


Abstract
Background/Aim: We investigated the anti-proliferative effect of quercetin on liver cancer cell lines. Materials and Methods: Thirteen liver cancer cell lines were cultured followed by treatment with varying concentrations of quercetin (0-100 μM) or quercetin and 5-FU, and the cell viability was analysed by the MTT assay. Flow cytometry was also used to examine cell cycle progression after treatment with quercetin. Results: The addition of quercetin resulted in a dose- and time-dependent suppression of cell proliferation. In some cell lines, treatment with quercetin and 5-FU caused an additional or synergistic effect. Most cell lines displayed cell cycle arrest at different phases of the cell cycle. Conclusion: Quercetin inhibits the proliferation of liver cancer cells via induction of apoptosis and cell cycle arrest.
Hepatocellular carcinoma (HCC) is the most frequent primary cancer, and an important medical problem (1). Many HCC patients have a history of chronic hepatitis or liver cirrhosis caused by Hepatitis B virus (HBV) or Hepatitis C virus (HCV) infection (2-4). Treatments for poor liver function associated with hepatitis are limited and outcomes are poor. Even with the most successful treatments, the 5-year recurrence rate of HCC is very high (1). Treating and preventing recurrence is critical for improving survival rates.
Quercetin is a type of flavonoid contained in many plants, and possesses antioxidant, anti-inflammatory, and immunological capabilities (5). Quercetin can suppress cell proliferation and induce apoptosis in human cancers, including breast, lung, oral, and prostate; however, there is no report on its effect on liver cancer (6-9). In the current study, we examined the antitumor effects of quercetin on 13 HCC cell lines in vitro.
Materials and Methods
Cell lines and cell culture. This study utilized 11 HCC cell lines (KIM-1, KYN-1, KYN-2, KYN-3, HAK-1A, HAK-1B, HAK-2, HAK-3, HAK-4, HAK-5, and HAK-6), and 2 human combined hepatocellular and cholangiocarcinoma (CHC) cell lines (KMCH-1 and KMCH-2). The cell lines were originally established in the Department of Pathology, Kurume University Faculty of Medicine, and each of them retains the morphological and functional features of the original tumor as previously described (10-18). Each cell line was grown in Dulbecco's modified Eagle medium (Nissui Seiyaku, Co., Japan) supplemented with 2.5% heat-inactivated (56°C, 30 min) fetal bovine serum (Bioserum, Victoria, Australia), 100 U/ml penicillin, 100 μg/ml streptomycin (GIBCO BRL/Life Technologies, Inc., Gaithersburg, MD, USA) and 12 mmol/l sodium bicarbonate, in a 5% CO2 humidified atmosphere at 37°C.
Effects of quercetin on the proliferation of hepatocellular carcinoma and combined hepatocellular and cholangiocarcinoma cell lines in vitro. The effects of quercetin with and without 5-FU on proliferation were examined using 3-(4,5-dimethylthiazol-2yl-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay kit (Chemicon, Temecula, CA, USA) as previously described (10). Quercetin was obtained from Sigma Chemical, and 5-fluorouracil (5-FU) was purchased from Kyowa Hakko (Tokyo, Japan). Briefly, cells (1.5-8×103 cells per well) were seeded on 96-well plates (Falcon, Becton Dickinson Labware, Tokyo, Japan), cultured for 24 h, and then culture medium was replaced with a new one containing quercetin (0, 12.5, 25, 50, 100 μM) with or without 5-FU (0, 10, 100 μM). After treatment of cells for 24, 48 or 72 h, the number of viable cells was examined.
Quantitative analysis of apoptosis induced by quercetin in vitro. Cells cultured with or without quercetin for 72 h were stained with Annexin V-EGFP (enhanced green fluorescent protein) and analysed by using an Apoptosis detection kit (Medical and Biological Laboratories Co., Ltd. Nagoya, Japan) according to the manufacturer's protocol. After staining, the cells were analyzed using a FACScan (Becton Dickinson Immunocytometry Systems Franklin Lakes, NJ, USA), and annexin V-EGFP-positive apoptotic cell rate was determined.
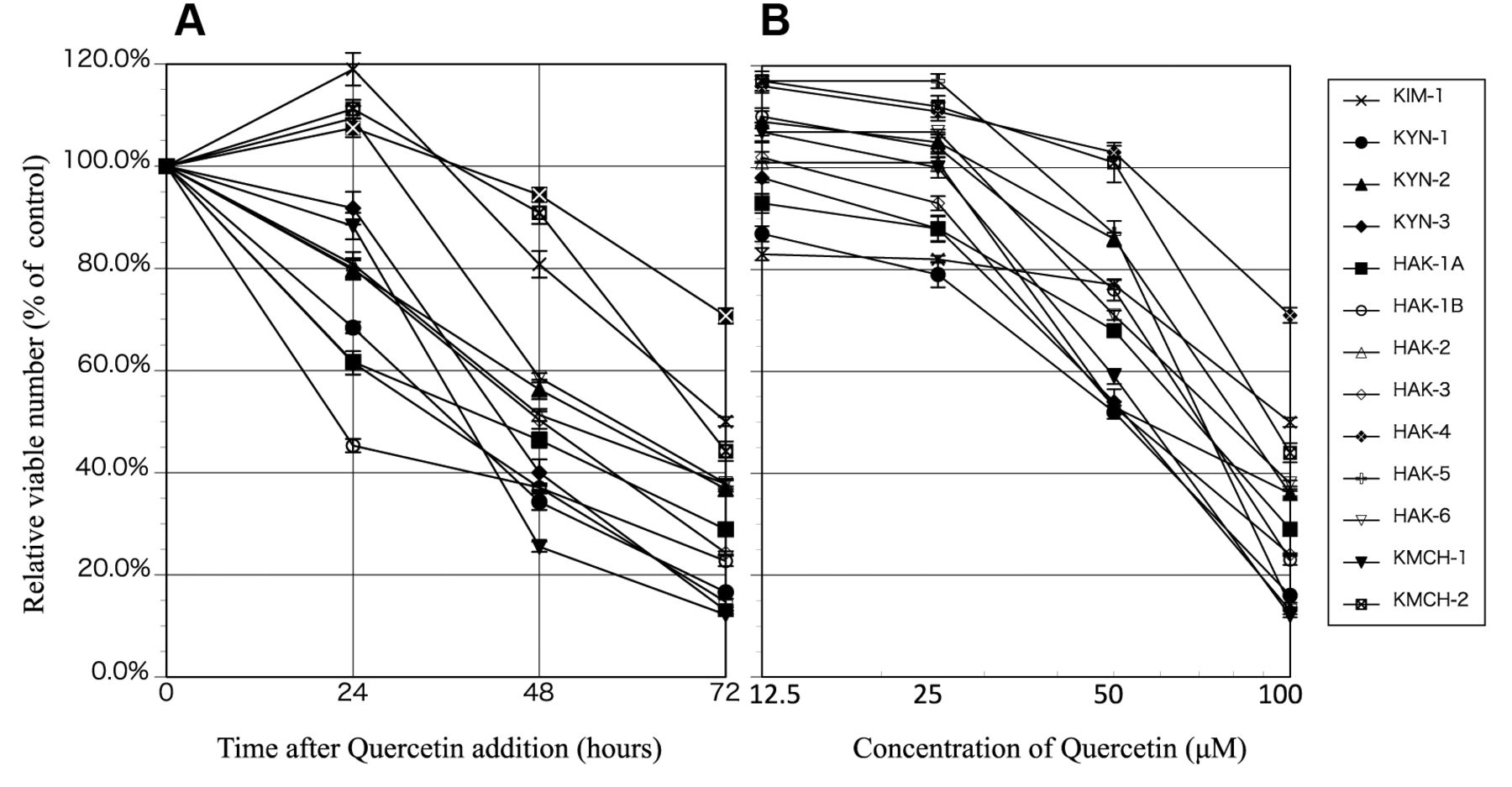
Effect of quercetin on the proliferation of liver cancer cell lines. (A) Relative viable cell number (% of the control) was reduced in a time-dependent manner after the addition of 100 μM of quercetin. (B) 72 h after adding 12.5, 25, 50 or 100 μM of quercetin cell proliferation was suppressed in a dose-dependent manner but in varying degrees. The suppression was statistically significant (p<0.01-0.0001) in the range of 12.5-100 μM of quercetin in all cell lines. Eight samples were used in each experiment (n=8). The experiment was repeated at least three times for each cell line to confirm reproducibility of the data, and almost identical results were obtained. Error bar represents SE. Mean absorbance of the control cells (cells cultured without quercetin) was determined as 100%, and the mean of the other groups was expressed as a ratio.
Cell cycle analysis. Cell lines were cultured with or without quercetin (25 or 100 μM) for 48 h, labeled with 10 μmol/l BrdU for 30 min, fixed in 70% cold ethanol at 4°C overnight, stained with anti-BrdU and propidium iodide (Sigma Chemical Co.), and analyzed using a FACScan. Staining was performed using the modified technique described elsewhere (19). Briefly, fixed cells were washed with PBS, subjected to double-strand DNA denaturation treatment with 2 mol/l HCl at room temperature for 30 min, neutralized with 0.1 mol/l Na2B4O7, washed twice with PBS containing 0.5% Tween 20, and 0.5% BSA (PBS-T), incubated for 30 min at room temperature with 20 μl anti-BrdU antibody, and washed with PBS-T. Then, cells were incubated for 30 min at room temperature with 4 μl fluorescein isothiocyanate–conjugated goat anti-mouse immunoglobulin, and washed with PBS-T. DNA was counterstained with 5 μg/ml of propidium iodide for at least 20 min before flow cytometric analysis. Percentage of cells in the G0/G1, S, and G2/M phases of the cell cycle was calculated from a dot or contour plot obtained after flow cytometric analyses of double-stained cells.
Statistics. Significance in differences was estimated by using the Student's t test (two-tailed).
Results
In all 13 cell lines, the viable cell number reduced to around 20 to 70%, after 72 h of treatment with 100 μM of quercetin. In four of the cell lines (KIM-1, HAK4, HAK-6, KMCH-1), the cell proliferation reached a plateau 24 h after quercetin addition, and there was a reduction in their relative viable cell numbers thereafter. In the other 9 cell lines, there were varying degrees of cell growth inhibition over the course of 72 h since quercetin addition (Figure 1A). All cell lines showed a dose-dependent decrease in the relative viable cell count 72 h after quercetin addition (Figure 1B). The relative viable cell count for cells cultured with 100 μM of quercetin ranged between 12 to 70%, with a mean of 31.4±17%. In 10 cell lines (KYN-1, KYN-2, KYN-3, HAk-1A, HAK-1B, HAK-2, HAk-3, HAK5, HAK-6, KMCH-1, KMCH-2), the relative viable cell number reduced to below 50%. Quantitative analysis of Annexin V-EGFP-positive apoptotic cells revealed that the appearance of apoptosis was significantly higher in the cells treated with quercetin compared to untreated cell lines (Table I).
Quantitative analysis of apoptosis induced by quercetin in 13 liver cancer cell lines.

Effect of quercetin on the cell cycle distribution of 13 liver cancer cell lines. Cells were treated with 0-100 μM quercetin for 48 h and analyzed by flow cytometry. Cell cycle distribution is shown as a histogram. Most liver cancer cell lines underwent cell cycle arrest, but the phase of arrest varied from cell line to cell line (A: G0/G1 phase arrest, B: G2/M phase arrest, C: S phase arrest).
We examined the effect of quercetin on the cell cycle of liver cancer cell lines. Cell cycle arrest was observed in 12 out of 13 cell lines. Following the addition of quercetin, a dose-dependent G0/G1 phase cell cycle arrest was observed in 4 cell lines (HAK-3, HAK-6, KMCH-1, KMCH-2), in the G2/M phase in 6 cell lines (KYN-2, KYN-3, HAK-1B, HAK-2, HAK-4, HAK-5), and in the S phase in 2 cell lines (KIM-1, KYN-1) (Figure 2).

Effects of quercetin or quercetin+5-FU on the growth of 13 cell line aftertreatment for 72 h. The experiment was repeated at least three times for each cell line to confirm reproducibility of the data, and almost identical results were obtained. Error bar represents SE.
The combination treatment of quercetin and 5-FU produced additional or synergic effects in 9 cell lines (KIM-1, KYN-2, KYN-3, HAK-1B, HAK-2, HAK-5, HAK-6, KMCH-1, KMCH-2) (Figure 3).
Discussion
Quercetin induced a dose-dependent and time -dependent antiproliferative effect in most of the 13 cell lines, and a certain degree of apoptosis in all 13 cell lines. The liver cancer cell lines used in this study had been established from different types of liver cancers, including well to poorly differentiated HCC, sarcomatous HCC, and combined hepatocellular and cholangiocarcinoma. The fact that potent growth inhibition was observed in cell lines of different histology should be emphasized. Although in vitro antiproliferative findings may not exactly resemble those in clinical applications, these findings indicate that quercetin may be efficacious in the treatment of various types of HCC. Experimental reports thus far indicate that quercetin activates caspases, which are involved in the induction of apoptosis, and has an inhibitory effect on the PI3K/Akt and ERK pathways (20). It is possible that the liver cancer cell lines used in this study may have undergone apoptosis and cell growth inhibition by similar mechanisms.
Even though our results on the cell cycle arrest induced by quercetin indicated that most liver cancer cell lines underwent cell cycle arrest, the arrest phase varied from cell line to cell line. Of the 13 liver cancer cell lines studied, 4 cell lines underwent cell cycle arrest in the G0/G1 phase, 2 cell lines in the S phase, and 6 cell lines in the G2/M phase. This is similar to cell cycle arrest by quercetin as reported previously, where the arrest has been shown to take place at different points of the cell cycle (21-23). Quercetin may be able to induce G0/G1 phase, G2/M phase and S phase cell cycle arrest, depending on the type of liver cancer cell line. Further research is needed to determine the exact mechanism.
Treatment with the combination of quercetin and 5-FU led to enhanced growth inhibition in some cell lines, compared to when quercetin was administered on its own. This result was similar to previous reports by Dai et al. demonstrating that 5-FU enhanced quercetin's inhibition of cell growth in vitro and in vivo (24). As the combined use of quercetin and 5-FU showed additional or synergistic effects relative to the use of quercetin alone, this result suggests that the combined use may inhibit the growth of liver cancer at a greater extent. In conclusion, the present study revealed that quercetin suppressed the proliferation of liver cancer cells via induction of apoptosis and cell cycle arrest. Further investigations are needed to clarify the signalling pathways involved in quercetin-induced apoptosis and proliferation inhibition in liver cancer cells.
Acknowledgements
The Authors are grateful to Ms Akemi Fujiyoshi, Ms Mika Hanada and Ms Atsuko Matsuo for their assistance in the experiments.
Footnotes
Authors' Contributions
Toru Hisaka designed the study, and wrote the initial draft of the manuscript. Toru Hisaka and Hisamune Sakai contributed to analysis and interpretation of data, and assisted in the preparation of the manuscript. All other Authors have contributed to data collection and interpretation, and critically reviewed the manuscript. All Authors approved the final version of the manuscript, and agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
This article is freely accessible online.
Conflicts of Interest
The Authors declare no conflicts of interest associated with this manuscript.
- Received May 23, 2020.
- Revision received June 14, 2020.
- Accepted June 15, 2020.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.