and invasiveness (7, 8). Several transcription factors including SNAIL family transcriptional repressor 1 (SNAI1, formerly SNAIL), SNAI2 (formerly SLUG), zinc finger E-box binding homeobox 1 (ZEB1), and twist family basic helix-loop-helix transcription factor 1 (TWIST1), which are modulated by complicated signaling pathways, such as phosphoinositide 3-kinase (PI3K), v-akt murine thymoma viral oncogene homolog 1 (AKT) and signal transducer and activator of transcription 3 (STAT3), have been identified as playing decisive roles in EMT (9). Moreover, molecular mechanisms of EMT involve large numbers of cellular components through multiple signaling networks, such as those of extracellular matrix (ECM) and matrix metalloproteinases (MMPs). In the processes of EMT, MMPs play a critical role in interacting with and degrading the basement membrane of ECM to remodel it for invasion and migration of metastatic cells to target organs, thereby contributing to tumor cell metastasis (10, 11). In addition, studies have shown MMP2 and MMP9 to be associated with HCC progression (12, 13). Thus, MMP2 and MMP9 might be used as the target proteins for screening small-molecule compounds in the treatment of HCC.
Rac family small GTPase 1 (RAC1), a member of the Rho GTPase family which also includes Ras homolog gene family member A (RhoA) and cell-division control protein 42 homolog (CDC42), has been reported to be involved in cell cytoskeletal remodeling and mesenchymal-like migration (14). RAC1 plays a pivotal role during malignant transformation, including in angiogenesis, invasion, tumorigenesis, and metastasis (15). A positive correlation between metastatic potential and endogenous RAC1 activity have been reported in breast cancer, urothelial carcinoma of the upper urinary tract, head and neck squamous cell carcinoma, and glioblastoma (16-19). Additionally, activation of the RAC1 signaling pathway by both activating mutations in Lin-11, Isl-1 and Mec-3 domains and cysteine-rich domains 1 protein, a downstream target of RAC1, and down-regulation of microRNA-142 has been demonstrated to increase the metastatic ability of HCC (15, 20). RAC1 therefore appears to regulate numerous critical elements of metastasis. According to these consistent findings, therapeutic agents that are capable of inhibiting designated components of RAC1 signaling may be efficacious.
In our previous work, cetyltrimethylammonium bromide (CTAB) was found to have strongly suppressive effects on the migration and invasion of different HCC cell lines, such as SK-HEP1, Mahlavu and HA22T/VGH. Our study demonstrated that treatment of SK-HEP1 cells with CTAB altered cellular mesenchymal spindle-like morphology by suppression of the canonical and non-canonical transforming growth factor β signaling pathways (21). However, the detailed mechanisms of the effects of CTAB on the migration and invasion of Mahlavu cells have not been examined. Therefore, the aim of the present study was to explore its potential molecular mechanisms.
Materials and Methods
Cell culture. The Mahlavu cell line was obtained from the American Type Culture Collection (Manassas, VA, USA). Mahlavu cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA, USA) with 10% fetal bovine serum (FBS; Hyclone Laboratories, Logan, UT, USA), 100 μM non-essential amino acids (Gibco), 100 units/ml penicillin, and 100 μg/ml streptomycin (Invitrogen, Carlsbad, CA, USA) in a humidified atmosphere containing 5% CO2 at 37°C. CTAB was purchased from Sigma Chemicals (St. Louis, MO, USA). Basic fibroblast growth factor (bFGF) was purchased from PeproTech (Rocky Hill, NJ, USA). Both CTAB and bFGF were dissolved in phosphate-buffered saline (PBS; Invitrogen, Carlsbad, CA, USA) as a stock solution.
Cell viability. Cell viability was analyzed by measuring the activity of mitochondrial malate dehydrogenase using the WST-1 assay (22). Cell proliferation reagent WST-1 was purchased from BioVision (Milpitas, CA, USA). Cell proliferation was determined at 24 h after treatment with 0, 1.0, 2.5 and 5.0 μM of CTAB by adding WST-1 reagent and incubating the cells for a further 3 h. The amount of formazan, which was proportional to the number of viable cells, was measured using the absorbance signal at 440 nm with an enzyme-linked immunosorbent assay microplate reader. The absorbance was corrected in accordance with a background reading.
Cell-cycle analysis. After treatment with 0, 1.0, 2.5 and 5.0 μM of CTAB, cells were washed twice with PBS and were then fixed in 70% ethanol overnight at −20°C. Fixed cells were stained with a solution containing 10 μg/ml of propidium iodide (Sigma), 100 μg/ml of RNase A (Sigma) and 0.1% Triton X-100 (Sigma) at 37°C in the dark. After 30 min incubation, cell-cycle distribution was analyzed using FACScan (BD Biosciences, Bedford, MA, USA) flow cytometry.
In vitro migration and invasion assays. The migration assay was performed using Ibidi Culture-Insert (Ibidi GmbH, Munich, Germany) and Millicell 24-well hanging inserts with 8 μm pores of polyethylene terephthalate membrane (EMD Millipore, Billerica, MA, USA). The upper side of Millicell 24-well hanging inserts was pre-coated with 50 μl/insert of Matrigel (1 mg/ml; BD Biosciences) for the invasion assay. Cells with CTAB treatment were placed in the upper part of the chamber, and the lower compartment was filled with DMEM containing 20% FBS. The experimental procedures were identical to the migration assay procedures (21). In addition, the present study also examined whether the effects of CTAB treatment on Mahlavu cells could be fully or partially rescued by PI3K agonist/cognate ligand of fibroblast growth factor receptor (FGFR), bFGF. Mahlavu cells pre-incubated with 10 or 20 ng/ml bFGF, for 16 h were subsequently treated with CTAB for another 16 h and then migration and invasion assays were performed.
Immunoblotting analysis. Cells were lysed in radio immuno-precipitation assay lysis buffer (EMD Millipore) containing protease and phosphatase inhibitors (Roche, Mannheim, Germany). Primary antibodies against the following were used: MMP2, MMP9, tissue inhibitors of metalloproteinase 1 (TIMP1), TIMP2, RAC1, SNAI1, SNAI2, TWIST1, vimentin, N-cadherin, E-cadherin, p-FGFR (Tyr653/654), FGFR, p-PI3K p85 (Tyr458), PI3K p85, p-AKT (Ser473), p-AKT (Thr308), AKT, p-STAT3, STAT3, and β-actin. All antibodies were purchased from Cell Signaling Technology (Beverly, CA, USA). Equal amounts of protein extracts were subjected to immunoblot following methods described previously (23).
RAC1 activation assay. Endogenous active GTP-bound RAC1 levels were determined by Active RAC1 Pull-down and Detection Kit (Thermo, Rockford, IL, USA). The p21-binding domain (PBD) of the target PAK1, fused to glutathione S-transferase (GST) (GST-PBD), was used to bind RAC1-GTP in cell lysates. Briefly, RAC1-GTP bound to GST-PBD was immobilized on a resin, and the active GTPase eluted with 2X sample buffer. Levels of total RAC1 and active RAC1 were then determined by immunoblotting with RAC1 antibody (24).
Statistical analysis. The data are expressed as mean±standard errors (SE) of three independent experiments and were analyzed by one-way analysis of variance (ANOVA) using the GraphPad prism software program (Graph Pad Software, La Jolla, CA, USA). Student's t-test was used in two-group comparisons. A value of p<0.05 was considered as statistically significant.
Results
Morphological changes induced by CTAB. In order to clarify the role of CTAB in EMT, Mahlavu cells were treated with CTAB for 24 h to track morphological cellular changes in the process of treatment. In accordance with the phenotypic changes of cells, 5 μM CTAB-treated Mahlavu cells transformed from fibroblast-like cells (see control cells) to cells with cobblestone-like morphology (Figure 1A).
Cytotoxicity of CTAB to Mahlavu cells. Next, we investigated the ability of CTAB to inhibit cell proliferation and survival. Cell viability was assessed by WST-1 assay after Mahlavu cells were incubated with vehicle (PBS) or different concentrations of CTAB for 24 h. As shown in Figure 1B, CTAB did not significantly inhibit the viability of Mahlavu cells. Dosage of CTAB up to 5 μM also retained more than 90% cell viability (Figure 1B).
To investigate the effect on the cell-cycle distribution, the relative DNA content at different phases (sub G1, G0/G1, S and G2/M) were determined by flow cytometry. After 24 h of exposure to 1 μM, 2.5 μM, and 5 μM CTAB, there were no significant differences of the cell-cycle distribution in CTAB-treated Mahlavu cells (Figure 1C) compared to the control. Altogether, CTAB had no great influence on growth and survival through modulation of the cell cycle or apoptosis (sub-G1 phase).
CTAB inhibits the migratory and invasive abilities of Mahlavu cells. Treatment with CTAB led to morphological change in Mahlavu cells, which displayed a round shape as compared to the control cells. Morphological changes induced by CTAB treatment was considered as being related to the progression of a mesenchymal phenotype to an epithelial phenotype. Thus, wound-healing and transwell chamber assays were performed to examine the effects of CTAB on the migration of Mahlavu cells. As shown in Figure 2A, in the CTAB-treated group, fewer cells migrated to the cell-free zone compared to the control cells. In addition, Figure 2B revealed that the number of migratory CTAB-treated Mahlavu cells was reduced to 28.3±5.17% at 5 μM when compared with the control group. Next, Transwell chambers coated with Matrigel were used to analyze the effects of CTAB on Mahlavu cell invasion. In comparison with the control group, the results of the assay showed that the relative invasiveness of Mahlavu cells treated with 5 μM CTAB was significantly reduced to 17.8±4.63% (Figure 2C). These data suggest that CTAB may diminish the migratory and invasive capacities of Mahlavu cells.
CTAB attenuates the expression of MMP2 and MMP9. MMPs are known to play an important role in the degradation and removal of ECM molecules, cell–matrix interaction and tumor invasion (25, 26). To determine whether CTAB is involved in the expression of MMP2 and MMP9 proteins in Mahlavu cells, western blot analysis was performed. The results showed that treatment of CTAB led to down-regulation of MMP2 and MMP9 protein levels but up-regulation of tissue inhibitors of metalloproteinase 1 (TIMP1) and TIMP2 protein levels in Mahlavu cells (Figure 3A). The variation between down-regulation of MMPs and up-regulation of TIMPs might be a result of the dynamic balance in restraining the invasiveness of Mahlavu cells.
CTAB inhibits RAC1 activation. RAC1 has been implicated in cell motility of multiple cancer types, and it is presumable that RAC1 might be regulated by CTAB in Mahlavu cells (27). In order to evaluate the relationship between anti-tumorigenesis effect of CTAB and the RAC1 signaling, we used a PAK1 PBD pull-down assay to measure the levels of active, GTP-bound RAC1. Our data showed that treatment of Mahlavu cells with CTAB caused a reduction in the level of RAC1-GTP compared to the control, suggesting that CTAB suppressed RAC1 activity in a dose-dependent manner (Figure 3B).
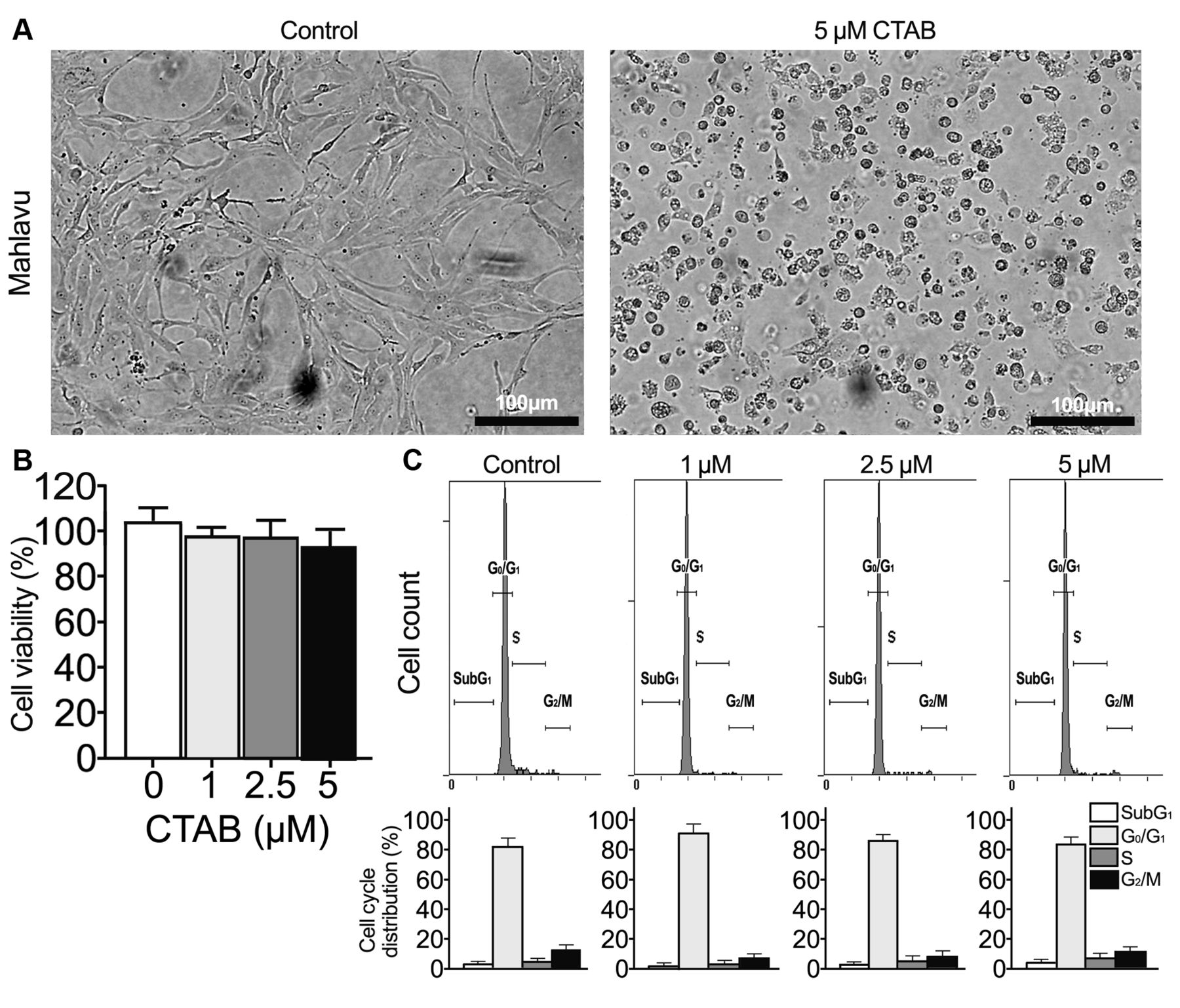
Effect of cetyltrimethylammonium bromide (CTAB) on morphological alterations of Mahlavu cells. A: Mahlavu cells were cultured with CTAB for 24 h. Images were captured at 24 h (scale bars: 100 μm). B: Cells were exposed to different concentrations of CTAB (0, 1.0, 2.5, and 5.0 μM) or an equal volume of phosphate-buffered saline (vehicle; control) for 24 h. Cell viability was then determined by the WST-1 assay. Absorbance values obtained from Mahlavu cells treated with vehicle were taken as 100%. C: Distribution of cells in subG1, G0/G1, S and G2/M phases of the cell cycle under different treatment conditions. Values are the mean±SE of the percentage of cell subgroups from flow cytometric analysis using propidium iodide staining. Data are representative of three independent experiments, and performed in duplicates.
CTAB reverses expression of EMT markers in Mahlavu cells. EMT and its related markers have been reported to be highly associated with cell migration and invasion in HCC (28). Thus, we assessed the expression levels of EMT-related proteins and the effect of CTAB on EMT in Mahlavu cells. We found that treatment of CTAB in Mahlavu cells dose-dependently increased the expression of E-cadherin and reduced the protein expression of SNAIL, SLUG, TWIST, vimentin, and N-cadherin (Figure 3C). Compared to control cells, Mahlavu cells treated with CTAB reduced expression of mesenchymal markers concomitant with an elevated expression of the epithelial marker.
CTAB inhibits the expression of PI3K/AKT and STAT3 in Mahlavu cells. To elucidate the possible signaling pathways involved in anti-EMT triggered by CTAB in Mahlavu cells, we performed immunoblot analysis to screen the expression levels of particular proteins associated with potential signaling pathways of MET, such as Receptor tyrosine kinases, NOTCH, wingless-type MMTV integration site family (WNT), hedgehog and others (21, 29). Immunoblot analysis revealed that CTAB treatment evidently attenuated the phosphorylation of proteins, including of PI3K, AKT, STAT3 and FGFR but there were no detectable changes in any of the non-phosphorylated forms of the proteins (Figure 4). These results suggested that the inhibitory effect of CTAB might be through the FGFR, PI3K/AKT and STAT3 signaling pathways.

Effect of cetyltrimethylammonium bromide (CTAB) on the migratory and invasive capacities of Mahlavu cells in vitro. A: Cells were treated with vehicle or 5 μM CTAB for 24 h, and wound-healing assay using Ibidi Culture-Insert was then performed to assess cell migration capacity. B: Anti-migratory and C: anti-invasion effects of CTAB on Mahlavu cells was examined by transwell migration and Matrigel invasion assays with and without CTAB treatment. Data are means±SE (n=3), and experiments were performed in duplicates. Scale bars: 200 μm. *Significantly different at p<0.05 versus the control.

Cetyltrimethylammonium bromide (CTAB) influences the expression of metastasis-associated proteins. Mahlavu cells were treated with vehicle or CTAB (1.0, 2.5 and 5.0 μM) for 16 h, and then whole-cell extracts were subjected to western blot analysis. A: Representative western blot images reflecting the protein levels of matrix metalloproteinase-2 (MMP2), MMP9, tissue inhibitor of metalloproteinases-1 (TIMP1) and TIMP2 in Mahlavu cells. B: The active GTP-bound form of Rac family small GTPase 1, RAC1-GTP, was analyzed with pull-down assay and total RAC1 protein was detected separately. C: Western blot analysis demonstrated levels of epithelial to mesenchymal transition-related markers SNAIL family transcriptional repressor 1 (SNAI1, formerly SNAIL), SNAI2 (formerly SLUG), twist family basic helix-loop-helix transcription factor 1 (TWIST1), vimentin, N-cadherin and E-cadherin in CTAB-treated Mahlavu cells. β-Actin was used as a loading control. The results are representative of three independent experiments.
PI3K/AKT/STAT3 signaling axis is involved in CTAB-mediated MET. There is accumulating evidence that suggests EMT is a process which not only invoIves complex multi-gene regulation but also potential a comprehensive change in gene expression. The next step of this study was to conform whether CTAB exerts an inhibitory effect on FGFR and its downstream target genes involved in EMT. Therefore, Mahlavu cells pre-incubated with PI3K agonist/cognate ligand of FGFR, bFGF, were subsequently treated with CTAB (30). Our results showed that the combined treatment with CTAB and bFGF restored the expression of mesenchymal markers and MMP2, as well as cell migration and invasion (Figure 5). Taken together, these results suggest that the FGF/PI3K/AKT signaling pathway is involved in CTAB-modulated suppression of Mahlavu cell motility and invasiveness.
Discussion
HCC is one of the significant causes of cancer-related death in the Asia-Pacific region (31). It has been confirmed that early detection increases the chance of potentially curative treatment of HCC but in fact it is too difficult to diagnose the early stages of HCC. This is because of the absence of early observable symptoms in most patients with HCC and the common presentation of advanced or terminal cancer at initial diagnosis (32). Even though local ablation, surgical resection, liver transplantation, catheter-based locoregional therapy, kinase or immune checkpoint inhibitors has improved survival rates in early-, intermediate-, and advanced-stage HCC, local recurrence and distant metastasis are still responsible for the majority of HCC-associated mortality. Because various unclear aspects affect the metastasis of cancer cells, progress in the treatment of cancer metastasis is limited.
Exploring the process of metastasis, metastatic cells begin with detaching from the original tumor, then migrating, invading to different sites via blood vessels or lymph vessels, and finally adhering and proliferating in a distant region. During the course of metastasis, the tumor microenvironment mediates aberrant tissue function and modulates subsequent progression of tumor. The tumor microenvironment, containing multi-network of cell types, growth factors, chemokines, MMPs and ECM structures, plays a significant role in tumor metastasis (7). The parameters involved in principal metastatic processes and maintenance of the tumor microenvironment have been suggested to be targets for anti-metastatic agents. To treat metastasis effectively, novel therapeutic schemes and development of new drugs targeting metastatic progression are indispensable.
In this study, we investigated the pharmacological activity of CTAB on Mahlavu cell mobility, including migratory and invasive capacities. CTAB, classified as a quaternary ammonium compound, is widely used as bacteriostatic and bactericidal agent. Quaternary ammonium derivatives, such as dequalinium chloride and benzalkonium chloride, inhibit tumor cell proliferation via reducing the mitochondrial respiration rate by inhibiting NADH-ubiquinone reductase activity (33, 34). Moreover, CTAB exhibits cytotoxicity against head and neck cancer cells through depolarization of the mitochondrial membrane potential (35). In addition, our previous data suggested that CTAB obviously reduced the migration and invasion of SK-HEP1 hepatic adenocarcinoma cells (21). However, the detailed mechanisms involved were unclear.
In the present study, we found that CTAB distinctly reduced the migration and invasion of Mahlavu cells. Moreover, WST-1 assay showed that CTAB did not strongly affect cell viability at a concentration up to 5 μM. We also found that CTAB treatment significantly attenuated the protein levels of MMP2/9, and the active form of RAC1-GTP, as well as reversing the EMT process, and inhibiting the activation of PI3K/AKT and STAT3 signaling pathway, which was associated with the inhibitory effect of CTAB on Mahlavu cell migration and invasion. Migration and invasion of tumor cells require the degradation of ECM and then interaction with stromal cells and ECM of other cells (36). During the above process, MMPs secreted from tumor cells participate in disruption of basement membrane components, such as fibronectin, collagen, and laminins (37). In addition, MMPs are the major proteinases regulating ECM remodeling. It has been reported that increasing the expression levels of MMP2/9 can modulate the malignant progression of HCC, and thus are considered as important enzymes in HCC development (38, 39). In this study, CTAB inhibition of MMP2/9 protein expression might have attenuated the mobility of Mahlavu cells.
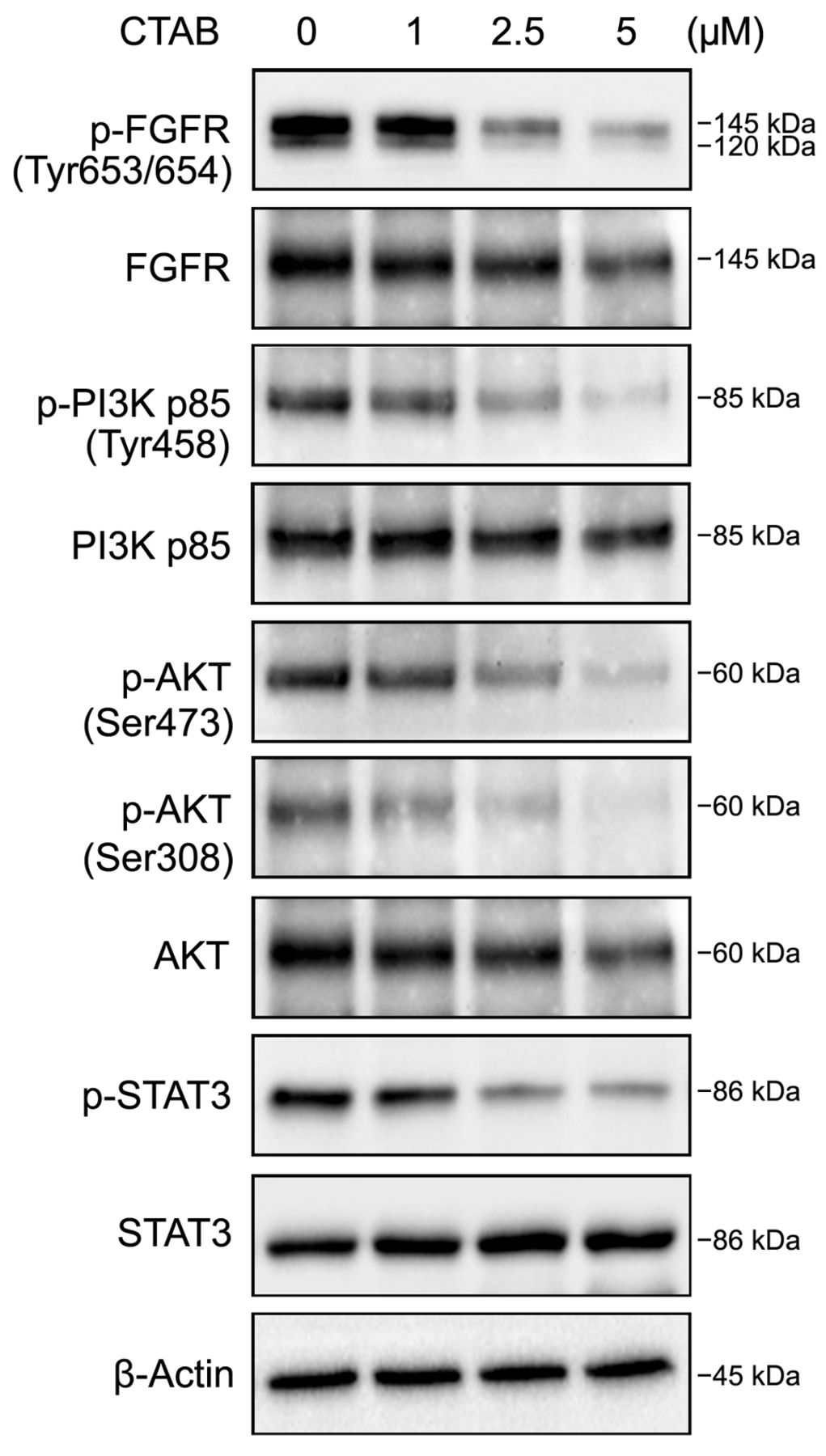
Effect of cetyltrimethylammonium bromide (CTAB) on phosphoinositide 3-kinase (PI3K)/v-Akt murine thymoma viral oncogene (AKT) and signal transducer and activator of transcription 3 (STAT3) signaling pathways. Whole-cell lysates of CTAB-treated Mahlavu cells were subjected to immunoblot analysis for determining levels of total and phosphorylated forms of fibroblast growth factor receptor (FGFR), PI3K, AKT and STAT3 proteins. β-Actin was used as a loading control. The results are representative of three independent experiments.

Cetyltrimethylammonium bromide (CTAB) affects the levels of epithelial to mesenchymal transition (EMT)-associated proteins in and phosphoinositide 3-kinase (PI3K)/v-Akt murine thymoma viral oncogene (AKT)/signal transducer and activator of transcription 3 (STAT3)-related pathways on Mahlavu cells. Cells were pretreated with vehicle or 10-20 ng/ml PI3K/AKT agonist, basic fibroblast growth factor (bFGF) for 16 h, and then co-treated with CTAB for another 16 h. A: Expression of EMT markers and PI3K/AKT signaling-related proteins in CTAB-treated cells with and without bFGF were analyzed using western blot. β-Actin was reprobed to confirm equal loading. B: Mahlavu cells were precultured with bFGF for 16 h prior to addition of CTAB for 16 h and then their morphological changes were imaged. Scale bars: 200 μm. bFGF restored CTAB-inhibited motility as analyzed using wound-healing (C), transwell migration (D) and Matrigel invasion (E) assays. The invasiveness of CTAB-treated cells was examined under the transwell system and the results are presented as a percentage of the control (100%). SNAI1: SNAIL family transcriptional repressor 1 (formerly SNAIL), SNAI2 (formerly SLUG). Significantly different at *p<0.05 and **p<0.01, when compared with the control group.
Clinically, up-regulation of RAC1 in HCC is correlated with advanced clinical stage, poor histological differentiation and other malignant characteristics (40). In addition, activation of the RAC1/AKT cascade is a risk factor associated with recurrence as well as poor prognosis of HCC (41). A recent study demonstrated that the increase of RAC1-GTP through activation of the STAT3 is related to the promotion of EMT-mediated invasion (42, 43). Our study showed that CTAB suppressed the activity of RAC1-GTP, and inhibited the activation of PI3K/AKT and STAT3 proteins in Mahlavu cells.
Previous studies have indicated that the transition of mesenchymal cells into an epithelial state through MET reduces migratory features. This change can lead to increased epithelial cell attachment, gain of cytoskeleton composition, and loss of mesenchymal composition (44). The transformation of MET includes the gain of expression of epithelial-associated genes such as E-cadherin and loss of its transcriptional repressors SNAI2 and TWIST (45, 46). In this regard, our results were consistent with the suppression of transcription factors of mesenchymal-associated genes and resultant increase of epithelial-associated genes. Both the morphological and molecular changes suggest that Mahlavu cells might undergo reversal of EMT with exposure to CTAB.
The PI3K/AKT signaling cascade is the main component upstream of EMT and is involved in metastasis, and tumor invasion in HCC (46). The PI3K/AKT signaling pathway is also involved in MMP-mediated cell invasion (47). Furthermore, bFGF is able to promote EMT through AKT/glycogen synthase kinase-3β/SNAI signaling, resulting in the induction of tumor cell motility (48). Our data showed that the treatment with CTAB regulated cellular signaling pathways, such as the FGFR and PI3K/AKT pathways (Figure 4). CTAB might cause the down-regulation of PI3K/AKT through FGFR signaling, thereby reducing migratory and invasive abilities of human Mahlavu liver cancer cells.
In conclusion, we reported for the first time that CTAB treatment led to inactivation of FGFR/PI3K/AKT signaling and caused mesenchymal transition to epithelial characteristics in Mahlavu HCC cells. Our results also showed that bFGF treatment led to restoration of PI3K/AKT signaling and re-transition of cells to cells with mesenchymal characteristics. Our findings illustrate the anti-metastatic effect of CTAB, and point to the potential therapeutic value of CTAB for the treatment of HCC.
Acknowledgements
This study was supported by the Research Fund of Tung's Taichung MetroHarbor Hospital (TTMHH-108R0028 and TTMHH-108R0051) and Chang Bing Show Chwan Memorial Hospital (BRD108016).
Footnotes
↵* These Authors contributed equally to this study.
Authors' Contributions
Conceptualization was by CJL, funding acquisition by TKW and CHC, Methodology and validation by YRP, WTL and FMH. Writing of the original draft was by FMH and CJL, reviewing and editing by TFS and CJL. All Authors read and approved the final article.
Conflicts of Interest
All Authors declare that they have no conflicts of interest in regard to this study.
- Received April 24, 2020.
- Revision received June 7, 2020.
- Accepted June 10, 2020.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.