


Abstract
Background/Aim: We investigated the clinical role of the molecular targets, APEX1 and Jagged-1, and the Apex1 - Jagged-1 cascade in gastric cancer cells. Materials and Methods: We used 6 human gastric cancer cell lines (SNU-1, SNU-5, SNU-16, NCI-N87, KATO- III and AGS), and demonstrated the chemosensitivity of APEX1 and Jagged-1 through the MTT assay and immunoblotting. Tumor growth was assayed following cisplatin and 5-FU treatment using a xenograft model injected with KATO-III cells. Moreover, gastric tumor samples from 9 patients, divided in 2 groups according to chemotherapy response, were examined by immunocytochemical (IHC) staining, and protein expression levels were scored. Results: Following APEX1 knockdown, the MTT assay revealed that the IC50 of cisplatin and 5-FU in AGS cells was decreased approximately 7% and 15%, respectively, however, their decrease in chemoresistant KATO-III cells was decreased by approximately 21% and 67% for cisplatin and 5-FU, respectively. The tumor volume of KATO-III/sicontrol mice treated with cisplatin and 5-FU was affected less, compared with KATO-III/siAPEX1 mice treated with cisplatin and 5-FU. Also, the expression levels of APEX1, Jagged-1 and CD133, assayed by IHC staining, were higher in the chemorefractory group than in the chemoresponsive group. Conclusion: Jagged-1-activated signaling by APEX1 plays a role in advanced gastric cancer.
Gastric cancer remains one of the most common and deadly cancers worldwide, especially in East Asian countries, including Korea. Although the incidence and mortality rate of gastric cancer has declined considerably in Korea owing to early detection and increased surgical resection, unresectable locally advanced and metastatic gastric cancer still remains uncurable. Cytotoxic chemotherapy has been shown to increase survival in unresectable advanced gastric cancer compared to supportive care only, however, the survival time was less than 12 months (1, 2). One of the major clinical problems associated with cytotoxic chemotherapy is chemoresistance (3). APEX1 is a multifunctional protein that is not only an essential enzyme in the base excision repair pathway, but also acts as a major redox-signaling factor that has a wide variety of important cellular functions including transcription factor regulation, oxidative signaling and cell cycle control. APEX1 stimulates the DNA binding activity of numerous transcription factors that are involved in cancer promotion and progression such as AP-1 (4, 5). Jagged-1 is one of the Notch ligands and Jagged-1-induced Notch activation plays an important role in various aspects of tumor biology (6-8). Several studies have also reported on APEX1-mediated mechanisms of Jagged-1-activated Notch Signaling (9, 10). In this study, we evaluated the clinical significance of Jagged-1 activated by APEX1 in advanced gastric cancer. Our study aimed to investigate the clinical role of the molecular targets, APEX1 and Jagged-1, and the Apex1 - Jagged-1 cascade in gastric cancer cells.
Materials and Methods
Cell culture. Six human gastric cancer cells (SNU-1, SNU-5, SNU-16, NCI-N87, KATO-III, and AGS) were procured from the Korea Cell Line Bank (Seoul, Republic of Korea). The cell lines were cultured in RPMI1640 (WELGENE Inc. Gyeong sang buk-do, Republic of Korea) medium supplemented with 10% FBS (FBS, Lonza, Alpharetta, GA, USA) and 1% penicillin/streptomycin (p/s, Invitrogen, Carlsbad, CA, USA). The cells were maintained in a humidified atmosphere with 5% CO2 at 37°C.
Drug solutions. Cisplatin and 5-FU were obtained as aqueous solutions at concentrations of 0.5 mg/ml and 50 mg/ml, respectively, from JW Pharmaceutical Corp (Seoul, Republic of Korea).
MTT assay. Cell proliferation was measured using the 3-(4, 5dimethylthiazole–2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) assay. Briefly, cell was seeded in 96-well plates. Next day, 10 μl of 5 mg/ml MTT in culture medium were added to each well. The plates were incubated for 3 h at 37°C. Then, culture medium was removed, and 100 μl 36% HCL in isopropanol was added to each well for 10 min and 100 μl of 3 deionized water (DW) were added to each well for 10 min. The absorbance was read on an ELISA plate reader (Tecan, Trading AG, Switzerland) using a 570-nm filter. Cell viability was calculated based on the relative color intensity of the treated and untreated samples.
siRNA-based knockdown of APEX1. To knockdown APEX1 expression, the gastric cancer cells were transiently transfected with APEX1 specific siRNA or negative control siRNA using RNAiMAX (Invitrogen) according to the manufacturer's instructions. The siRNA sequence used to target APEX1 was 5’-AAGTCTGGTACGACTGGAGTA-3’, while that of the negative control siRNA (Bioneer, Daejeon, Republic of Korea) was 5’-CCUACGCCACCAAUUUCGUdTdT-3’.
KATO-III gastric carcinoma xenograft mouse model. The six-week-old female BALB/c nude mice (Orient Bio Inc. Seongnam, Republic of Korea) used in this study, were handled in accordance with standard protocols and animal welfare regulations. Control siRNA-KATO-III cells (1×106 cells), siAPEX1-KATO-III Cells (1×106 cells) were resuspended in IMDM (WELGENE Inc.) media and injected subcutaneously into the left flank of BALB/c nude mouse. Once the tumor reached ~5 mm in size, we initiated treatment with, saline containing 10% dimethyl sulfoxide (s.c. once every three days for 3 weeks), 5-FU (10 mg/kg, s.c. next to the tumor, once every two days for 3 weeks), cisplatin (30 mg/kg, s.c. once every three days for 3 weeks). Tumor volumes were calculated by the following formula tumor volume=[width2 × length]/2. Tumor size was measured with a caliper every 3 days and tumors were excised.
Immunoblotting. Whole cell lysates were prepared in lysis buffer (150 mM Nacl, 100 mM Tris-HCL (pH 7.6), 1% NP-40, 5 mM EDTA, 1 mM dithiothreitol, 1 mM phenylmethyl sulfonyl fluoride, 10 μg/ml leupeptin, 10 μg/ml aprotinin, 1 mM Na3VO4, and 5 mM NaF). The protein concentration of each sample was determined using a dye-binding microassay (Bio-Rad, Hercules, CA, USA). Equal protein amounts of cell or tissue extracts were resolved using 6-12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS–PAGE) followed by the electrophoretic transfer of the proteins onto a polyvinylidene difluoride membrane (PALL Life Sciences, NY, USA). The membranes were blocked for 1 h with Tris-buffered saline-Tween [TBS-T; 10 mM Tris-HCl (pH 7.4), 150 mM NaCl, and 0.1% Tween-20] containing 5% non-fat milk and incubated with specific primary antibodies overnight at 4°C. The blots were washed four times with TBS-T for 15 min per wash and incubated for 1 h with the corresponding peroxidase-conjugated secondary antibodies (1:4000 dilution; Jackson Immuno Research Inc, West Grove, PA, USA). The blots were washed four times with TBS-T and developed using an enhanced chemiluminescence (ECL) detection system (iNtRON Biotechnology, Gyeonggi, Republic of Korea). The antibodies used for western blotting were as follows: mouse anti-APEX1 (sc-17774; Santa Cruz Biotechnology, Dallas, TX, USA), mouse anti-Jagged-1 (sc-390177; Santa Cruz), and rabbit anti-CD133 (ab19898; Abcam, Cambridge, UK).
Analysis of gastric cancer tissue. The in vivo clinical specificity of Jagged-1 activation by APEX1 was analyzed in tissue samples acquired from gastric cancer patients. The protocol was approved by the Chosun University Hospital Ethics Committee. Patients with metastatic or recurrent gastric cancer were enrolled through retrospective chart review. Of these, patients who had received an appropriate first-line chemotherapy and had been evaluated concerning their response, were available for follow-up, had good performance status (Eastern Cooperative Oncology Group 0-1), and normal gastric cancer, were selected. The gastric cancer tissue samples were paraffin embedded and sectioned followed by IHC staining and clinical examination.
Immunohistochemistry. The gastric cancer tissue samples were obtained from the Chosun University Department of Pathology Tissue Bank. The sections were dewaxed with xylene, then rehydrated and prepared for immunohistochemistry by routine methods. The endogenous peroxidase activity was blocked with 0.03% H2O2 for 15 min. The nonspecific binding was suppressed by incubation with 10% normal horse serum (Jackson ImmunoResearch Laboratories Inc., Suffolk, UK) for 1 h at room temperature (20-25°C). Thereafter, the sections were incubated with the primary antibodies, mouse anti-APEX1, mouse anti-Jagged-1, and rabbit anti-CD133 was performed for 24 h at 4°C. For IHC, a biotinylated goat anti-mouse or goat anti-rabbit antibody (Vector Laboratories, Burlingame, CA, USA) followed by horseradish peroxidase-conjugated streptavidin (Vector Laboratories, Burlingame, CA, USA) was employed according to manufacturer's instructions. Labeling was visualized by incubating with 0.05% 3’3-diaminobenzidine (Sigma, St Louis, MO, USA) and 0.01% H2O2 for 5-20 min. The sections were dehydrated through a graded ethanol series, cleared with xylene and mounted under polymount (Shandon, Cheshire, UK). Images were acquired using an Olympus C-4040Z digital camera and an Olympus BX-50 microscope (Olympus Corp, Lake Success, NY, USA). The immunoreactivities of APEX1, Jagged-1, and CD133 were determined by scoring for the staining intensity (0, none; 1, weak; 2; moderate; and 3, strong) and percent of positive cells (0, <5%; 1, 6%-25%; 2, 26%-50%; 3, 50%-75%; and 4, >76%), with the product of the two scores being the final value.
Statistical analysis. Data in all the experiments are presented as the mean±standard deviation (SD). The Student's t-test or ANOVA was used to compare tumor volumes between the two groups. A p-values<0.05 were considered statistically significant. All experiments were repeated more than three times, and the results from representative experiments are shown. The analysis was performed using Graph Pad Prism (Graph Pad) and Excel (Microsoft) software.
Results
Immunoblot analysis of APEX1 and Jagged-1 expression in gastric cancer cells. First, we analyzed the protein expression of APEX1 and Jagged-1 in the gastric cancer cells SNU-1, SNU-5, SNU-16, NCI-N87, KATO III, and AGS. Expression of APEX1 and Jagged-1 was detected by immunoblotting. α-tubulin served as a loading control. The analysis of the western blots revealed that all the gastric cancer cell lines expressed high levels of APEX1, but only two of the gastric cancer cells (NCI-N87 and KATO-III) express both APEX1 and Jagged-1; in particular, only KATO-III cells strongly express both APEX1 and Jagged-1 (Figure 1). Therefore, we selected KATO III and AGS cell lines for further studies. KATO III cells strongly express both APEX1 and Jagged-1, whereas AGS cells strongly express only APEX1.
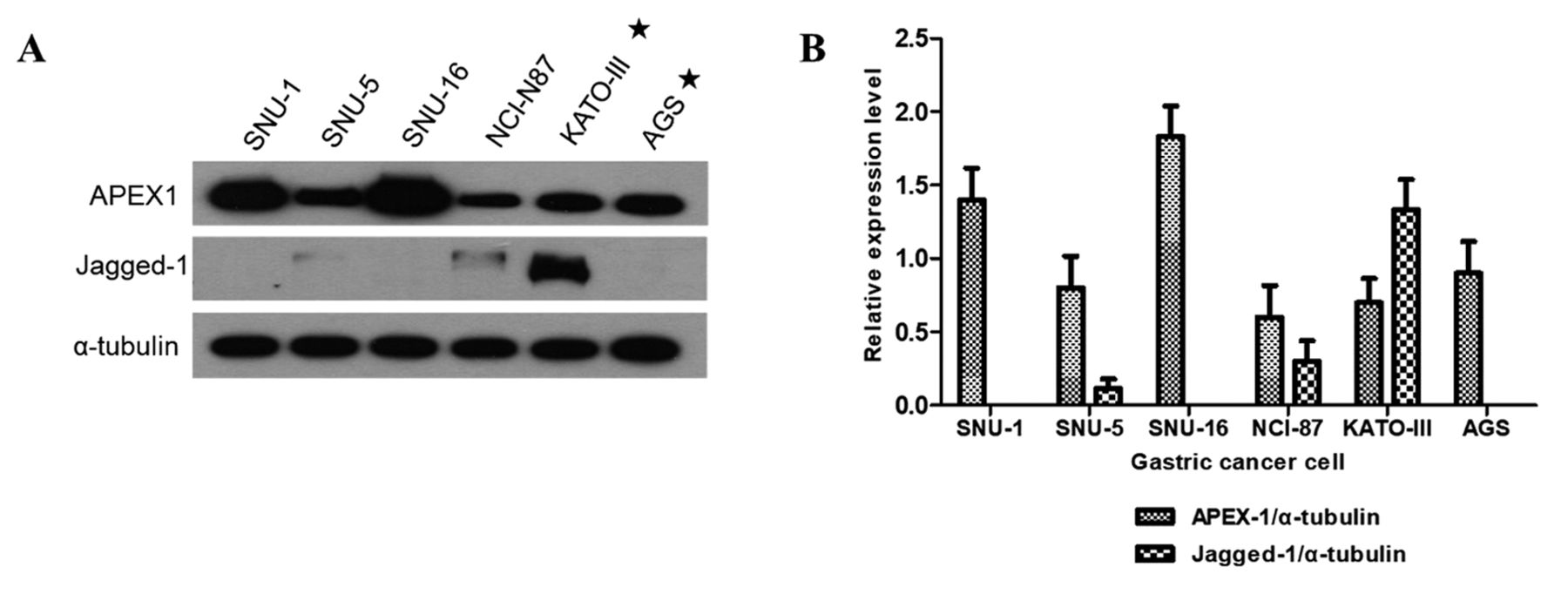
Representative images of western blot analysis for APEX1 and Jagged-1 protein expression in gastric cancer cells. α-tubulin served as a loading control. These results indicated that all gastric cells expressed high levels of APEX1, but only the KATO-III expressed both APEX1 and Jagged-1.
MTT assay following treatment with 5-FU and cisplatin. The MTT assay was used to assess the sensitivity of the four gastric cancer cell lines (SNU-1, SNU-16 and AGS vs. KATO-III) to the well-known and effective drugs: cisplatin and 5-FU (Figure 2). KATO-III cells were more resistant to both the drugs (cisplatin and 5-FU) compared to SNU-1, SNU-16 and AGS cells. The IC50 values in the resistant cell line (KATO-III) were higher than those in the sensitive cell line (SNU-1, SNU-16 and AGS). The IC50 values of 5-FU and cisplatin were 3- and 1.5-fold higher in KATO-III cells, respectively, then those in SNU-1, SNU-16 and AGS cells. These results suggest that co-expression of APEX1 and Jagged-1 may be associated with resistance to chemotherapeutic drugs (5-FU and cisplatin) in atypical glandular cells (SNU-1, SNU-16 and AGC) (Table I).
Efficiency of chemotherapeutic drugs following siRNA APEX1 knockdown. We investigated the drug sensitivity of KATO III and AGS gastric cell lines following APEX1 knockdown. Following APEX1 knockdown, the MTT assay revealed that the IC50 of cisplatin and 5-FU in AGS cells was decreased approximately 7% and 15%, respectively, however, their decrease in chemoresistant KATO-III cells was decreased by approximately 21% and 67% for cisplatin and 5-FU, respectively (Table I). These results suggested that APEX1, by itself, is not significantly associated with chemoresistance in gastric cancer. However, gastric cancer cells co-expressing both APEX1 and Jagged-1 were chemoresistant to cisplatin and 5-FU (Figure 3). These results suggested that Jagged-1 activation by APEX1 provides chemoresistance for cisplatin and especially for 5-FU in gastric cancer.
The results of the MTT assay for the viability of advanced gastric cancer cells following treatment with chemotherapeutic drugs.
APEX1-mediated Jagged-1 inhibition was associated with chemoresistance in a gastric cancer xenograft mouse. Gastric cancer KATO-III/sicontrol (1×106) and KATO-III/siAPEX1 (1×106) cells were subcutaneously injected to the flanks of 6weeks old female nude BALB/c nude mice. The mice were divided into 6 groups; each group contained 5 mice. At 14 days after cell inoculation, we initiated treatment with 5-FU (10 mg/kg, s.c. next to the tumor, once every three days for 3 weeks) and cisplatin (30 mg/kg, s.c. next to the tumor, once every three days for 3 weeks). The control group was intraperitoneally (IP) administered with saline containing 10% dimethyl sulfoxide (DMSO). Tumor volumes were calculated by the following formula tumor volume=[width2 ×length]/2. As shown, KATO-III/siAPEX1 mice treated with 5-FU or cisplatin showed significant reduction in tumor volume and weight compared to KATO-III/sicontrol mice treated with cisplatin or 5-FU (Figure 4A). Specifically, KATO-III/siAPEX1 mice treated with cisplatin or 5-FU in experienced tumor shrinkage as indicated by tumor volume (Figure 4B) and tumor weight (Figure 4C). These results demonstrate that the simultaneous expression of Jagged-1 and APEX1 was associated with chemoresistance in gastric cancer.

MTT assay for viability of advanced gastric cancer cells following treatment with drugs (A: 5-FU, B: Cisplatin). The cells were plated in 96-well plates and treated with 5-FU or cisplatin. The IC50 values for 5-FU and cisplatin in KATO-III cells were 3- and 1.5-fold higher than that in SNU-1, SNU-16, and AGS cells, respectively.

MTT assay for viability of advanced gastric cancer cells following treatment with drugs. Cells were plated in 96-well plates and treated with cisplatin or 5-FU. Following APEX1 knockdown, the chemosensitive cell line (AGS) showed a decrease in IC50 for both the chemotherapeutic drugs by approximately 15% and 7% for 5-FU and cisplatin, respectively (A and B). The chemoresistant cell line (KATO-III) displayed a marked decrease in IC50 for both chemotherapeutic agents by approximately 67% and 21% for 5-FU and cisplatin, respectively (C and D).

Tumor Xenograft Experiments. (A) Photographs of human advanced gastric cancer cells, KATO-III/sicontrol and KATO-III/siAPEX1 (1×106 cells/injection s.c.) were injected into BALB/c nude mice (n=5) for establishing a xenograft model. At 14 days after cell inoculation, mice were separated into six groups: KATO-III/sicontrol (saline containing 10% dimethyl sulfoxide, 5-FU, cisplatin) and KATO-III/siAPEX1 (saline containing 10% dimethyl sulfoxide, 5-FU, cisplatin). Mice were sacrificed 35 days after inoculation. (B) Tumor growth curves. Treatment was initiated on day 14 (arrow: tumor size was ~5 mm). (C) Tumor weight on day 35.

IHC scoring of advanced sections of gastric cancer patient specimens. Correlation between APEX1, Jagged-1 and CD133 expression in gastric cancer patients' tissue. APEX1, Jagged-1 and CD133 proteins in gastric cancer patients' tissue are shown by IHC with anti-APEX1, anti-Jagged-1 and anti-CD133 antibodies. Brown staining reveals positive APEX1, Jagged-1 and CD133 staining (p<0.01, Scale bars, 150 μm) (A). APEX1, Jagged-1 and CD133 expression levels assessed by IHC scoring (B).
IHC scoring of advanced gastric cancer patient tissues. Next, we planned a pilot study to evaluate the clinical significance and expression of APEX1 and Jagged-1 in gastric cancer tissues. Nine patients were categorized into 2 groups based on their response to chemotherapy (chemoresponsive patients vs. chemorefractory patients). We performed IHC staining on paraffin-embedded chemo native cancer tissues from all the patients and the IHC results were scored for the staining intensity. Our results showed that chemorefractory advanced gastric cancer tissues had higher scores (APEX1, 7.5; Jagged-1, 6.6) than the chemoresponsive advanced gastric cancer tissues (APEX1, 3.6; Jagged-1, 2.5) (Figure 5). Therefore, co-expression of APEX1 and Jagged-1 determines the chemotherapeutic response in advanced gastric cancer. Activation of Jagged-1 signaling by APEX1 has been shown to be associated with chemoresistance. Therefore, we investigated the chemoresistance mechanism of Jagged-1 signaling pathway activated by APEX1. We analyzed protein expression of CD133, which is a representative marker for cancer stem cells, in KATO-III gastric cancer cells (strongly expressing both APEX1 and Jagged-1) and AGS (strongly expressing only APEX1) by western blotting. CD133 expression was higher in KATO-III cells compared to AGS cells and it correlated with Jagged-1 expression (Figure 1). Metastatic gastric cancer tissues were also stained for CD133. The resistant group expressed higher levels of CD133 (7.8 times) and stained also for JAGGED-1 compared to the chemo responsive group (3 fold) (Figure 5). Although this was a pilot study with only 9 cases, the results suggest that the mechanism of resistance of the Jagged-1 signaling pathway activated by APEX1 may be related with proliferation of cancer stem cells. Further research on jagged-1, notch signaling and cancer stem cells may be needed.
Discussion
The treatment outcomes of advanced gastric cancer are determined by the stage at the time of presentation. Complete resection is the only potentially curative intervention. Patients with unresectable advanced gastric cancer have extremely poor survival rates of less than 12 months after diagnosis (1, 2). Cytotoxic chemotherapy is the only palliative treatment and chemoresistance remains a major problem. The mechanisms of chemoresistance are diverse and have not been fully understood yet. Several theories have been proposed, such as membrane transporters, drug inactivation, DNA repair of chemotherapy induced DNA damage, dysregulation of cell survival, and a combination of different mechanisms. However, very little information is available regarding the molecular chemoresistance factors in gastric cancer (3, 11-13). Currently, combination chemotherapy with 5-fluorouracil and platinum is the most effective chemotherapy regimen for gastric cancers (2, 14, 15). Cisplatin is a highly potent DNA-damaging chemotherapeutic platinum agent, which interferes with DNA replication and kills the fastest proliferating cells (16). The mechanism of 5-FU cytotoxicity has been recognized to be the misincorporation of fluoronucleotides into RNA and DNA through inhibition of the nucleotide synthetic enzymes such as thymidylate synthase (17). Some studies have shown that APEX1 expression levels were correlated with chemoresistance in various cancer cells (4, 5). However, no study has been conducted to evaluate the role of APEX1 in advanced gastric cancer cells. Our research suggests that APEX1 may be associated with chemoresistance in advanced gastric cancer.
Notch signaling has emerged as a focus of intense research in various solid tumors and the Notch signaling pathway has been determined to be the driver of gastric epithelial cell proliferation. Jagged-1-induced Notch activation is involved in various aspects of tumor growth through the maintenance of cancer stem cells, promotion of cancer cell survival, inhibition of tumor apoptosis, and driving malignancy proliferation and metastasis. Furthermore, it affects not only the cancer cells, but also components of the tumor micro environmental. High expression of Jagged-1 has been reported to be a poor prognostic factor in breast cancer, bladder cancer, leukemia, prostate cancer, biliary cancer, and gastric cancer (6, 8). In gastric cancer, the activation of Jagged-1 has been associated with poor prognosis and Jagged-1 has also been suggested as a prognostic biomarker for overall survival and postoperative clinical outcome. Moreover, the involvement of Jagged-1-induced Notch pathway activity in chemoresistance has been reported (18-21). However, the role of Notch pathway involving Jagged-1 activation by APEX1 in chemoresistance has not been investigated in gastric cancer.
Previous studies have shown that APEX1 mediates Jagged-1 activation through two mechanisms. First, APEX1 was shown to act as a positive regulator of Jagged-1/Notch activity in colon cancer; wherein, colon cancer progression was driven by APEX1-mediated upregulation of Jagged-1 (9). Second, Jagged-1 activation by APEX1 was shown to induce chemoresistance in advanced biliary cancer and the co-expression of APEX1 and Jagged-1 was associated with chemoresistance in biliary cancer (10). Additionally, APEX1-induced expression of Jagged-1 appears to be important in the chemoresistance to 5-FU, oxaliplatin, and irinotecan in colorectal cancer. The co-expression of APEX1 and Jagged-1 might be used as a potential biomarker to predict poor response to chemotherapy in colorectal cancer (22).
Our results showed that in the absence of Jagged-1, APEX1 expression is related to a certain extent to chemoresistance in gastric cancer. However, co-expression of both APEX1 and Jagged-1 resulted in chemoresistance of gastric cancer cells to cisplatin or 5-FU. Also, the tumor volume of KATO-III/sicontrol mice was less affected by the treatment with cisplatin or 5-FU compared with KATO-III/siAPEX1 mice treated with cisplatin or 5-FU. Our results demonstrate that the simultaneous expression of Jagged-1 and APEX1 was associated with chemoresistance in gastric cancer.
The exact chemoresistance mechanism of APEX1-driven Jagged-1 activation has not been identified yet. However, we have observed that Notch signaling plays an important role in the survival and persistence of cancer stem cells. Recent studies have suggested that cancer stem cells promote resistance of chemotherapy. One of the best markers of cancer stem cells is CD133 (23-27).
Finally, our present study revealed that the co-expression of Jagged-1 and APEX1 was associated with chemoresistance in advanced gastric cancer. Therefore, activation of Jagged-1 by APEX1 predicts the chemo-response and may provide an important tool for the selection of the appropriate therapeutic strategy for neoadjuvant chemotherapy for patients with borderline resectable gastric cancer.
Acknowledgements
This work was supported by a grant from the Clinical Medicine Research Institute of the Chosun University Hospital (2016).
Footnotes
↵* These Authors contributed equally to this study as co-first authors.
Authors' Contributions
Kim HB and Lee HJ performed the experiments and analyzed the data; Kim GB and Lim HJ drafted and contributed to writing the manuscript; Park SG drafted, wrote, edited, and reviewed the manuscript. All Authors read and approved the final manuscript.
Conflicts of Interest
The Authors declare no conflicts of interest regarding this study.
- Received February 20, 2020.
- Revision received March 2, 2020.
- Accepted March 4, 2020.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}