


Abstract
Background/Aim: Resistance to chemotherapeutic agents is the main cause of reduced survival in non-small cell lung cancer (NSCLC) patients. The Hedgehog (HH) pathway has been shown to be crucial in cell development and survival. Activated in several types of cancer it might be a potent bypass mechanism mediating chemotherapy resistance. Materials and Methods: HCC827 NSCLC cells were treated with sub-lethal doses of pemetrexed to produce pemetrexed resistance. RT-qPCR was performed to measure gene expression of HH pathway proteins. A cell growth assay was used to measure the impact of the HH-inhibitor Gant61 in naïve and chemoresistant cell lines. Results: Pemetrexed resistant cells showed significantly increased expression of HH signaling genes (GLI1, GLI2, GLI3, PTCH1, SHH). Supporting these results, pemetrexed resistant cells treated with the HH inhibitor Gant61 showed reduced proliferation compared to naïve cells. Conclusion: HH pathway may play an important role in mediating pemetrexed resistance in NSCLC cells. Blocking the HH pathway may be a potential option to overcome this resistance.
Worldwide, lung cancer is one of the most common malignant tumors and non-small cell lung cancer (NSCLC) accounts for about 85% of all lung cancer cases (1). The combination of pemetrexed and cisplatin (plus immunotherapy) is one of the current first-line standard treatments for NSCLC adenocarcinoma without an oncogenic driver mutation. Furthermore, pemetrexed is widely used as maintenance treatment (2-5). As a multi-target folate antagonist pemetrexed inhibits tumor cell growth by inhibiting the normal acid metabolism of folate in the cells and thus inhibits DNA synthesis in tumor cells (6, 7). However, as with other chemotherapeutic drugs, primary or secondary resistance to pemetrexed treatment limits its clinical application.
The Hedgehog (HH) pathway is involved in the embryonic phase, cell differentiation, organ formation and development (8-10). It is activated in many cancer entities, such as lung cancer, pancreatic cancer, myeloid leukemia, ovarian cancer and glioblastoma and is thought to be an important oncogenic driver (11-15).
However, HH signaling not only participates in the formation and development of tumors, but has also been shown to promote therapy resistance (16-19). Thus, we sought to examine whether HH signaling is involved in pemetrexed resistance in NSCLC. Accordingly, this study aimed at investigating the role of the HH pathway in pemetrexed resistant cells and the ability to use this pathway as a potential therapeutic target.
Materials and Methods
Cell culture. HCC827 NSCLC cells (ATCC, Manassas, VA, USA) were cultured in RPMI1640 medium (Sigma–Aldrich, Deisenhofen, Germany) containing 10% fetal bovine serum (Invitrogen, Carlsbad, CA, USA), 100 U/ml penicillin (Biochrom GmbH, Berlin, Germany) and 100 μg/ml streptomycin (Biochrom GmbH, Berlin, Germany) at 37°C in an incubator containing 5% CO2 and saturated humidity.
Obtaining resistance of cells to pemetrexed. To obtain resistance to pemetrexed, HCC827 cells in the logarithmic phase of growth were treated with 130 nM of pemetrexed for 24 h every week. After 4 weeks the cells were able to grow normally in a culture medium containing 130 nM pemetrexed.
Quantitative real-time reverse transcription polymerase chain reaction (qPCR). Total RNA was extracted from cell lines following the Trizol method (Sigma–Aldrich, Deisenhofen, Germany). Total RNA was depleted from DNA and subsequently purified according to the manufacturer's manual. Reverse transcription of total RNA was performed using random hexamers (Roche Diagnostics, Penzberg, Germany) and SuperScriptII reverse transcriptase (Invitrogen). qPCR amplifications of the respective genes were carried out using 40 ng complementary DNA, 500 nM forward and reverse primer, and iTaq SYBR Green Supermix (Bio-Rad Lab-oratories, Hercules, CA, USA) in a final volume of 20 μl. PCR reactions were run for 40 cycles that consisted of 15 sec' denaturation at 95°C, primer annealing for 15 sec at 55°C, and extension for 30 sec at 72°C. We used the following primer pairs (5’->3’ orientation): GLI1, AGCTACATCAACTCCGGCCA, GCTGCGGCGTTCAAGAGA; GLI2, TTCTCCAACGCCTCGGAC, GCCTGGGATCTTGCAGATGT; GLI3, TCAAAGCGGGAAGAATGCC, CTGACCACCAGGGCTTGG; PTCH1, TTGATTGTGGGTGGCACAGT, GCTTGGGAGTCATTAACTGGAAC; SMO, GGAGAGGAGCCATACTGCCC, TCAACCAGCCACAGGTTGG; SHH, CTGGGTGTACTACGAGTCCAAGG, CAGCCTCCCGATTTGGC; HHIP, TGTACATCATTCTTGGTGATGGG, AGCCGTAGCACTGAGCCTGT; TBP, GCCCGAAACGCCGAATAT, CCGTGGTTCGTGGCTCTCT. qPCR amplifications were performed on Life Touch Cycler (Bioer, Hangzhou, China). All experiments were performed in doublets. Amplification of the housekeeping gene TATA-Box-binding-Protein (TBP) was performed to standardize the amount of sample RNA. Relative quantification of gene expression was performed using the ΔΔ-ct method as described earlier (20).
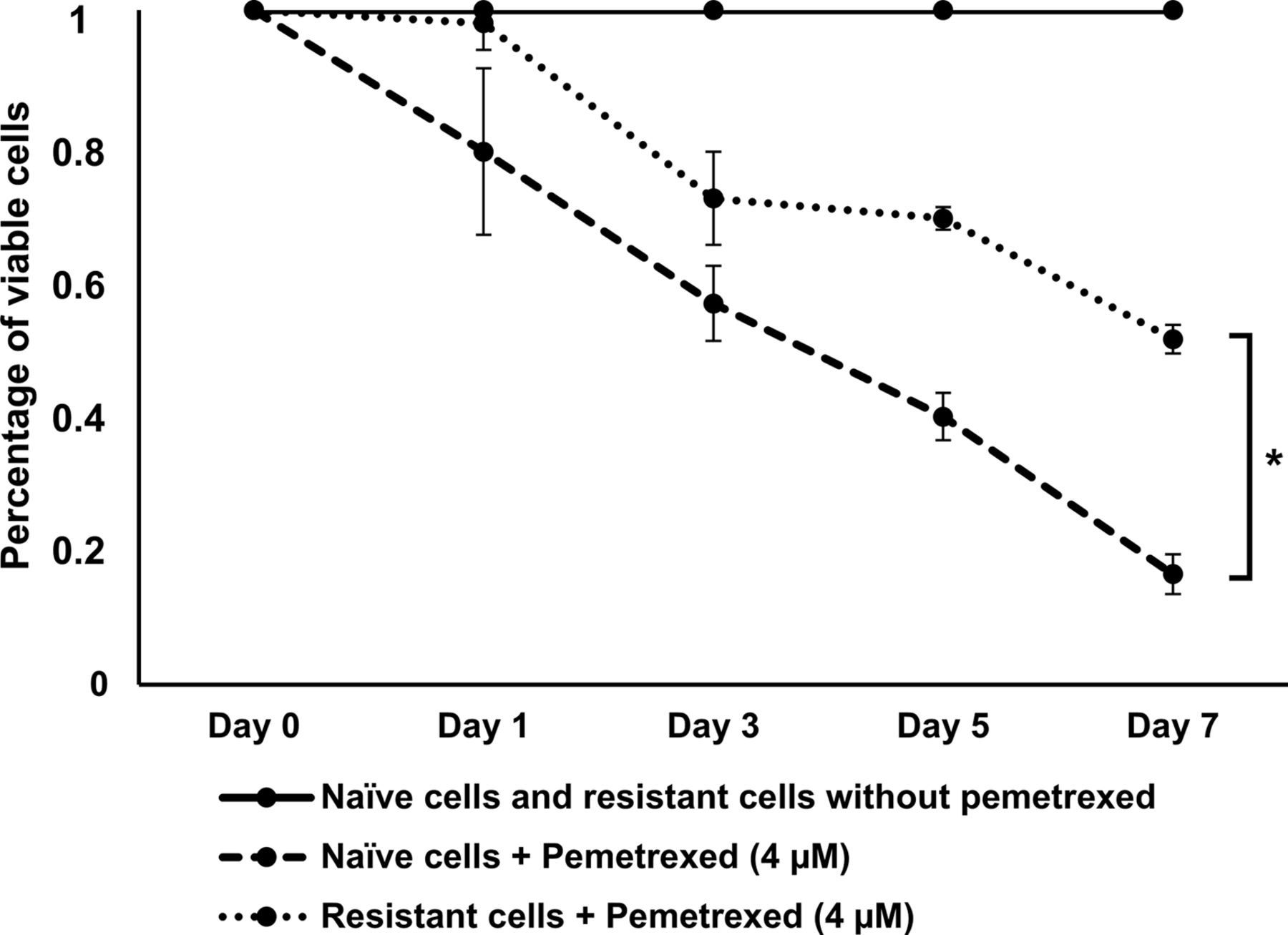
Cell viability of naïve and pemetrexed resistant cells treated with pemetrexed. Pemetrexed [4 μM (IC50)] had a stronger effect on naïve cells compared to pemetrexed resistant cells.
Measurement of cell viability. HCC827 naïve cells and HCC827 pemetrexed-resistant cells were seeded in 96-well plates at 2×103/well and cultured overnight. Cells were treated with pemetrexed (4 μM) and/or Gant61 (27.7 μM). The DMSO-treated control group and the non-treated blank group were set up in the same way. Cell viability was tested on day 1, day 3, day 5 and day 7 respectively by adding 100 μl Celltiter-Glo reagent (Promega Corporation, Madison, WI, USA) according to the manufacturer's manual. The luminescence values for each well were measured by a Victor plate reader (Berthold Technologies, Bad Wildbad, Germany).
Statistical analyses. All data were expressed as means±standard deviation and analysed with the Student unpaired t-test. A level of p<0.05 was considered statistically significant. All analyses were performed using SPSS 24 statistical software (IBM Corp., Armonk, NY, USA).
Results
Pemetrexed resistant NSCLC cells show increased expression of genes involved in the hedgehog pathway. HCC827 cells were treated with sub-lethal doses of pemetrexed to induce pemetrexed resistance. Figure 1 shows that the viability of naïve HCC827 cells was significantly lower following treatment with pemetrexed at the IC50 concentration than pemetrexed resistant cells.

mRNA expression levels were quantified by qPCR. SHH (5.9-fold), GLI2 (4.3-fold), GLI1 (3.7-fold), GLI3 (3.5-fold) were significantly upregulated in pemetrexed resistant HC827 cells compared to naïve cells.
qPCR was used to quantify mRNA expression levels of relevant hedgehog pathway proteins. The ligand of hedgehog signalling SHH revealed to be expressed 5.9-fold compared to naïve cells. Besides, expression of the transcription factors GLI1, GLI2 and GLI3 was significantly increased 3.7-, 4.3- and 3.5-fold, respectively, compared to non-resistant cells. The levels of the trans-membrane receptor PTCH1, the inhibitor of the hedgehog pathway HHIP and the receptor associated protein SMO were not significantly elevated in pemetrexed resistant cells (Figure 2).
HH inhibitor Gant61 inhibits cell growth in pemetrexed resistant cells more effectively and might restore pemetrexed sensitivity. As the expression of the transcription factors involved in the hedgehog pathway GLI1, Gli2 and GLI3 was significantly elevated in pemetrexed resistant cells we sought to examine whether the GLI-inhibitor Gant61 could inhibit cell growth in pemetrexed resistant HCC827 cells and whether treatment with Gant61 might restore pemetrexed sensitivity.
Figure 3A shows that treatment with Gant61 significantly reduced cell viability in pemetrexed resistant cells. Furthermore, the addition of pemetrexed improved the effectiveness of the treatment, indicating a possible restoration of pemetrexed sensitivity in pemetrexed resistant cells by concomitant blockage of the hedgehog pathway. Besides, treatment of naïve HCC827 with Gant61 plus pemetrexed or Gant61 alone did not result in reduced cell viability compared to treatment with pemetrexed alone (Figure 3B).
Interestingly, both naïve and resistant cells showed good response to Gant61 alone. Nevertheless, the inhibition seems to be stronger in the resistant cells. This might be due to the higher activity of the hedgehog signalling in these cells (Figure 4).
Discussion
Chemotherapy resistance not only affects the efficacy of chemotherapeutic drugs, but also promotes tumor recurrence and metastasis and accordingly reduces patients' survival. Thus, understanding the underlying mechanisms of chemotherapy resistance would help to pilot treatment and select patients who will benefit from a distinct therapy. Pemetrexed is a multi-target anti-folate drug that inhibits cell growth by interfering with the metabolism of folate in tumor cells during replication, and has been shown to be very effective in first line and maintenance treatment of lung adenocarcinoma (6, 7). However, many patients show primary or acquired resistance to pemetrexed (21). Several mechanisms have been proposed regarding the emergence and maintenance of pemetrexed resistance in NSCLC. For example, the overexpression of thymidylate synthase (TYMS) (22) and uracil–DNA glycosylase (UNG) (23), as well as the low expression of solute carrier family 19 member1 (SLC19A1) (24). In this context, our study aimed at investigating the influence of HH signaling on the emergence and maintenance of pemetrexed resistance in NSCLC cells.

Impact of Gant61 treatment and its combination with pemetrexed on cell viability in resistant and naive cells. (A) Gant61 significantly reduced cell viability in pemetrexed resistant cells. Addition of pemetrexed further improved the effectiveness of Gant61. (B) Gant61 plus pemetrexed or Gant61 alone did not result in reduced cell viability compared to pemetrexed treatment.

Treatment of naïve and resistant cells with Gant61. Both naïve and resistant cells showed good response to Gant61 treatment. Nevertheless, inhibition of cell viability was stronger in resistant cells and was associated with higher activity of hedgehog signalling in these cells.
Compared with non-resistant NSCLC cells, genes involved in the HH pathway were overexpressed in pemetrexed-resistant cells. In line with these results, some previous studies have also shown that SHH is overexpressed in non-small cell lung cancer and may be involved in chemotherapy resistance (25-27). Furthermore, our experiments revealed that pemetrexed-resistant cells were more sensitive to pemetrexed when combined with the HH inhibitor Gant61, which possibly indicates that pemetrexed-resistance could be reversed in these cells by inhibiting HH signaling. Thus, the HH pathway may be a possible new target to reverse pemetrexed-resistance in NSCLC cells. Another inhibitor of the HH pathway, vismodegib, has been reported to have anti-tumor activity by binding to SMO and inhibiting SMO signal transduction (28). However, it has been shown that vismodegib is less toxic to tumor cells with diffusely expressed SMO, and is less effective in inhibiting tumor cell growth and promoting tumor cell necrosis compared to Gant61 in NSCLC (29, 30). In this study, blocking the HH pathway with vismodegib did not result in a significant inhibition of tumor growth in resistant or in naïve cells (data not shown). This might be due to SMO-independent GLI signal transduction in NSCLC (31). In our study, we found that Gant61 itself has a strong inhibitory effect on the growth of pemetrexed-resistant and naïve NSCLC cells.
HH signaling pathway has been shown to be involved in chemo resistance in several tumors such as lung cancer (32), breast cancer (33), pancreatic cancer (34) or gastric cancer (35). The results of this study are in line with these studies indicating that HH signaling might be involved in pemetrexed resistance in NSCLC cells. However, the mechanism by which HH signaling mediates chemotherapy resistance is not very clear. HH signaling alters the regulation of epithelial mesenchymal transition (EMT) in tumor cells (36-38), which might lead to decreased sensitivity to chemotherapy. Furthermore, the HH pathway regulates E-cadherin expression through its downstream target proteins such as Snail, Slug, ZEBI1 and TWIST (38-40). By doing this, the HH pathway indirectly or directly regulates the EMT process. ABC transporters, that carry intracellular and exogenous compounds out of the cells are overexpressed in many tumor cells leading to chemo resistance. Recent studies have found that EMT-associated transcription factors can activate the promoter of the ABC transporter (41, 42). The overexpression of transcription factors such as Twist, Snail and FOXC2 that are regulated by the HH pathway also increases the activity of the ABC transporter promoter, which enhances the resistance of tumor cells to chemotherapeutic drugs (41, 42). Furthermore, HH signaling induces Bcl-2 expression and thus inhibits tumor cell apoptosis (43). In conclusion, our results are the first to indicate an impact of the HH signaling in pemetrexed resistance of NSCLC cells. However, the results need to be confirmed in further experimental and clinical studies.
Acknowledgements
The financial support from China Scholarship Council (CSC) is acknowledged. Yichao Liu (Award No. 201708080182) received a three-year scholarship of the China Scholarship Council (CSC).
Footnotes
Authors' Contributions
YL, RH, AT and DK designed the study, performed the literature search, and wrote and revised the manuscript. YL and RK performed experimental procedures, and YL, RH, AR and DK performed the statistical analysis.
Conflicts of Interest
The Authors declare no conflicts of interests. The abstract of this paper was presented at the Conference ELCC 2019 as a poster presentation with interim findings. The poster's abstract was published in Annals of Oncology 30(Supplement_2), DOI: 10.1093/annonc/mdz073.017
- Received January 22, 2020.
- Revision received February 1, 2020.
- Accepted February 11, 2020.
- Copyright© 2020, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.